
SIMULATED CHANGES IN PLANT DISTRIBUTIONS
The geographic ranges of plant species are controlled by climate change--in this section, we explore potential changes in the distributions of selected western plant species due to future climate change. Following the methods described in Lipsitz (1988) and Bartlein and others (1994), we interpolated present-day climatic data onto a 15-km equal-area grid of the Western United States to provide a fine-scale basis for the exploration of potential changes in plant distributions (fig. 9). The differences between the RegCM-simulated present-day climate and the RegCM-simulated 2 X CO2 climate were applied to this target grid to provide inputs into vegetation models simulating potential changes in plant distributions that would occur when comparing present-day and potential future climates.
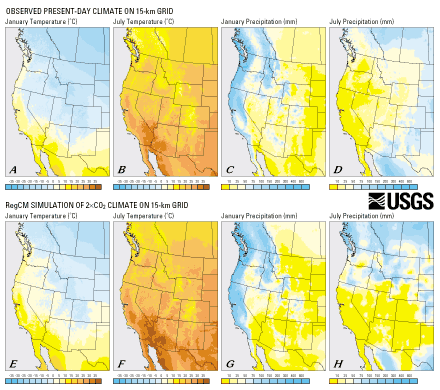
| Figure 9. Observed present-day (A-D) and simulated 2 X CO2 (E-H) climates illustrated on 15-km grid. These latter values were formed by interpolating the RegCM climate anomalies (see bottom row of fig. 3) onto the 15-km grid and adding interpolated values to present-day values. (Click here for scalable 788K PDF version) |
PRESENT-DAY CLIMATE AND PLANT DISTRIBUTIONS
Bartlein and others (1994) prepared an equal-area 25-km grid of 30-year climate "normals" for North America. January and July temperature and precipitation data from this grid were used in the analyses presented below. GSMAP software (Selner and Taylor, 1992) was used to digitize maps of tree distributions (Little, 1971, 1976) and to determine which of the 32,211 points in the North American climate grid were within the geographic range of the each taxon and which were outside of this range. The presence/absence data were matched with temperature and precipitation data to determine the present-day relationships between plant distributions and climate (Thompson and others, in press). The distributions of 16 common western trees and shrubs (table 2) are analyzed for the present study.
Table 2. Common and scientific names of plant species with simulated present-day ranges and potential ranges under a simulated 2 X CO2 future climate. [The 15-km grid used in this study has 23,961 cells in western North America. "Present number" and "2 X CO2 number" are the number of those cells occupied by the species at the 0.40 level of probability for the present-day and potential future climate simulations, respectively. "Percent of present coverage" represents the percentage increase or decrease of the taxon under the future climate scenario compared to the present-day simulation (present-day range = 100 percent)] | ||||
| Common name | Scientific name | Present number | 2 X CO2 number | Percent of present coverage |
| Sitka spruce | Picea sitchensis | 493 | 1,033 | 210 |
| Western red cedar | Thuja plicata | 1,010 | 1,143 | 113 |
| Western hemlock | Tsuga heterophylla | 945 | 1,185 | 125 |
| Incense cedar | Libocedrus decurrens | 517 | 1,103 | 213 |
| Engelmann spruce | Picea engelmannii | 3,387 | 566 | 17 |
| Douglas fir | Pseudotsuga menziesii | 4,898 | 2,474 | 51 |
| Lodgepole pine | Pinus contorta | 4,350 | 1,508 | 35 |
| Ponderosa pine | Pinus ponderosa | 2,319 | 3,147 | 136 |
| Oregon white oak | Quercus garryana | 626 | 1,330 | 212 |
| California white oak | Quercus lobata | 447 | 3,322 | 743 |
| Gambel oak | Quercus gambelli | 753 | 959 | 127 |
| Pinyon pine | Pinus edulis | 974 | 935 | 96 |
| Big sagebrush | Artemisia tridentata | 5,733 | 2,356 | 41 |
| Joshua tree | Yucca brevifolia | 187 | 1,482 | 793 |
| Cresote bush | Larrea divaricata | 3,891 | 6,235 | 160 |
| Saguaro | Cereus giganteus | 626 | 806 | 129 |
Scatter diagrams (fig. 10) illustrate the climatic tolerances of four of these plants in terms of seasonal temperature and precipitation. For example, Sitka spruce lives where January mean temperatures are above -10oC and mean July temperatures are below 20oC. In contrast, Douglas fir lives under a wide variety of temperature regimes, and the frost-sensitive species, California white oak and Joshua tree, live where mean January temperatures are above freezing. Sitka spruce is largely limited to sites with very high levels of January precipitation and relatively low levels of July precipitation. Douglas fir lives in a range of seasonal precipitation regimes, whereas California white oak lives in areas with high winter precipitation and pronounced summer drought. Joshua tree lives in very dry climates where winter precipitation tends to exceed that of summer.
| Figure 10. Bivariate scatter plots of January vs. July temperature (left column) and January vs. July log precipitation (right column) showing presence/absence data (green and gray, respectively) for four plant species in western North America. These illustrations depict the unique climatic adaptations of various tree species in western North America. Sitka spruce (A, B), a tree of the moist Pacific Northwest coast, lives in some of the wettest winter climates on the continent under January temperatures between +10oC and +10oC and July temperatures between 5oC and 20oC. Douglas fir (C, D) lives under a broad band of seasonal temperatures and precipitation conditions. California white oak (E, F) is restricted to environments west of the Sierra Nevada in California and lives under a fairly wide range of July temperatures but a very narrow band of January temperatures. This tree receives essentially all of its moisture in the winter and lives under pronounced summer drought. Joshua tree (G, H), an arborescent yucca of the Mojave Desert, lives in places where summers are extremely hot and winters are rarely below freezing. It survives in very arid environments. (Click here for scalable 11M PDF version) |  |
COMPARISON OF PRESENT-DAY AND POTENTIAL FUTURE RANGES
The relationships illustrated in figure 10 can be analyzed statistically to produce derived presence/absence response surfaces (e.g., Bartlein and others, 1986) that determine the probability of occurrence for each plant for a given combination of seasonal temperature and precipitation characteristics. These statistical relationships can then be used with observed climate data to estimate the present-day range of the species. The relationships can also be used with climate-model output to simulate the potential future range of the plant. Figure 11 shows the simulated present-day and potential future ranges of the four plants discussed above. In the left-hand illustration of each row (figs. 11A, 11E, 11I, 11M) is shown the observed present-day range of the plant on the 15-km target grid. For comparison, the next illustration to the right (figs. 11B, 11F, 11J, 11N) shows the probability of occurrence of the species produced by response-surface analysis of the observed present-day climatic and plant-distribution data. In most cases, the present-day range is aligned with approximately the 0.4 probability of occurrence. The third illustration from the left (figs. 11C, 11G, 11K, 11O) illustrates the simulated probability of occurrence for the same taxa under the 2 X CO2 climate simulation interpolated onto the 15-km target grid. The right-hand illustrations (figs. 11D, 11H, 11L, 11P) illustrate changes in the distribution of each species at the 0.4 probability level between the present-day simulated distribution and the distribution calculated for the 2 X CO2 climate. In this case (i.e., the right-hand illustrations of the figure), green represents areas occupied by the species under both the present-day and 2 X CO2 climate simulations, red represents range lost by the species between the present-day and 2 X CO2 simulations, and blue represents new areas that could potentially be colonized by the species under the 2 X CO2 climate simulation. In the examples presented here, all of the four species could potentially increase their total coverage, although not necessarily within or adjacent to their current range.

Figure 11 (above and below). Maps of plant-species distributions showing, by column from left to right, observed present-day distributions (A, E, I, M) (Little, 1971, 1976), simulated present-day distributions at 0.4 probability of occurrence (B, F, J, N), simulated distributions for 2 X CO2 climate at 0.4 probability of occurrence (C, G, K, O), and changes in species distribution between simulated present-day distribution and simulated distribution under 2 X CO2 climate (D, H, L, P). Species shown are Sitka spruce (A-D), Douglas fir (E-H), California white oak (I-L), and Joshua tree (M-P). Comparison of the simulated present-day distributions with observed present-day distributions demonstrates that the presence/absence response surface methodology captures the overall distribution pattern of all of the species. Comparison of the simulated present-day distributions and the simulated 2 X CO2 distributions illustrates the potential range shifts that could occur if species were able to migrate rapidly enough to reach their new potential habitats during the transition from the present-day climate to a 2 X CO2 climate. The right-hand columns of maps (D, H, L, P) show these shifts more directly-green represents sites where species live today and where they could continue to live under the simulated 2 X CO2 climate; red indicates sites where species live today but could not survive under the simulated 2 X CO2 climate; blue represents sites where species cannot live today but could potentially live under the simulated 2 X CO2 future climate. (Click on A for scalable 484K PDF version or B for scalable 442K PDF version) |

Figure 12 illustrates potential range changes that could occur under the 2 X CO2 climate simulation. Here the habitats of plants along the Pacific Coast from northern California to British Columbia (Sitka spruce, western red cedar, western hemlock) are reduced slightly, but new habitat develops in the northern interior (figs. 12A, 12B, 12C). Incense cedar nearly dies out in its current range in California and Oregon, but potential habitat opens up in the present-day steppe environment of the Four Corners region (fig. 12D). Under this scenario, major forest trees and range shrubs (Engelmann spruce, Douglas fir, lodgepole pine, and big sagebrush) die off across much of their present-day range without much replacement habitat becoming available (figs. 12E, 12G, 12M). Ponderosa pine would lose much of its western range and find new potential habitat east of its present-day limits (fig. 12H). California white oak and Oregon white oak could potentially grow under a 2 X CO2 winter-wet summer-dry climate in the Southwest (figs. 12I, 12J). Gambell oak and pinyon pine would die off in the Southwest, but could potentially find new range in the northern interior (figs. 12K, 12L). Joshua tree and creosote bush, two shrubs adapted to arid conditions, would considerably expand their spatial coverage relative to today under the 2 X CO2 simulation (figs. 12N, 12O). Saguaro, a frost-limited, dry-adapted plant of the Sonoran Desert, would largely die off in its present-day range, but could potentially find new range farther east and at higher elevations (fig. 12P).
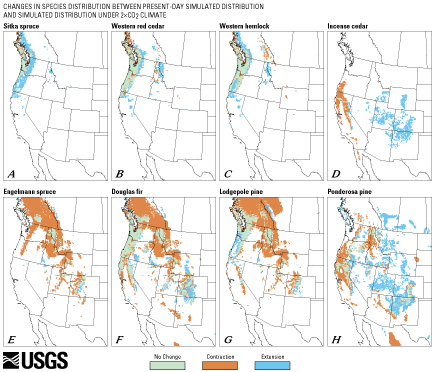
Figure 12 (above and below). Maps showing potential changes in distributions between simulated present-day distributions and simulated distributions under 2 X CO2 climate that could occur for 16 tree species in western North America. Green represents sites where a species lives today and where it could continue to live under the simulated 2 X CO2 climate. Red indicates sites where a species lives today but could not survive under the simulated 2 X CO2 climate. Blue represents sites where a species cannot live today but potentially could live under the simulated 2 X CO2 future climate. All species show some change in potential distribution, although, in some cases, these changes are relatively small (such as those for Sitka spruce (A), western red cedar (B), and western hemlock (C), each of which has a general expansion of their ranges in the Pacific Northwest and into the Northern Rocky Mountains). In other cases, there are large changes in the potential distributions of some taxa (i.e., Gambell oak (K), incense cedar (D), and pinyon pine (L)). Some taxa with rather restricted distributions at present become widespread (i.e., Joshua tree (N)), whereas others with widespread present-day distributions become restricted under a 2 X CO2 climate scenario (i.e., lodgepole pine (G) and Engelmann spruce (E)). The changes illustrated by these maps are specific to the particular set of climate simulations used here but are probably representative of the kinds of general changes that may occur. (Click on A for scalable 164K PDF version or B for scalable 170K PDF version) |

If dispersal rates and (or) human intervention allowed these plants to reach their new potential habitats, how large would the changes be in the coverage of each species? Joshua tree and California white oak have potential future ranges under the 2 X CO2 simulation that would cover more than seven times their current total area (table 2). Incense cedar, Oregon white oak, and Sitka spruce would expand to more than twice their current areal extents; and creosote bush, ponderosa pine, and saguaro would have somewhat greater-than-present-day coverage, although generally not within their current range limits. Western hemlock and western red cedar could occupy modest amounts of new range near their present-day limits, whereas Gambell oak and pinyon pine would have slightly larger and slightly smaller coverages, respectively--but these species would have to disperse hundreds of kilometers northward to reach their new habitats. Douglas fir, big sagebrush, and lodgepole pine would have half or less of their present-day extents, whereas Engelmann spruce would have less than one-fifth of its present-day areal coverage. Could some of the forest trees have potential habitat north of the 15-km target grid under the 2 X CO2 climate simulation? This is probably not the case--if potential habitat existed north of the target grid, it would be logical to assume that the species would be surviving in cool habitats in higher elevations on the northern edge of the grid. Inspection of the output (fig. 12) indicates that this is not the case. The surviving trees are instead located in the lower elevations in this region.
DISCUSSION OF CHANGES IN PLANT DISTRIBUTIONS
The range changes simulated to occur between today and the 2 X CO2 climate would dramatically affect ecosystems across the Western United States. Similarly large shifts in plant distributions have been simulated for a potential 2 X CO2 climate in Europe (Dahl, 1990; Huntley and others, 1995; Sykes and others, 1996). The plant species modeled here exhibit individualistic responses to climatic change, as has been demonstrated in the response of western plants to warming following the end of the last Ice Age (Thompson, 1988; Betancourt and others, 1990; Thompson and others, 1993). In the latter case, warming was relatively gradual and plants had thousands of years to disperse from their old ranges to new ones. In contrast, the anticipated onset of "greenhouse" warming may occur in only a few decades, and it is unknown whether or not displaced plants and animals could migrate to new habitats this rapidly. Many widespread taxa would lose much of their current range under the 2 X CO2 climate, whereas other presently uncommon taxa could greatly expand their ranges (table 2). Similar post-Ice-Age patterns have been documented in packrat midden plant assemblages, illustrating the transition from Pleistocene to Holocene vegetation in the American Southwest (Thompson, 1988). For example, pinyon pine (Pinus edulis) was relatively rare during the last Ice Age, whereas papershell pinyon pine (Pinus remota) was common (Van Devender, 1990), a situation that reversed during the transition from Pleistocene to Holocene conditions.
The economic impacts of projected changes in vegetation for the 2 X CO2 climate could be substantial. Many forest trees, such as Engelmann spruce and lodgepole pine, could be driven to the edge of extinction. The habitats of National Parks, Bureau of Land Management (BLM) lands, and Native American lands would be strongly affected by these and other associated changes (e.g., Bartlein and others, 1997). Land managers could be faced with deciding if they should be protecting the land or the ecosystems presently occupying the lands. The hierarchy of climate and process models used here provides detail and geographic scope unavailable from the current generation of AGCM's and employs embedded physics and process models that enhance the regional depiction of climate change. Analyses such as this provide a means for land-resource managers to identify and assess elements of ecosystems that are sensitive to climatic variability and climate change.
| Contents | Top | Previous Section | Next Section |
| AccessibilityFOIAPrivacyPolicies and Notices | |
| |
|