
Prepared in cooperation with the U.S. Air Force, Arnold Air Force Base and the University of the South
Tree-Regeneration and Mortality Patterns
Hydrologic Change in Sinking Pond, 1854-2002
Ecological Effects of Hydrologic Change in Sinking Pond
Conclusion: Management and Scientific Implications of Hydrologic and Ecolog...
Appendix A. Flora of Sinking Pond.
Appendix B. Evaluating Effects of Local Human Activity on the Hydrology of ...
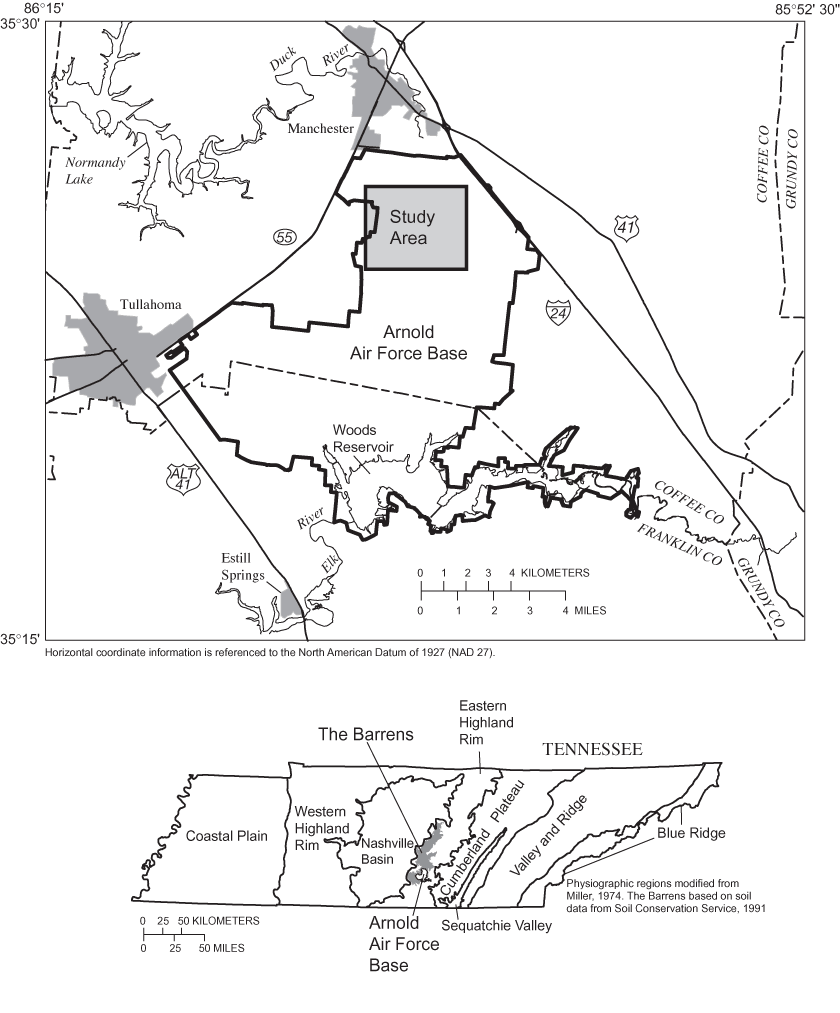
Figure 1. Major physiographic regions of Tennessee and locations of The Bar...
Figure 2. Location of Sinking Pond, adjacent drainage basins, monitored wel...
Figure 3. Water-surface elevations in Sinking Pond and adjacent well 355 fo...
Figure 4. Normal (30-year mean, computed decennially) annual precipitation ...
Figure 5. Photographs of areas (A) in the interior and (B) along the margin...
Figure 6. Aggregate mortality of adult overcup oaks by ponding depth and si...
Figure 7. Distribution of overcup oak by ponding-depth class and life-histo...
Figure 8. Size-class distribution of adult overcup oak in a 2.3-hectare are...
Figure 9. Distribution of sweetgum by ponding-depth class and life-history ...
Figure 10. Size-class distribution of adult sweetgum in a 2.3-hectare area ...
Figure 11. Distribution of willow oak by ponding-depth class and life-histo...
Figure 12. Size-class distribution of adult willow oak in a 2.3-hectare are...
Figure 13. Size-class distribution of adult river birch in a 2.3-hectare ar...
Figure 14. Frequency distributions of (A) tree diameters at 1.5 meters abov...
Figure 15. Elevations, ring-count ages, and age-frequency distribution of o...
Figure 16. Departures from 1900-1994 mean annual precipitation for (top) na...
Figure 17. Water-balance model of Sinking Pond showing simulated hydrologic...
Figure 18. Schematic diagram of a parameterized water-balance model of Sink...
Figure 19. (A) Observed and simulated stage, (B) surface-water outflow, and...
Figure 20. Observed and simulated stage of Sinking Pond for verification pe...
Figure 21. Simulated stage for Sinking Pond, January 1854 through September...
Figure 22. Days per water year from 1855-2002, in which simulated stage at ...
Figure 23. Temporal patterns in Sinking Pond hydroperiod, based on centered...
Figure 24. Timing and elevation of simulated annual 200-day inundation comp...
Figure 25. Frequency distributions for recruitment inundation (5-year media...
Figure B1. Photographs showing vegetation and topography of (A) the Sinking...
Figure B2. Changes in land cover in the area drained by the Sinking Pond ou...
Figure B3. Water levels in monitored wells and surface-water stations durin...
Table 1. Height and diameter criteria for tree-size classes.
Table 2. Aggregate density of overcup oak seedlings in Sinking Pond by coho...
Table 3. Aggregate percent survival of overcup oak seedlings in Sinking Pon...
Table 4. Recruitment and survival of overcup oak saplings in sampled plots ...
Table 5. Aggregate density of overcup oak adults by size class and ponding ...
Table 6. Density of adult trees by species and ponding-depth class in a 2.3...
Table 7. Density and percentage of 4-square-meter subplots containing overc...
Table 8. Station data for climate stations used to develop input data set f...
Table 9. Names, units, and calibrated values of constant terms in parameter...
Table 10. Gage data for surface-water stations used in development, calibra...
Appendix A. Flora of Sinking Pond.The following table lists plants observed...
Table B1. Sources and dates of aerial photographs used in land-use analysis...
Table B2. Land-use/land-cover classes used in land-use analysis of the Sink...
1 U.S. Geological Survey
2 University of the South
3 University of Michigan
| Multiply | By | To obtain |
|---|---|---|
| millimeter (mm) | 0.03937 | inch |
| centimeter (cm) | 0.3937 | inch |
| basin centimeter (bcm) | 0.3937 | basin inch |
| meter (m) | 3.281 | foot |
| kilometer (km) | 0.6214 | mile |
| square kilometer (km2) | 0.3861 | square mile |
| Hectare (ha) | 2.471 | acre |
| cubic meter (m3) | 35.31 | cubic foot |
| cubic meter per second (m3/s) | 35.31 | cubic foot per second |
Temperature in degrees Celsius (°C) can be converted to degrees Farenheit (°F) as follows:
°F = 1.8 x °C + 32
Vertical coordinate information is referenced to the National Geodetic Vertical Datum of 1929 (NGVD 29); horizontal coordinate information is referenced to the North American Datum of 1927 (NAD 27).
| AAFB | Arnold Air Force Base |
| AEDC | Arnold Engineering Development Center cdf cumulative distribution function |
| DBH | diameter at 1.5 meter above land surface |
| DOQ | digital orthophoto quadrangle |
| ET | evapotranspiration |
| G1 | globally extremely rare |
| GCM | global circulation model |
| GIS | geographic information system |
| GPS | global positioning system |
| L3/t | volume divided by time |
| NAVD 29 | North American vertical datum of 1929 |
| NCDC | National Climatic Data Center |
| RTE | rare, threatened, or endangered |
| USGS | United States Geological Survey |
| USHCN | United States Historical Climatology Network |
| TVA | Tennessee Valley Authority |
Multiple lines of evidence point to climate change as the driving factor suppressing tree regeneration since 1970 in Sinking Pond, a 35-hectare seasonally flooded karst depression located on Arnold Air Force Base near Manchester, Tennessee. Annual censuses of 162-193 seedling plots from 1997 through 2001 demonstrate that the critical stage for tree survival is the transition from seedling to sapling and that this transition is limited to shallow (less than 0.5 meters) ponding depths. Recruitment of saplings to the small adult class also was restricted to shallow areas. Analysis of the spatial and elevation distribution of tree-size classes in a representative 2.3-hectare area of Sinking Pond showed a general absence of overcup oak saplings and young adults in deep (ponding depth greater than 1 meter) and intermediate (ponding depth 0.5-1 meter) areas, even though overcup oak seedlings and mature trees are concentrated in these areas.
Analysis of tree rings from 45 trees sampled in a 2.3-hectare spatial-analysis plot showed an even distribution of tree ages across ponding-depth classes from the 1800s through 1970, followed by complete suppression of recruitment in deep and intermediate areas after 1970. Trees younger than 30 years were spatially and vertically concentrated in a small area with shallow ponding depth, about 0.5 meter below the spillway elevation. Results of hydrologic modeling, based on rainfall and temperature records covering the period January 1854 through September 2002, show ponding durations after 1970 considerably longer than historical norms, across ponding-depth classes. This increase in ponding duration corresponds closely with similar increases documented in published analyses of streamflow and precipitation in the eastern United States and with the suppression of tree regeneration at ponding depths greater than 0.5 meter indicated by tree-ring analysis. Comparison of the simulated stage record for Sinking Pond with the ages and elevations of sampled trees shows that prolonged (200 days or more per year) inundation in more than 2 of the first 5 years after germination is inversely related to successful tree recruitment and that such inundation was rare before 1970 and common afterwards.
Wetlands in karst (bold terms can be found in Glossary) landforms are widely distributed in the upland valleys and plateaus of the southeastern and south-central United States. A recurrent theme in the literature on these wetlands is the presence of disjunct populations of northern or coastal plain plants and animals found far from their normal range (Barclay, 1957; Greear, 1967; DeSelm, 1981; Ellis and Chester, 1989; Wolfe, 1996b). The disjunct biota of southern karst wetlands contribute to regional biodiversity and may provide insights into the resilience and stability of wetland ecosystems in the face of environmental change (Barclay, 1957; Greear, 1967; Tryon, 1990).
Numerous studies describe biota in southern karst wetlands (Barclay, 1957; Greear, 1967; DeSelm, 1981; Ellis and Chester, 1989; Wolfe, 1996b; Tryon, 1990; McCarthy and Evans, 2000), but few of these studies combine ecological observations with detailed measurements of surface flooding, soil saturation, and ground-water levels (Hendricks and Goodwin, 1952; Greear, 1967; Wolfe, 1996a, b). Fewer still have undertaken long-term monitoring of hydrologic and ecological conditions or attempted to track their interaction through time.
One karst wetland that has received interdisciplinary scientific attention for more than a decade is Sinking Pond (Patterson, 1989; Wolfe, 1996a, b; McCarthy and Evans, 2000). Sinking Pond is a seasonally ponded karst depression whose interior is dominated by overcup oak (Quercus lyrata), a Coastal Plain tree endemic to bottomland hardwood forests (Wolfe, 1996a; McCarthy and Evans, 2000). Sinking Pond is located on Arnold Air Force Base (AAFB), a U.S. Air Force reservation occupying about 160 square kilometers (km2) on the divide between the Duck River and Elk River watersheds, near Manchester and Tullahoma, Tennessee (fig. 1). Sinking Pond and other natural areas provide a sight and sound buffer around Arnold Engineering Development Center (AEDC), a research and development facility operated by the U.S. Air Force at AAFB.
Within Sinking Pond, overcup oaks are concentrated in the deepest areas subject to the longest annual inundation (Wolfe, 1996a). This concentration in deep areas reflects the high tolerance of overcup oak to inundation relative to other bottomland hardwood trees (Broadfoot and Williston, 1973; Solomon, 1990). Clearly, in the past, hydrologic conditions in the deep interior of Sinking Pond were suitable not only for the survival of mature overcup oaks but also for their reproduction and replacement by younger trees. The near absence of overcup oak saplings and young adults in the interior of Sinking Pond (McCarthy and Evans, 2000) suggests that hydrologic conditions have changed in a way that has made formerly suitable sites unsuitable for tree regeneration.
Numerous studies show that regeneration and survival of overcup oak and other bottomland hardwoods are affected by environmental variables related to the hydrologic regime (Hosner and Boyce, 1962; Gill, 1970; Broadfoot and Williston, 1973; Harms and others, 1980; Reily and Johnson, 1982; Hook, 1984; Schupp, 1995; Wada and Ribbens, 1997). Such variables include basic climatic factors, such as precipitation and temperature, human influences ranging from direct hydrologic alteration of wetlands to indirect effects of land use in nearby uplands (Hewlett and Hibbert, 1961), and topographic effects on the timing, frequency, depth, and duration of inundation (Huenneke and Sharitz, 1986; Streng and others, 1989; Hall and Harcombe, 1998).
In 1999, the U.S. Geological Survey (USGS), in cooperation with the U.S. Air Force and the University of the South, began a 3-year study to examine the relation between hydrologic change and regeneration of overcup oak and other species in and around Sinking Pond. The main objectives of the study were to:
1. Determine the nature, extent, timing, and proximate cause of changes in tree regeneration and recruitment patterns in and around Sinking Pond; and 2. Reconstruct the historical ponding regime of Sinking Pond and evaluate climate and human activities as factors driving hydrologic change.This report describes an analysis of the interaction between hydrology and tree regeneration in Sinking Pond. The analysis includes: (1) field observations of germination, recruitment, and mortality of overcup oak and other tree species; (2) analysis of the spatial relations among seedling, saplings, adult trees, and elevation; (3) tree-ring analysis, mapping, and topographic surveying to determine tree-age distribution among overcup oaks in relation to elevation; (4) development and analysis of a 154-year time series of daily temperature and precipitation; (5) development, calibration, verification, and application of an empirical model of water level in Sinking Pond as a function of historical rainfall and temperature; and (6) evaluation of the effects of climate change on tree recruitment and survival during the past 150 years. Additional activities, described in Appendix B, included field observations, air-photo analysis, and water-level monitoring to evaluate possible human influences on the hydrology of Sinking Pond.
Patterson (1989) mapped soils and vegetation in a 650-hectare (ha) area around Sinking Pond, noting high variability of vegetation on wetland soils and a strong correlation between moss lines and soil indicators of flooding. Wolfe (1996a, b) monitored surface-water and ground-water levels in and around Sinking Pond and other karst depressions on AAFB and related tree-distribution patterns to elevation and hydroperiods in the Sinking Pond drainage basin. Wolfe (1996a, b) documented the dominance of overcup oak (Quercus lyrata), a Coastal Plain disjunct, in the deepest parts of Sinking Pond, the absence of sub-canopy plants, including overcup oak saplings, and widespread germination and mortality of overcup oak seedlings in the interior of Sinking Pond. Wolfe speculated that oaks regenerate during infrequent periods of prolonged (several years) drought, producing a population composed of distinct age cohorts.
McCarthy and Evans (2000) inferred the population age distribution of overcup oaks in Sinking Pond by coring 16 adult overcup oaks, relating size to age, and measuring more than 300 additional trees. They concluded that prior to the 1960s, age distribution was relatively uniform, with no distinct cohorts, but that afterwards, regeneration and recruitment of overcup oak had virtually ceased. McCarthy and Evans (2000) speculated that conditions in the pond had become sufficiently wetter to preclude regeneration where it had formerly occurred. However, the cause and mechanism of such a wetting trend remained unidentified, and the absence of hydrologic records for Sinking Pond prior to 1992 left the existence of such a trend open to question.
The results of previous studies raise several questions. What is the age distribution of the overcup oak population, and how is it related to elevation and ponding regime? What are current patterns of germination, recruitment, and mortality of overcup oak and other tree species in Sinking Pond? How have these patterns changed? Have human activities or climate change altered the ponding regime of the pond?
The study was conducted in and around Sinking Pond, a seasonally ponded karst depression with a maximum flooded area of about 35 hectares (ha). Sinking Pond drains a basin of about 335 ha in the northwestern part of Arnold Air Force Base (figs. 1 and 2). The study area lies in the Highland Rim subsection of the Interior Low Plateau Physiographic Province (Fenneman, 1938) near the southern limit of The Barrens, a broad area of low-relief karst extending about 100 kilometers (km) north from the Elk River (fig. 1). The Barrens is notable for supporting numerous plants and animals listed as rare, threatened, or endangered in Tennessee (Svenson, 1941; DeSelm and others, 1974; Kral, 1973; Bowen and Pyne, 1995), including several that are Coastal Plain endemics for which The Barrens and analogous areas of Tennessee and Kentucky are the only significant inland occurrences (Pyne, 2000). The northern part of AAFB represents a major concentration of State-listed Barrens taxa as well as many Coastal Plain disjuncts, such as overcup oak in Sinking Pond (Pyne, 2000).
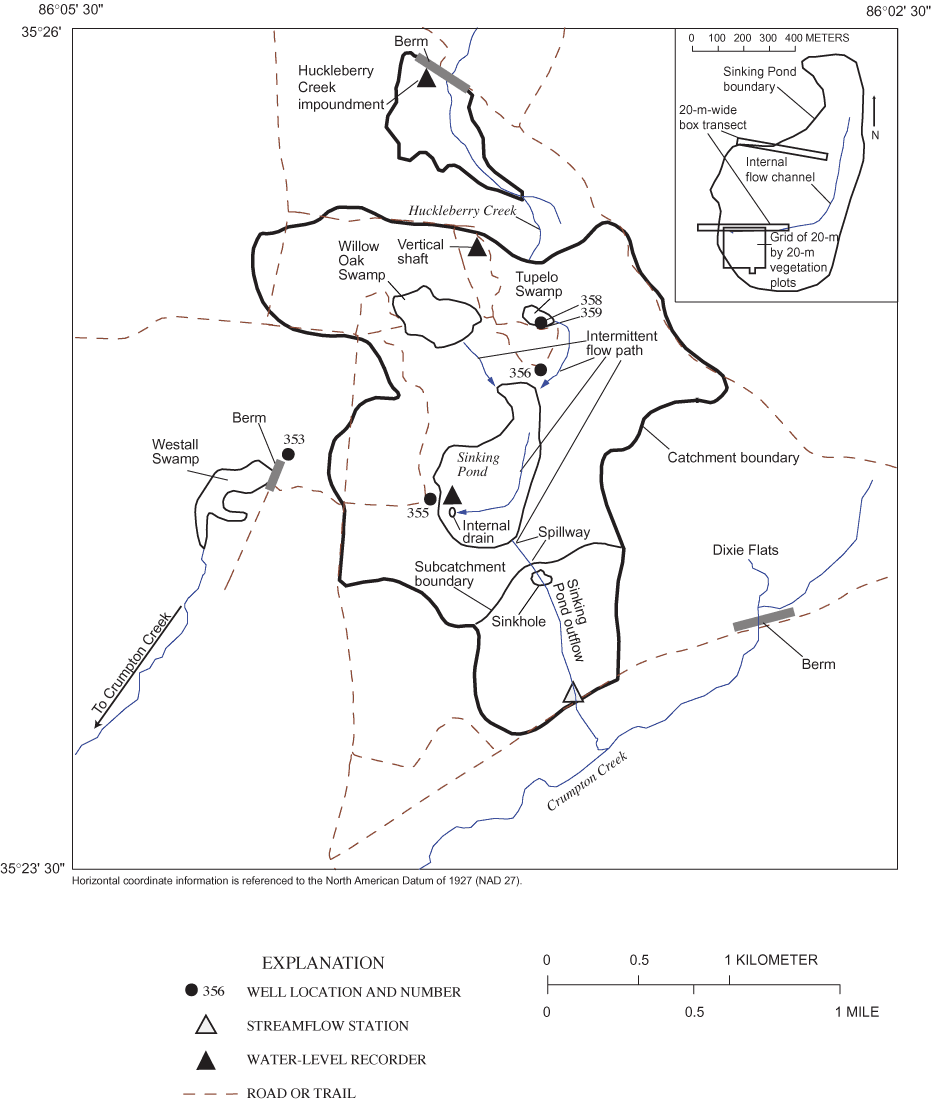
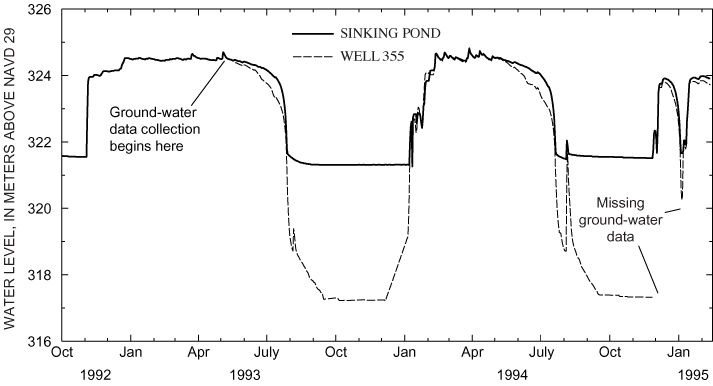
The name "Sinking Pond" dates from the 1830s (Jack Bennett, Archeological Assessments, Inc., written commun., 1999) and reflects the pond's abrupt seasonal rises and recessions (Wolfe, 1996a). The pond generally fills in the fall or winter and remains full for several months before receding in the summer. The seasonal rise can be rapid—commonly 2 meters (m) or more from a fully drained condition in 1 or 2 days (fig. 3).
The distinctive hydrologic behavior of Sinking Pond is largely a result of the pond's karstic origin and its geomorphic expression. The ponded area of Sinking Pond comprises about 10 percent of the area of the Sinking Pond drainage basin, but the pond's 3.5 m of internal relief represents more than 30 percent of the overall relief in the basin (about 11 m). More than two thirds of Sinking Pond's internal relief is contained in a complex of coalesced sinkholes and connecting channels that compose the pond's internal drainage system. Several of these sinkholes receive concentrated recharge in the weeks preceding the seasonal rise and throughout the recession (Wolfe, 1996a, b).
In addition to Sinking Pond, deep (greater than 2.5 m), complex sinkholes with efficient internal drains are found in Willow Oak Swamp, an 80-ha sub-basin northwest of Sinking Pond, in Westall Swamp, a seasonally flooded depression draining 150 ha located west of the Sinking Pond drainage basin, and along the intermittent channel that carries overflow from Sinking Pond to Crumpton Creek, a tributary of the Duck River (Wolfe, 1996a; fig. 2). Wolfe (1996a, b) noted fresh karst collapse features in the Sinking Pond area.
Karst development in the Sinking Pond area is more active, and local relief higher, than is typical of The Barrens. More typical are numerous shallow (1.5 m or less) sinkholes without visible internal drains (Wolfe, 1996a). One shallow, perched sinkhole, Tupelo Swamp, is a sub-basin of Sinking Pond, draining an area of about 13 ha north of Sinking Pond (fig. 2). Other shallow depressions drain small areas on the broad, flat ridges.
The bedrock geology of the Eastern Highland Rim is dominated by limestones and interbedded cherts and shales of Mississippian age that dip gently to the east, away from the Nashville Basin, a structural dome, and towards the base of the Cumberland Plateau. From top to bottom, the major Mississippian-age strata are the St. Louis Limestone, the Warsaw Limestone, and the Fort Payne Formation, which form the Highland Rim aquifer system. The lower boundary of the Highland Rim aquifer system is the Chattanooga Shale of Upper Devonian/Lower Mississippian age (Wilson, 1976; Burchett, 1977), a regional confining unit (Brahana and Bradley, 1986a, b; Haugh and Mahoney, 1994; Wolfe and others, 1997). In general, the Mississipian strata are massive, thickly bedded, and contain a high percentage of calcium carbonate with less insoluble material near the upper units of the St. Louis Limestone and become progressively thinner-bedded and more heterogeneous, with a higher proportion of insoluble material, with increasing depth.
The gentle dip of the strata and their compositional and textural trends down the stratigraphic column have produced distinctive topographic and hydrologic patterns across the surface of the Eastern Highland Rim. Near the base of the Cumberland Plateau, a sinkhole plain is developed on thick outcrops of the upper St. Louis Limestone. Karst development and sinkhole density decrease westward as the St. Louis Limestone thins out, eventually grading into the subtle, low-relief karst of The Barrens, which has developed on the lower Warsaw Limestone and upper Fort Payne Formation. Farther west, along the margins of the Nashville Basin, the mechanical resistance of the lower Fort Payne Formation contributes to the formation of the steep, fluvially disected Highland Rim Ecarpment (Wolfe, 1996a, b; Wolfe and others, 1997).
Most of the AAFB area is mapped as Warsaw and St. Louis Limestones (Wilson, 1976). In the study area, both formations are weathered to clay-rich residuum. The uppermost unit of relatively unweathered bedrock is the Fort Payne Formation.
The primary aquifers in the study area are, from top to bottom, the shallow aquifer, the Manchester aquifer, and the Fort Payne aquifer. The shallow aquifer consists of 1.5 to 23 m of clay-rich residuum and includes the soil cover and root zone. The Manchester aquifer is formed from the weathering of the lower Warsaw Limestone and the Fort Payne Formation (Burchett and Hollyday, 1974) and is the most productive and most complex aquifer in the study area. The upper part of the Manchester aquifer consists of chert gravel, weathered limestone, and rubble. The lower part includes fractures and solution openings in the Fort Payne Formation. The Fort Payne aquifer consists of that part of the Fort Payne Formation which is relatively dense, with few small fractures or solution openings. The characteristics that define the Fort Payne aquifer also limit its transmissivity and productivity (Haugh and Mahoney, 1994).
Solution openings are most common near the top of bedrock in the Fort Payne Formation but have been found 25 m or more below the top of bedrock (Haugh and others, 1992). The lower part of the Manchester aquifer is more prevalent in areas, such as the northern part of AAFB, where weathering profiles are relatively shallow (less than about 15 m thick) and the concentration of solution openings in the Fort Payne Formation is relatively high (Haugh and others, 1992; Haugh and Mahoney, 1994); the distinctive karst topography of the Sinking Pond area is a surface expression of these hydrogeologic characteristics (Wolfe, 1996a).
Soils in the Sinking Pond area belong to the Dickson-Mountview-Guthrie soil association and consist chiefly of Ultisols developed on a thin (about l.5 m), silty mantle overlying cherty limestone residuum (Love and others, 1959; Springer and Elder, 1980; Smalley, 1983; Patterson, 1989). The Dickson silt loam and Mountview silt loam are the dominant soils on well-drained slopes and ridges. Both of these soils are strongly to very strongly acid, moderately permeable in their surface horizons, and low in fertility; they differ primarily in that the Dickson soil has a discontinuous fragipan at the base of the silty upper mantle (Love and others, 1959).
The Guthrie and Purdy silt loams are characteristic soils of headwater wetlands in The Barrens and dominate the ponded area of Sinking Pond. These soils are developed on parent materials similar to those of the Dickson and Mountview soils. The main distinction between them is the higher clay content of the Purdy silt loam. These soils are strongly to very strongly acid and low in fertility. The Guthrie and Purdy silt loams differ from the Dickson and Mountview silt loams primarily in their poor drainage and landscape position (Love and others, 1959; Springer and Elder, 1980; Patterson, 1989). Other soils within the Dickson-Mountview-Guthrie soil association are the moderately well-drained Sango silt loam and the somewhat poorly drained Taft (formerly Lawrence) silt loam (Love and others, 1959; Patterson, 1989).
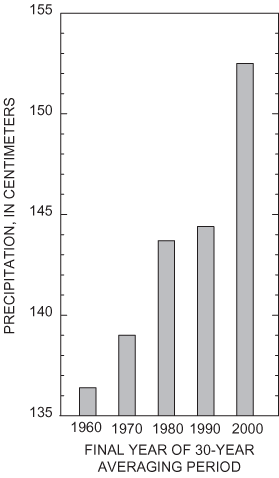
Long-term weather records for Tullahoma, Tennessee, near the southwest boundary of AEDC, are representative of conditions in the study area. Normal (30-year average computed decennially) annual precipitation for the period 1971 through 2000 is 152.5 centimeters (cm). Monthly normal precipitation ranges from 8.9 cm in August to 17.1 cm in March. Monthly normal mean temperatures range from 2.72 °C in January to 25 °C in July (National Oceanographic and Atmospheric Administration, 1900-2003). The normal precipitation at Tullahoma has not been stationary during the past several decades but has increased steadily since 1960 (fig. 4). This increase is consistent with increased precipitation and streamflow since 1970 documented in much of the eastern United States (Karl and others, 1996; McCabe and Wolock, 2002).
The term "barrens" was used widely by English-speaking settlers east of the Mississippi River to describe areas where trees were sparsely distributed or stunted in growth. In Tennessee, the term was applied to areas of heterogeneous oak-savanna vegetation on the Highland Rim with physiographic characteristics that included: (1) low (less than 20 m) relief, (2) thick, infertile, residual soils, and (3) extensive areas of poor drainage interspersed with well or excessively drained slopes and ridges (Killebrew and Safford, 1874; Lollar, 1924; Fenneman, 1938; Wolfe, 1996b; Pyne, 2000). The Barrens (fig. 1) is the largest contiguous occurrence of these areas in Tennessee.
Information on the historical and present-day vegetation of The Barrens and analogous areas of the Highland Rim was reviewed and summarized by Pyne (2000). Fire suppression and other land-use change in the 19th and 20th centuries have altered the vegetative structure and composition of these areas, greatly increasing the forested area and the prevalence of formerly rare or absent tree species, such as fire-intolerant red maple (Acer rubrum L.) and commercially introduced loblolly pine (Pinus taeda L.) (Gattinger, 1901; Lollar, 1924; Chester and others, 1995, 1997; Pyne, 2000). Despite these changes, The Barrens retains considerable botanical importance because of its high concentration of plants and animals listed as rare, threatened, or endangered (RTE) by the State of Tennessee (Svenson, 1941; DeSelm and others, 1974; Kral, 1973; Bowen and Pyne, 1995).
Pyne (2000) tabulated a list of 70 State-listed RTE plant taxa found in The Barrens. A striking characteristic of the list is the high proportion of taxa that are endemic to areas far removed in distance and physiography from the Highland Rim. Of the 70 taxa, 25 are endemic or nearly endemic to the Coastal Plain Physiographic Province—The Barrens and, in some cases, similar areas in Tennessee and Kentucky, appear as prominent and anomalous "inland stations" in their distribution. An additional 15 taxa are broadly disjunct from the Coastal Plain. Seven of the State-listed species of The Barrens are disjunct or peripheral from the northern United States or Canada, and six are disjunct from west of the Mississippi River. Fifteen taxa fit none of the above categories, including several that are widely distributed in the eastern United States and Canada; only two taxa are endemic or nearly so to the Interior Low Plateau (Pyne, 2000). In addition to the 40 State-listed Coastal Plain disjuncts discussed previously, numerous non-listed Coastal Plain plants are found in The Barrens. Two of these, water tupelo (Nyssa aquatica L.) and overcup oak (Quercus lyrata Walt.), are prominent wetland tree species in the Sinking Pond drainage basin.
In the Sinking Pond area, vegetation is generally correlated with topography, drainage, and soil (Patterson, 1989). Well-drained ridges and slopes support deciduous trees such as scarlet oak (Quercus coccinea Muenchh.), southern red oak (Quercu falcata Michx.), and mockernut hickory (Carya tomentosa [Poir.] Nutt.). Moderately well-drained slopes are characterized by white oak (Quercu alba L.), hornbeam (Carpinus virginiana [Mill.] K. Koch), sourwood (Oxydendrum arboreum [L.] D.C.), and yellow poplar (Liriodendron tulipifera L.). The vegetation of poorly drained sites commonly includes sweetgum (Liquidambar styriciflua L.), blackgum (Nyssa sylvatica Marsh.), red maple (Acer rubrum L.), and willow oak (Quercus phellos L.). Some of the wettest sites support stands of coastal plain trees such as overcup oak and water tupelo (Patterson, 1989; Wolfe, 1996a; McCarthy and Evans, 2000).
The deep areas of Sinking Pond are dominated by an assemblage of overcup oak, river birch (Betula nigra L.), and resurrection fern (Pleopeltis polypodioides L.). This assemblage has a preliminary ranking by the Nature Conservancy of G1—extremely rare globally (Tennessee Department of Environment and Conservation, 2002). Appendix A provides a comprehensive list of the flora in the ponded area of Sinking Pond.
This study expands upon the results of McCarthy and Evans (2000) through the development of three lines of botanical evidence: (1) extension of the demographic census of overcup oak in Sinking Pond described in McCarthy and Evans (2000) to better characterize temporal patterns of tree regeneration, recruitment, and mortality with 5 years of continuous monitoring; (2) examination of the relation between elevation—an indicator for ponding depth and duration—and the spatial distribution of different size classes of overcup oak and other tree species in a representative area of Sinking Pond; and (3) analysis of the age-distribution of overcup oak and the relation of germination date to elevation in a representative area of Sinking Pond.
The establishment, survival, and recruitment of overcup oak seedlings and the survival and mortality of adult overcup oaks in Sinking Pond were studied in two 20-m-wide box transects (fig. 2). The box transects were described by McCarthy and Evans (2000), and their center lines correspond to narrower vegetation transects established in a previous study (Wolfe, 1996a). The transects encompass the range of elevation and ponding depth and duration present in Sinking Pond (Wolfe, 1996a).
Seedling establishment and recruitment within the overcup oak population in Sinking Pond was studied using 162 circular seedling plots with a radius of 0.5 m established in 1997 and described by McCarthy and Evans (2000), supplemented by an additional 31 plots established in 1998. The seedling plots were randomly located within two 20-m-wide box transects (fig. 2). The plots were assigned to ponding-depth classes, based on the height above land surface of the watermark (moss line) on the nearest tree. The watermark is a striking discoloration left on trees marking the typical high-water level within the pond (fig. 5), which approximates (plus or minus 15 cm) the spillway elevation, 324.5 m above the National Geodetic Vertical Datum of 1929 (NGVD 29). The distance between the watermark and the land surface was the designated ponding depth for each plot.

Two ponding-depth classes were used in this analysis. "Shallow" plots had a ponding depth of 0.5 m or less; "deep" plots had a ponding depth greater than 0.5 m. Of the original 162 plots, 76 had shallow ponding depths, and 86 had deep ponding depths. The 31 plots established in 1998 all had shallow ponding depths.
Seedlings were defined as individuals less than 0.5 m tall. First-year seedlings were identified by the persistent attachment of the acorn to the stem and recorded. Saplings were defined as individuals 0.5 m to 1.5 m tall at the time of census (table 1). Subsequent seedling cohorts were identified, measured, and tagged annually from 1997 through 2001. The 1997 census identified seedlings from the 1996 age cohort. Each seedling cohort was monitored for survival and growth in all plots for the 5-year period from 1996 through 2001.
Table 1. Height and diameter criteria for tree-size classes.
[m, meters; cm, centimeters; DBH, diameter at 1.5 meters above land surface; --, no criteria; <, less than; ≥, equal to or greater than]
| Size class | Height range (m) | DBH range (cm) |
|---|---|---|
| Seedling | 0-1.499 | -- |
| Sapling | 0.5-0.99 | -- |
| Small adult | >1.5 | <5 |
| Adult | >1.5 | ≥5 |
Seedling density and survival differed between the two ponding depth classes. Seedling density was greater in deep plots than in shallow plots for each of the census years except 1999 (table 2). The 1999 cohort represented germination from a very light seed rain in 1998 that was extremely patchy, particularly in the deep part of the two transects. However, no seedlings from the deep plots survived longer than 3 years past germination (table 3). In comparison, seedlings established in 1997 in shallow areas still had 8.8 percent survival 4 years after germination (table 3).
[Seedling density in seedlings per hectare, determined from 0.5-meter-radius circular plots; Bold numbers represent density of first-year seedlings; n, number of plots; shallow, ponding depth 0.5 meter or less; deep, ponding depth greater than 0.5 meter]
| Cohort year | Census year | ||||
|---|---|---|---|---|---|
| 1997 | 1998 | 1999 | 2000 | 2001 | |
| Shallow | |||||
| 1996 (n = 76) | 26,302 | 11,057 | 5,193 | 3,853 | 3,351 |
| 1997 (n = 76) | 65,002 | 25,623 | 12,062 | 7,706 | 5,696 |
| 1998 (n = 107) | 10,472 | 5,355 | 3,332 | 2,618 | |
| 1999 (n = 107) | 9,163 | 8,211 | 4,998 | ||
| 2000 (n = 107) | 24,989 | 19,277 | |||
| 2001 (n = 107) | 11,542 | ||||
| Deep (n = 86) | |||||
| 1996 | 17,766 | 2,517 | 148 | 0 | 0 |
| 1997 | 67,215 | 5,330 | 148 | 0 | 0 |
| 1998 | 15,545 | 148 | 0 | 0 | |
| 1999 | 1,184 | 296 | 296 | ||
| 2000 | 59,813 | 35,088 | |||
| 2001 | 71,657 | ||||
[Percent survival determined from 0.5-meter-radius circular plots; n, number of plots; shallow, ponding depth 0.5 meter or less; deep, ponding depth greater than 0.5 meter]
| Cohort year | Percent survival at annual census | ||||
|---|---|---|---|---|---|
| 1997 | 1998 | 1999 | 2000 | 2001 | |
| Shallow | |||||
| 1996 (n = 76) | n/a | 42.0 | 19.7 | 14.6 | 12.7 |
| 1997 (n = 76) | 100 | 39.4 | 18.6 | 11.9 | 8.8 |
| 1998 (n = 107) | 100 | 51.1 | 31.8 | 25.0 | |
| 1999 (n = 107) | 100 | 89.6 | 54.5 | ||
| 2000 (n = 107) | 100 | 77.1 | |||
| Deep (n = 86) | |||||
| 1996 | n/a | 14.2 | 0.83 | 0 | 0 |
| 1997 | 100 | 7.93 | 0.22 | 0 | 0 |
| 1998 | 100 | 0.95 | 0 | 0 | |
| 1999 | 100 | 25.0 | 25.0 | ||
| 2000 | 100 | 58.7 | |||
Observed recruitment from seedling to sapling showed a strong relation to ponding-depth class. In ponding depths less than 0.5 m, a pool of saplings was maintained over the 5-year sampling period through successful recruitment of saplings from the seedling pool (table 4). Attrition from this pool was mainly through transition of saplings back into the seedling category through stem dieback and subsequent height loss (table 4). In contrast, no seedlings that grew to saplings in deep plots survived more than 1 year during this study. All individuals that grew from seedling to sapling during the 5-year study were individuals that were already seedlings prior to 1997. The transition from seedling to sapling appears to be the critical point at which the recruitment of new trees currently is failing in the deep areas of Sinking Pond.
[Determined from 0.5-meter-radius circular plots; n, number of plots; --, no data; shallow, ponding depth 0.5 meter or less; deep, ponding depth greater than 0.5 meter]
| Year | Overcup oak saplings | |||
|---|---|---|---|---|
| Total | New | Deaths | Regress to seedling | |
| Shallow plots | ||||
| 1997 (n = 76) | 4 | -- | -- | -- |
| 1998 (n = 107) | 5 | 2 | 0 | 1 |
| 1999 (n = 107) | 3 | 1 | 1 | 2 |
| 2000 (n = 107) | 3 | 0 | 0 | 0 |
| Deep plots (n = 86) | ||||
| 1997 | 14 | -- | -- | -- |
| 1998 | 6 | 0 | 8 | 0 |
| 1999 | 0 | 0 | 6 | 0 |
| 2000 | 0 | 0 | 0 | 0 |
In 1997, 55 seedling plots were randomly selected as center points for 10-m-radius plots in which adult overcup oaks were measured for diameter at 1.5 meters above land surface (DBH), tagged for future reference, and recorded. Survival of established adult trees and recruitment of new adults were tracked in annual censuses of the 10-m-radius plots through 2001.
Overall density of adults was more than five times greater in the deep areas compared to the shallow areas (table 5). Throughout the 5-year period, there was a complete absence of small adults (trees taller than 1.5 m with DBH less than 5 cm) in the deep plots (table 5). This pattern was consistent with the seedling recruitment failure previously described and with the conclusion reached by McCarthy and Evans (2000) that seedlings currently are unable to become saplings and small adults at ponding depths greater than 0.5 m in Sinking Pond. In contrast, limited recruitment of small adults was documented in 2000 and 2001 in shallow areas of the pond (table 5) associated with light openings in the canopy (McCarthy and Evans, 2000). For individuals greater than or equal to 5-cm diameter in both the deep and shallow areas, there were no other gaps in size-class distribution, suggesting continuous recruitment historically in these areas (McCarthy and Evans, 2000).
[Determined from 55 circular plots (radius 10 meters); density in stems per hectare; shallow, ponding depth 0.5 meter or less; deep, ponding depth greater than 0.5 meter; DBH, diameter at 1.5 meters above land surface; cm, centimeters; <, less than; >, greater than]
| DBH, in cm | Number of adult overcup oaks, by year | ||||
|---|---|---|---|---|---|
| 1997 | 1998 | 1999 | 2000 | 2001 | |
| Shallow | |||||
| <5 | 0 | 0 | 0 | 7 | 9 |
| 5-40 | 25 | 25 | 17 | 17 | 17 |
| >40 | 30 | 30 | 25 | 25 | 25 |
| Total | 55 | 55 | 42 | 49 | 51 |
| Deep | |||||
| <5 | 0 | 0 | 0 | 0 | 0 |
| 5-40 | 205 | 204 | 188 | 184 | 181 |
| >40 | 92 | 92 | 80 | 78 | 76 |
| Total | 296 | 296 | 268 | 262 | 257 |
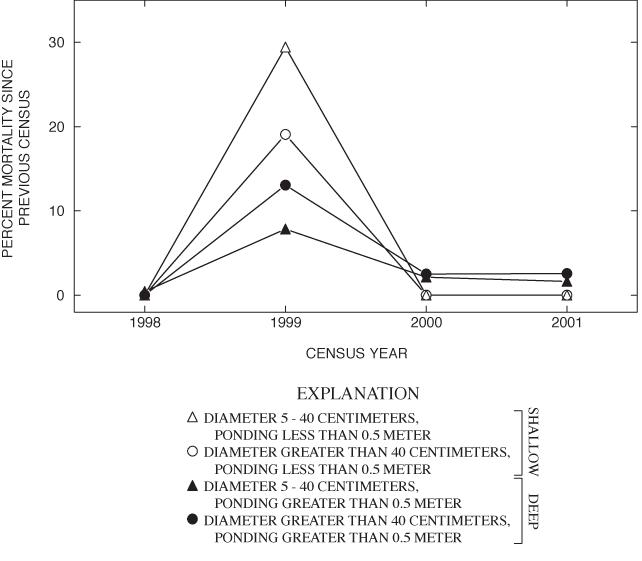
Annual adult-tree mortality between 1997 and 2001 ranged from negligible to nearly 30 percent and varied with size and ponding-depth classes (fig. 6). For all size and ponding-depth classes, the greatest adult-tree mortality occurred in the winter of 1998-99 (fig. 6), during and after ice storms. The ice storms had the greatest effect on trees with DBH of 5 to 40 cm in the shallow areas (fig. 6). Trees in the deep area, however, sustained a slightly higher rate of mortality following the ice storm (fig. 6). The rate of small adult recruitment in the shallow area appeared to compensate for the loss due to mortality (table 5). Such is not the case in the deep area where recruitment was absent. At the current rate of mortality absent replacement, the deep part of Sinking Pond will lose most of its forest canopy by the second half of this century.
The critical stage at which tree-regeneration failure occurs is the transition from the seedling to sapling class. This transition appears to occur only at ponding depths of 0.5 m or less. These results are consistent with earlier findings for overcup oak in Sinking Pond (McCarthy and Evans, 2000). However, generalization of these results to the pond as a whole is limited by the linear sampling framework on which they are based, the imprecision of using watermarks as a surrogate for elevation, and the narrow focus on just one tree species. This section examines tree regeneration and mortality in the context of the overall forest community across a larger and more representative area of the pond.
In August 2001, a marked grid system of plots was established within a 2.3-ha area of forest (fig. 2) to examine spatial patterns of regeneration among the three most common tree species in Sinking Pond: overcup oak, willow oak, and sweetgum. The grid area encompasses the internal flow channel and spans the range of ponding depths from greater than 2 m to the edge of the pond. The grid consisted of 57 plots, 20 m by 20 m, which were divided into 2-m by 2-m subplots.
Within each 2-m by 2-m subplot, the density of overcup oak, willow oak, and sweetgum seedlings, saplings, and small adults were noted. Inside the larger 20-m by 20-m plots, adult overcup oak, willow oak, and sweetgum individuals were measured for DBH and mapped to the nearest meter in grid location. All trees with a DBH of 5 cm or greater were identified to species and mapped with grid location to the nearest meter.
The topography of the grid was surveyed to the nearest centimeter using a total station. Surveyed points included the corners and center of each 20-m by 20-m plot and topographic features such as sinkholes, channels, and major breaks in slope. Geographic coordinates were obtained for five points within the grid using a global positioning system (GPS), and the surveyed points were georeferenced, rectified, and converted into a surface of 2-m by 2-m cells, corresponding to the subplots of the same dimensions.
All subplot and plot data were entered into a spatial database and mapped, using grid coordinates to link them to the rectified surface. For data analyses, the grid was divided into three ponding-depth classes: shallow (less than 0.5-m ponding depth or at or above 324 m above NGVD 29); intermediate (0.5-1.0-m ponding depth or 323.5-323.99 m above NGVD 29); and deep (greater than 1.0-m ponding depth or below 323.5 m above NGVD 29). The distribution of subplots among depth classes was as follows: 26 percent (1,486) were shallow; 35 percent (2,007) were intermediate; and 39 percent (2,207) were deep.
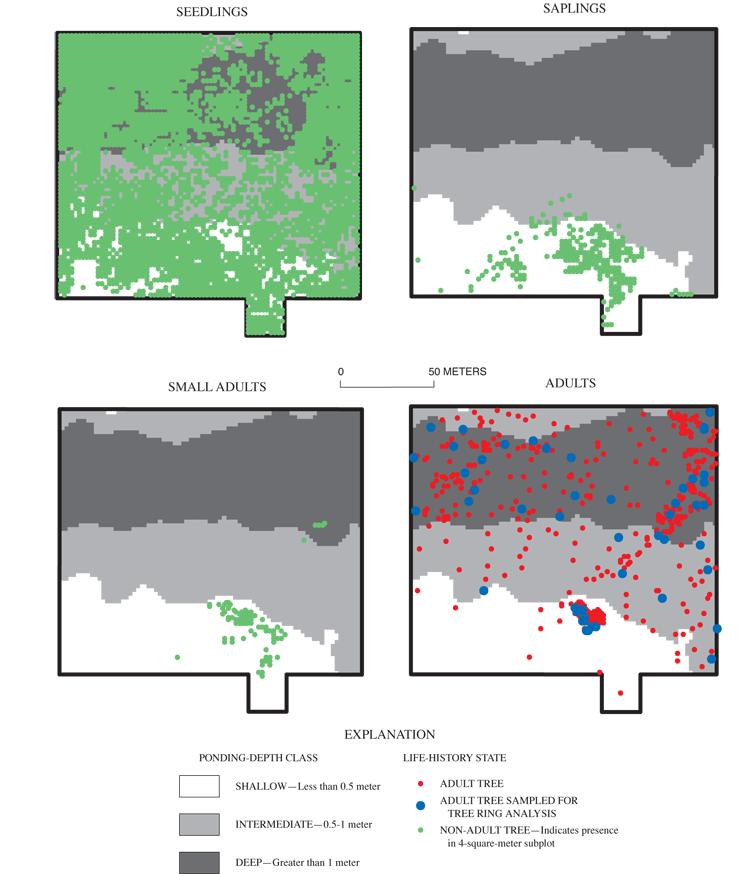
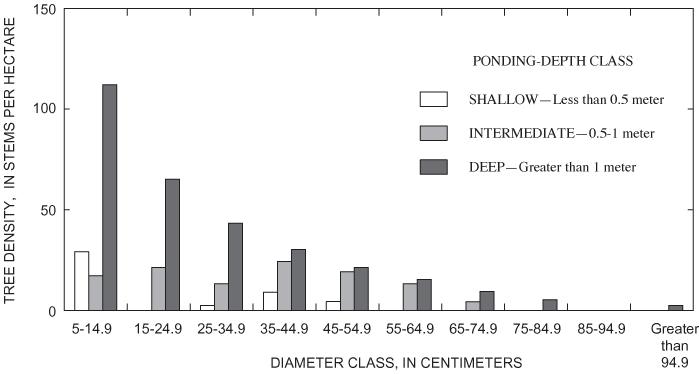
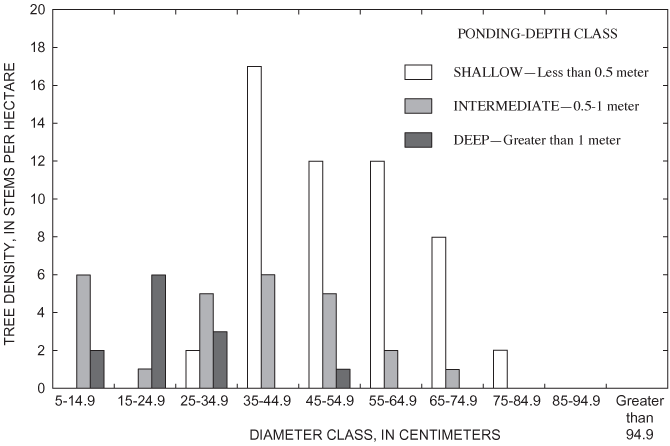
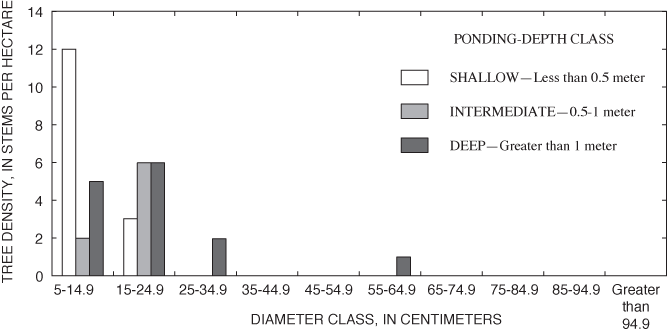
Overall density of adults in the deep area (626 trees per ha) was greater than the density of adults found in the intermediate (324 trees per ha) and shallow (355 trees per ha) areas (table 6). Nine tree species were present within the grid (table 6). All nine species were found in the shallow area, and tree-species richness decreased with increasing ponding depth (table 6). Only overcup oak, sweetgum, willow oak, and river birch composed the canopy of the deep area (table 6). Adult overcup oak density increased with ponding depth (table 6; fig. 7). Density of adult overcup oaks in the deep area was 307 trees per ha, the highest of any species throughout the grid (table 6). The smooth distribution of overcup oak adult size classes for the deep areas (fig. 8) suggests that prior to the recent failure of sapling/small adult recruitment the size distribution of adult overcup oaks in deep areas of the pond was stable. Within each of the depth classes, there was some degree of spatial clumping of overcup oak individuals (fig. 7), as noted by McCarthy and Evans (2000).
[Shallow, ponding depth 0.5 meter or less; intermediate, ponding depth 0.5 - 1.0 meter; deep, ponding depth greater than 1.0 meter]
| Tree species | Tree density (stems per hectare) | ||
|---|---|---|---|
| Shallow | Intermediate | Deep | |
| Sweetgum | 128 | 112 | 22 |
| Red Maple | 74 | 17 | 0 |
| Black Gum | 96 | 26 | 0 |
| Willow Oak | 31 | 22 | 12 |
| Overcup Oak | 28 | 93 | 307 |
| River Birch | 9 | 7 | 14 |
| Sourwood | 3 | 0 | 0 |
| Water Oak | 2 | 0 | 0 |
| Persimmon | 1 | 0 | 0 |
| Total | 626 | 345 | 355 |
Unlike overcup oak, sweetgum density decreased with ponding depth (fig. 9). The density of adult sweetgum in the shallow part of the surveyed area, 128 trees per ha, was the second highest density of adult trees found in this analysis (table 6). Sweetgum was the dominant canopy species in both the shallow and intermediate areas (table 6). The intermediate area was the only part of the sweetgum population that had representation in DBH size classes greater than 55 cm (fig. 10). This pattern, in combination with the high density of individuals having DBH less than 15 cm in the shallow area, suggests a recent population shift of sweetgum to shallower areas within the pond.
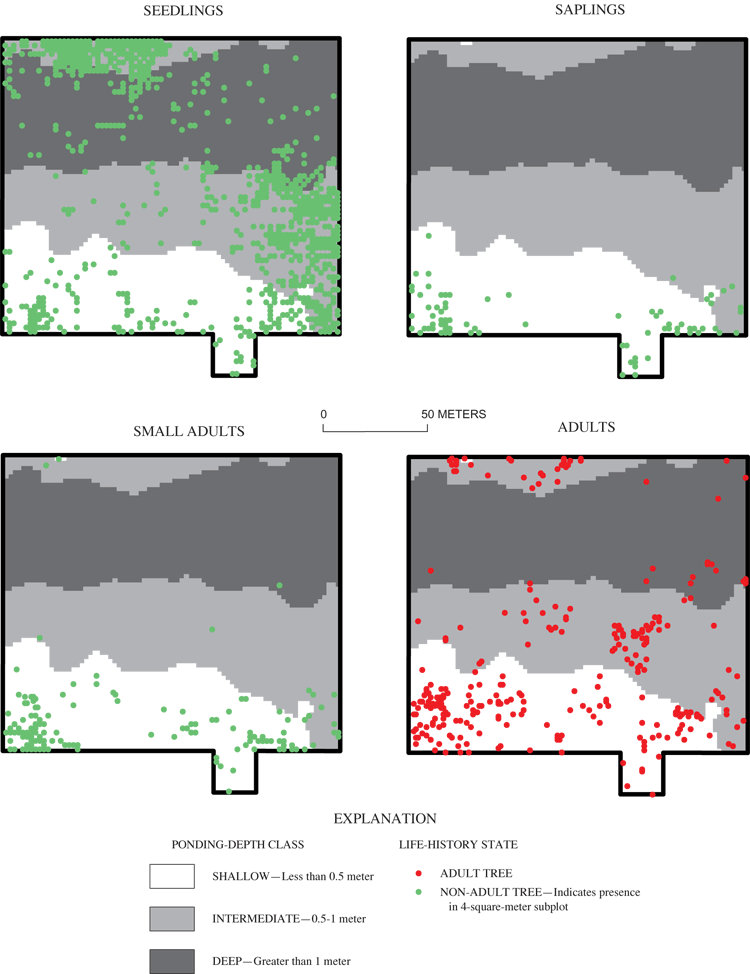
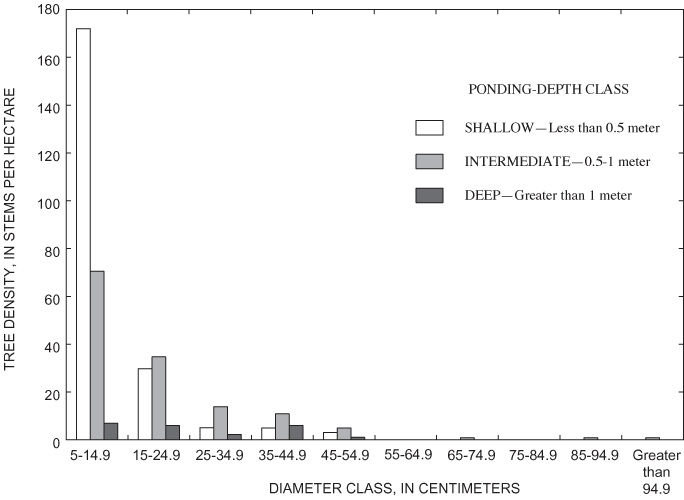
Willow oak had the fourth highest density in each of the ponding-depth classes (table 6). The highest density of Willow oak was noted in the shallow area (table 6, fig. 11). However, the lack of trees in the 5-25 cm DBH class in the shallow area suggests that the population is on the decline in this area (fig. 12). Similar to the other species, recruitment of saplings is strongly concentrated in shallow areas (fig. 11). Recruitment of willow oak small adults appears to be generally failing, even in areas where sapling recruitment is successful (fig. 11).
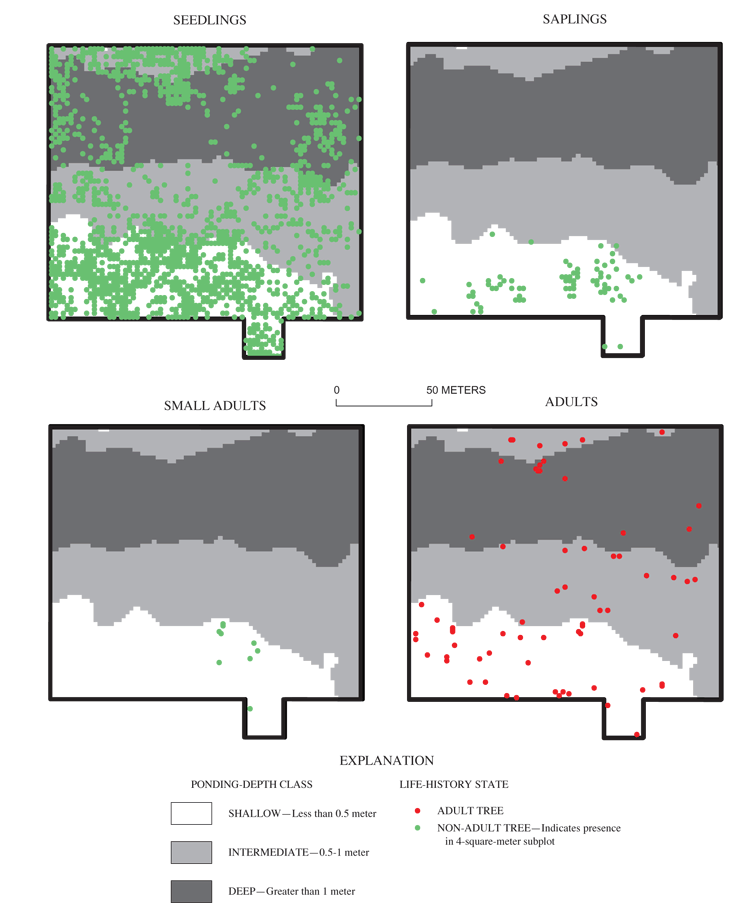
River birch was similar to overcup oak in being the only other species whose density was highest in the deep part of the surveyed area (table 6). The distribution of river birch, like that of overcup oak, is skewed toward smaller size classes in the intermediate and shallow areas, suggesting recent displacement of river birch from deeper to shallower areas of the pond (fig. 13).
Saplings and small adults of all three focal species (overcup oak, sweetgum, and willow oak) were found almost exclusively in the shallow part of the grid (table 7; figs. 7, 9, 11). There were no saplings of any species found in the deep part of the grid. A few small adults were found in deep areas (figs. 7 and 9), but ring counts of recently dead small adult overcup oaks in these areas showed them to be stunted trees older than 30 years, rather than recently recruited. Overcup oak and sweetgum saplings and small adults were present in the intermediate part of the surveyed area in low densities (12 or fewer stems per ha), with less than 0.5 percent of subplots containing these size classes (table 7). The concentration of overcup oak saplings and small adults in shallow areas, the suppression of recruitment to these size classes elsewhere in the pond, and the continued mortality of overcup oak adults discussed in the previous section indicate a spatial shift of the overcup oak population from deep and intermediate areas to shallow areas in Sinking Pond. Few willow oak and sweetgum small adults or saplings were found in the intermediate area (table 7). No river birch saplings or small adults were observed anywhere within the grid. Within the shallow area, saplings and small adults were found in clumped patterns (figs. 7, 9, 11), most likely reflecting the distribution of light gaps in the canopy (McCarthy and Evans, 2000).
[Shallow, ponding depth 0.5 meter or less; intermediate, ponding depth 0.5 - 1.0 meter; deep, ponding depth greater than 1.0 meter; stems/ha, stems per hectare; >, greater than; <, less than]
| Size class and species | Ponding-depth class | |||||||
|---|---|---|---|---|---|---|---|---|
| Shallow | Intermediate | Deep | ||||||
| Density (stems/ha) | Percentage of subplots | Density (stems/ha) | Percentage of subplots | Density (stems/ha) | Percentage of subplots | |||
| Adults | ||||||||
| Overcup oak | 47 | 1.9 | 116 | 4.6 | 307 | 11.7 | ||
| Sweetgum | 215 | 8.6 | 140 | 5.6 | 22 | 0.86 | ||
| Willow oak | 52 | 2.1 | 27 | 1.1 | 12 | 0.45 | ||
| Small adults | ||||||||
| Overcup oak | 106 | 2.6 | 1 | 0.05 | 3 | 0.14 | ||
| Sweetgum | 230 | 6.4 | 12 | 0.45 | 1 | 0.05 | ||
| Willow oak | 10 | 0.34 | 0 | 0 | 0 | 0 | ||
| Saplings | ||||||||
| Overcup oak | 907 | 15.2 | 30 | 0.80 | 0 | 0 | ||
| Sweetgum | 175 | 4.4 | 9 | 0.30 | 0 | 0 | ||
| Willow oak | 156 | 4.4 | 2 | 0.10 | 0 | 0 | ||
| Seedlings | ||||||||
| Overcup oak | 7,206 | 73.5 | >16,000 | 66.8 | >46,000 | 77.4 | ||
| Sweetgum | 390 | 10.8 | 1,668 | 24.6 | 281 | 5.7 | ||
| Willow oak | 2,238 | 47.7 | 1,056 | 24.4 | 430 | 11.6 | ||
Seedlings of all three species were distributed throughout the grid, regardless of water depth (table 7; figs. 7, 9, 11). Overcup oak seedlings were by far the most widespread and numerous, being present in more than 65 percent of the subplots in each of the ponding-depth classes and representing the highest seedling densities of any species (table 7, fig. 7). Overcup oak seedling density increased with ponding depth; total density in the deep area exceeded 46,000 seedlings per ha, nearly triple the density of overcup oak seedlings in the intermediate area and more than six times the density in the shallow part of the sample grid (table 7). Sweetgum seedling density was highest in the intermediate area, and willow oak seedlings were most abundant in the shallow area.
Approximately 10 percent of live adult overcup oak in the 2.3-ha plot grid were sampled for tree-ring analysis to better characterize the age distribution of the overcup oak stand in Sinking Pond. At the time of sampling, information on site topography and tree sizes was insufficient to support a random sample stratified by elevation and size. However, the trees had been mapped, and their spatial distribution provided a framework for initial stratification and sampling.
Adult overcup oaks in the plot grid were divided into five groups based on stem density in different areas of the grid. At least 10 percent of the trees in each group were selected for sampling. The spatial distribution of sampled trees within the population of adult overcup oaks in the plot grid is shown in figure 7. Selection of individual trees was made in the field. The frequency distributions of the elevations and diameters of the sampled trees are similar to those of the live adult overcup oak population in the plot grid (fig. 14). Trees with DBH greater than 75 cm were not sampled because such trees represent less than 2 percent of the plot-grid population (fig. 14), and commonly are hollow in the center. The final sample consisted of 45 of 442 live adult overcup oak in the plot grid.
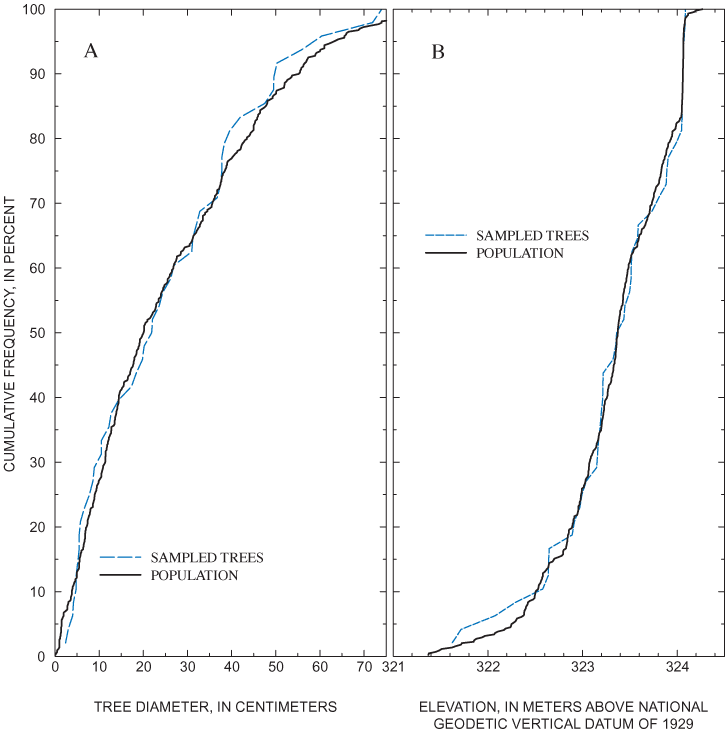
Sample collection, processing, and analysis followed procedures described by Phipps (1985). Trees with diameters of less than 6 cm were sectioned with a crosscut saw, as was one 9-cm tree that had sustained damage from a fallen neighboring tree. All other selected trees were sampled with an increment borer. Cores were mounted in wooden core clamps and shaved with a disposable scalpel. Rings on the mounted, shaved cores were counted using a dissecting microscope for magnification. For the purposes of this study, rings were counted from the outer edge to the center, but individual annual growth increments were not measured.
The completeness and clarity of the rings on each core and section were evaluated, and the probable error associated with each ring count was estimated. Some of the samples failed to include the tree center. In others, the rings were small and indistinct. The maximum number of missing or false rings was estimated for each sample and expressed as an age-estimation error, with values ranging from -8 to 8 years. Positive values indicate less than complete samples with generally clear rings; negative values indicate complete samples with possible false rings. Plus-or-minus values indicate samples with very small, indistinct rings.
For plotting and analysis, the ring-count for each sample was assumed to be the age of the tree, with the age-estimation error as an indication of the probable magnitude and direction of uncertainty associated with each age. Where two or more cores from the same tree yielded different ring counts, the most reliable ring count was used. Ring-count dates were calculated as 2001 minus the ring count. The topographic survey and surface-fitting described in the previous section were used to relate grid location and tree age to elevation.
The results of the tree-ring analysis show a sharp discontinuity in the relation between age and elevation at about 30 years (fig. 15). All trees sampled in shallow areas (elevation above 324 m above NGVD 29) had ring-count ages of less than 30 years. The elevations of young adults revealed by tree-ring analysis thus correspond to the elevation range at which small adult and sapling overcup oaks were found in the spatial analysis of tree-size classes and successful recruitment of saplings observed in the population dynamics study. Ring-count ages among trees sampled in the intermediate and deep areas ranged from 32 to 148 years. The older trees show greatest temporal concentration at about 50 to 65 years (fig. 15); but overall, the results support McCarthy and Evans's (2000) conclusion that the age distribution of overcup oaks in Sinking Pond is relatively even for trees greater than about 30 years, without large gaps.
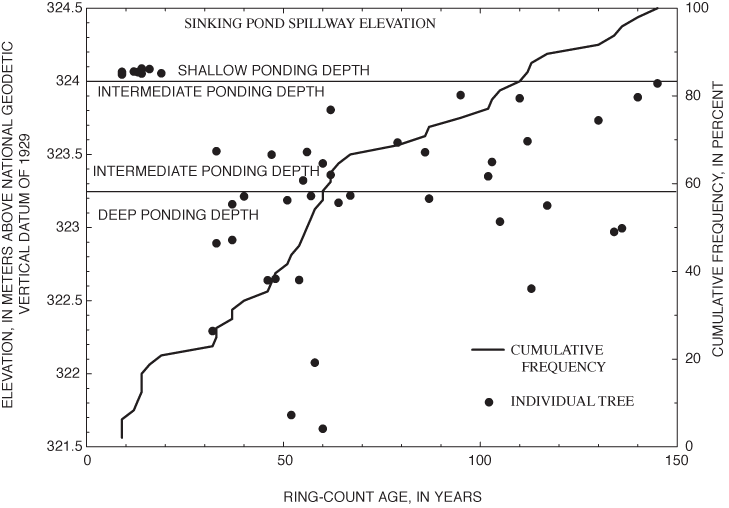
Several lines of botanical evidence suggest that hydrologic change is responsible for the absence of overcup oak saplings and young adults in the deep interior of Sinking Pond. Annual censuses of 162 seedling plots from 1997 through 2001, and of 31 additional plots from 1998 through 2001, show that recruitment to large saplings and small adults is confined to shallow ponding depths, 0.5 m or less below the normal high-water surface. Spatial analysis of tree-size classes for overcup oak, willow oak, and sweetgum shows seedlings and large adults are widely distributed, but saplings and small adults are concentrated in areas with shallow ponding depths. Analysis of tree rings from 45 adult overcup oaks indicates a relatively even age distribution across the pond's elevation gradient for trees dated from 1856 to 1968. Trees dated after 1970 were restricted to shallow ponding depths around 324.2 m above NGVD 29.
These mutually supporting lines of botanical evidence show that: (1) the distribution of overcup oaks relative to ponding depth has changed through time; (2) the most important change is a spatial shift in successful recruitment of saplings and small adults from areas with deep and intermediate ponding depths to areas with shallow ponding depths; and (3) this change happened abruptly beginning around 1970. These observations make a strong circumstantial case that hydroperiods in Sinking Pond have increased during the past several decades and that increase has altered ecological conditions since about 1970. Questions left unaddressed by the botanical evidence include the nature and timing of hydrologic change in Sinking Pond, the cause of such change, and whether it is purely a local phenomenon or reflects a broader regional or global climatic pattern.
Two hypotheses were examined to identify the nature and probable cause of hydrologic change in Sinking Pond. The first hypothesis was that the pond's hydrologic regime had been affected by human activity in and around the Sinking Pond basin. The second hypothesis was that climate change had altered Sinking Pond's seasonal ponding pattern sufficiently to affect tree recruitment and survival.
Direct modification of the pond's spillway or internal drain, land-use and land-cover changes in the pond's drainage basin, and impoundment of surrounding catchments all have the potential to affect hydrologic conditions in the pond. Based on field observations, collection and analysis of hydrologic records, and air-photo analysis, none of these factors explained tree-regeneration patterns and apparent hydrologic change in Sinking Pond (Appendix B). Absent evidence that human activity had altered the hydrology of Sinking Pond, analysis focused on climate change.
Widely reported increases in historical streamflow and rainfall across the eastern United States during the second half of the 20th century (Groisman and Easterling, 1994; Lettenmaier and others, 1994; Karl and others, 1996; Karl and Knight, 1998; Lins and Slack, 1999; Douglas and others, 2000; Easterling and others, 2000; Groisman and others, 2001; McCabe and Wolock, 2002) suggest climate change as an explanation for the apparent increase in the Sinking Pond hydroperiod. For example, McCabe and Wolock's (2002) analysis of daily streamflow records from 400 stations in the conterminous United States for the period 1941-99 revealed distinct step increases around 1970 in annual minimum and median daily streamflow at nearly half (48 percent) of the stations, mostly those located in the eastern United States. Similarly, Karl and others (1996), describing precipitation records for the period 1900-94 from 600 stations distributed across the conterminous United States, noted that "since 1970, precipitation has tended to remain above the 20th-century mean and has averaged about 5 percent more than in the previous 70 years." Precipitation increases of 10 to 30 percent have been documented in the southeastern United States during the 20th century (Burkett and others, 2001), with a regional increase of about 10 percent in Middle Tennessee (Karl and others, 1996).
Temporal patterns in the annual departures from long-term average precipitation at Tullahoma, Tennessee, show strong similarity with national averages (fig. 16) published by Karl and others (1996). The long-term (1900-94) average precipitation at Tullahoma (1,412 mm) is nearly twice the national long-term average of 737 mm, and annual departures from the mean at Tullahoma are proportionately larger than those from the national mean (fig. 16). Both time series show a marked increase in the frequency and magnitude of positive departures and corresponding decreases in negative departures beginning around 1970 (fig. 16).
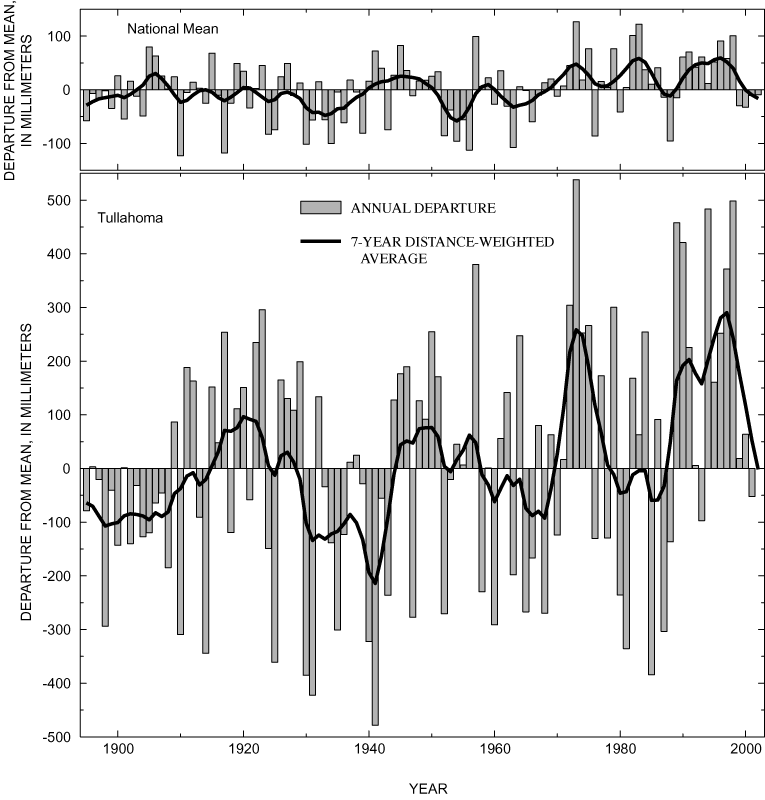
Local and national increases in precipitation and streamflow beginning around 1970 coincide closely with the suppression of overcup oak regeneration in deep and intermediate areas of Sinking Pond shown by tree-ring analysis in the previous section. This similarity in timing suggests a link between climate and changing recruitment patterns in Sinking Pond. Demonstrating such a link requires analysis of the relation between rainfall and the pond's hydrologic conditions. That relation was examined through development of a hydrologic model of Sinking Pond, which used climatic records for input and local hydrologic records for calibration and verification.
Evaluating climate-driven hydrologic change in Sinking Pond required a surrogate measure of the pond's filling and draining behavior over historical time that could be explicitly linked to climate. Long-term monthly and daily climate records for several sites in south-central Tennessee and hydrologic records collected in and around Sinking Pond since 1992 provided a basis for such a surrogate in the form of a simple hydrologic model.
Developing a model required making numerous assumptions, which are described in detail below. The central assumption of the entire modeling exercise is that the hydrologic response of the Sinking Pond basin to climatic inputs has not changed through historical time. The intent of the modeling effort was not to calculate actual water levels in Sinking Pond but rather to evaluate the pond's response to historical temperature and precipitation inputs, holding other independent factors—such as vegetation, human activity, and the hydraulic characteristics of the regional ground-water system—constant. The model had to be able to account for antecedent conditions in the basin, determine when conditions are right for routing water to the pond, and allocate water in the pond to runoff, the atmosphere, or the ground-water system. Additional constraints imposed by limitations of the input and calibration data were:
1. The model had to have the simplest possible structure capable of providing a reasonable representation of the Sinking Pond hydrologic system and an adequate calibration; 2. The number of unconstrained terms could not exceed the number of inflections from distinct storm peaks and seasonal rises and recessions in the calibration data set; and 3. Simulated surface-water discharge from the pond and simulated evapotranspiration had to approximate published local or regional values summed over the calibration period.Daily precipitation and mean temperature data are the inputs for the Sinking Pond hydrologic model. The best available data sets are historical precipitation and temperature records archived by the National Climatic Data Center (NCDC), located in Asheville, North Carolina. The long-term station most representative of the study area is Tullahoma, Tennessee (NCDC station 409155), located about 15 km west of Sinking Pond (fig. 1). The Tullahoma climate data was supplemented by data from several other stations in Tennessee (table 8).
[NCDC, National Climatic Data Center, Asheville, North Carolina; ID, identification; d-m, degrees-minutes; NGVD 29, National Geodetic Vertical Datum of 1929; USHCN, United States Historical Climatology Network; *, station included in the U.S. Historical Climatology Network; D, daily; M, monthly; --, not applicable]
| Station name | NCDC ID number | Latitude (d-m) | Longitude (d-m) | Elevation (NGVD 29) | USHCN | Period of record used | Order of preference | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Daily records | Monthly records | D | M | |||||||
| Tullahoma, Tenn. | 409155 | 35° 21' N | 86° 12' W | 319.4 | * | Apr. 1, 1893 through Sept. 30, 2002 | Apr. 1893 through Sept. 2002 | 1 | 1 | |
| McMinnville, Tenn. | 405882 | 35° 41' N | 85° 48' W | 286.5 | * | Jan. 1, 1927 through Sept. 30, 2002 | Aug. 1872 through Sept. 2002 | 2 | 2 | |
| Lewisburg Experiment Station, Tenn. | 405187 | 35° 27' N | 86° 48' W | 239.9 | * | Jan. 1, 1928 through Sept. 30, 2002 | Apr. 1888 through Sept. 2002 | 3 | 3 | |
| Shelbyville Water Dept., Tenn. | 408246 | 35° 30' N | 86° 29' W | 231.6 | -- | Jan. 1, 1928 through Sept. 30, 2002 | Jan. 1928 through Sept. 2002 | 4 | 4 | |
| Nashville, Tenn. | 406402 | 36° 8' N | 86° 41' W | 182.9 | -- | -- | May 1876 | -- | 5 | |
| Clarksville Sewage Plant, Tenn. | 401790 | 36° 33' N | 87° 22' W | 116.4 | * | -- | Jan. 1854 through July 1872 | -- | 6 | |
| Rogersville, Tenn. | 407884 | 36° 25' N | 82° 59' W | 413 | * | Apr. 1, 1893 through Dec. 31, 1927 | -- | 5 | -- | |
The United States Historical Climatology Network (USHCN), maintained at NCDC, provides relatively complete monthly and daily time series for precipitation and temperature at Tullahoma beginning in 1893 and 1896, respectively. During model development, both time series extended through December 1997. The USHCN monthly data include adjustments intended to improve regional consistency among stations over time (Karl and others, 1996). However, monthly records without regional adjustments were favored for this effort to better represent local hydrologic conditions. Additional climate records for Tullahoma and other Tennessee stations, covering the period from 1998 to 2002, were obtained from NCDC's national network of cooperative weather stations.
Because monthly total precipitation is generally more robust than the set of daily observations, a complete set of monthly values was compiled and used to check and adjust the daily data for modeling. Of 1,314 monthly precipitation totals for Tullahoma between April 1893 and September 2002, 17, or 1.2 percent, were estimated from monthly totals for other Tennessee stations (table 8) using simple linear regression. The station records used as independent variables for the regression estimation were ranked by proximity to the study area, elevation, and completeness of record; the highest ranked station with data for a given month was used to estimate precipitation for that month.
The daily precipitation data set for the period April 1893 through September 2002 consists of 39,994 records. Approximately 97.7 percent of the daily precipitation records were taken from the Tullahoma, Tennessee, daily precipitation data. The remaining 2.3 percent (912 records) were taken from other stations, in order of preference shown in table 8. For months having missing record, the assembled daily precipitation data were summed and uniformly scaled to equal monthly estimated totals.
Monthly and daily time series for mean temperature were developed using the same sources described for precipitation. The procedure for assembling the monthly and daily temperature time series differed from that described above for precipitation in that monthly means, rather than totals, were used.
Synthetic daily precipitation and temperature time series, covering the period January 1854 through March 1893 were generated to extend the hydrologic simulation back to the earliest ring-count dates found in the tree-ring analysis presented above. The synthetic daily time series were based on disaggregated monthly climate records from McMinnville and Clarksville, Tennessee, beginning in 1872 and 1854, respectively. Because of its closer geographic proximity to the study area, the McMinnville monthly record was used whenever available.
Disaggregation of monthly totals was based on the application of a scaled distribution of daily precipitation values adapted from daily observations in a set of 12 typical reference months selected from the observed record (1893-2002). Selection of the reference months was based on two criteria: (1) a complete daily precipitation record for the month (no estimated values), and (2) a total number of rain days within 2 days of the 1893-2002 average for that calendar month. The 12 selected reference months were: January through April 1905, May through September 1904, October 1905, and November and December 1904. Daily values in the collected reference months were used to represent the corresponding daily values for each day in the disaggregated calendar year. Disaggregated record for the period January 1854 through April 1893 was generated by multiplying daily reference precipitation within each month by the ratio of that month's precipitation total to its corresponding reference total.
A synthetic time series of daily mean temperatures for the period January 1854 through March 1893 was generated using methods similar to those described above for precipitation. Because temperature generally varies more smoothly from day to day than precipitation, the 1970-99 daily normal mean temperatures were used as the reference daily temperature values. The 1970-99 daily normal mean temperatures were multiplied by the ratios of the 1854-93 monthly mean temperatures to the corresponding 1970-99 monthly normal mean temperatures to produce the synthetic daily temperature time series.
The starting point for modeling water fluxes into and out of Sinking Pond is a classical water budget in the form:
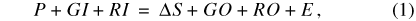
where P is precipitation; GI is ground-water inflow; RI is surface-runoff inflow; ΔS is change in volume of stored water; GO is ground-water outflow; RO is surface-runoff outflow; and E is evapotranspiration (ET). The storage term ΔS has inherent dimensions of volume (L3). The remaining terms in equation 1 are hydrologic fluxes, which can be expressed as volumes by integrating them over a standard time step. For the hydrologic model presented in this report, the standard time step is daily, and water-storage and flux terms have units of basin centimeters (bcm). For the Sinking Pond basin (area 335 ha), 1 bcm equals 33,500 cubic meters (m3).
The first task in developing a water-balance model is to specify the number and definitions of one or more storage compartments and to conceptualize the movement of water into, out of, and between them. For this study, the model must account for precipitation and runoff inputs to the pond and outflows from the pond to surface water, the ground-water system, and the atmosphere. The number of possible approaches for structuring such a model is very large. In addition to the surface water in Sinking Pond, the modeled system could be defined to include such additional compartments as surface-water storage in sub-basins, vadose-zone storage, and shallow or deep ground-water storage beneath different parts of the basin and the pond. Water movement between these compartments could be conceptualized as unidirectional or bi-directional. Water in one compartment might be routed to another compartment or to a sink outside the modeled system.
After consideration of several alternatives, a model form was selected based on two water-storage compartments: (1) water stored in the basin vadose zone, and (2) water stored in Sinking Pond. In terms of equation 1, each compartment receives rainfall, gains or loses stored water, returns water to the atmosphere through evapotranspiration, loses water to the ground-water system, and generates surface-water runoff. Neither compartment receives ground-water inflow. The two compartments are linked by the surface runoff outflow from the basin, which enters Sinking Pond as surface-water inflow. Modifying equation 1 to reflect the particular fluxes and linkage of the two compartments yields water-budget equations for the basin vadose zone:
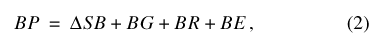
and for Sinking Pond:
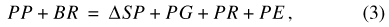
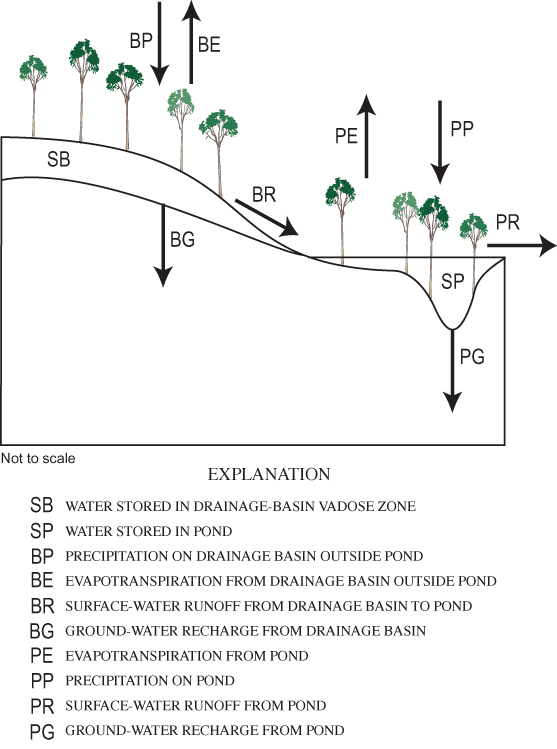
where BP is precipitation falling on the basin outside the pond, SB is the water storage in the basin vadose zone outside the pond perimeter, BG is water flux from the basin vadose zone to ground water, BR is surface-water runoff from the basin to the pond, BE is ET from the basin outside the pond, PP is precipitation falling on the pond, SP is water storage in the pond, PG is water flux from the pond to ground water, PR is surface runoff from the pond, and PE is ET from the pond. A conceptual diagram of the two-compartment water-balance model for Sinking Pond is shown in figure 17.
Equations 2 and 3 provide the basic framework for the model. Using that framework to produce a simulated historical hydrologic record required defining (1) initial and boundary conditions, (2) model parameters, and (3) algorithms to track water storage in the two compartments and the fluxes into, out of, and between them through time. As with the basic model structure, there were many possible approaches to parameterization that might produce acceptable results. Of the several alternatives that were evaluated, the approach described below was the simplest acceptable version, yielding an adequate calibration with the smallest number of terms.
The final model uses the 10 variables from equations 2 and 3. Two of these, SB and SP are state variables (in bcm) that define water storage in the model compartments at any given time; the model's initial conditions are specified by the starting values of SB and SP. Basin storage SB represents water stored in the basin vadose zone, which can either be saturated (water surplus) or unsaturated (water deficit). Conceptually, pond storage, SP, is more complex. Based on previous studies (Wolfe, 1996a, b), Sinking Pond is treated in the model as a karst window—the surface expression of a ground-water reservoir. For SP, water surplus is defined as the condition under which surface runoff is generated, and water deficit is defined as all other conditions. Deficit states for SP encompass all pond stages below the spillway, including fully drained conditions.
The remaining eight terms from equations 2 and 3, BP, BG, BR, BE, PP, PG, PR and PE, are flux variables (in bcm) that represent water movement into, out of, or between the compartments for a given daily time step.
The model utilizes two input terms, temp (degrees Celsius [°C]) and precip (in cm), which are climate data, based on records of daily mean temperature and precipitation, respectively. The input terms are used to determine two forcing functions, PT (in bcm), representing total precipitation, and PET (in bcm), representing maximum potential ET.
In addition to the variables, input terms, and forcing functions, the model includes 10 constant terms or parameters (table 9). Two of the constants are scaling factors used to determine spatial or temporal distribution of input terms or forcing functions. The pond-area scaling factor, PA (dimensionless), is set at 0.1, the ratio of the flooded area of Sinking Pond at spillway stage (34 ha) to the total area of the Sinking Pond drainage basin (335 ha) rounded to one significant figure. The potential ET scaling factor, FE (in bcm per degree Celsius squared [bcm/°C2]), is constrained to a value of 0.00115 bcm/°C2 by the requirement that cumulative PET through the 2-year calibration period approximate the published regional average pan evaporation of 1,200 mm per year (U.S. Geological Survey, 1970, p. 96).
[PET, potential evapotranspiration; bcm/°C2, basin centimeters per degree Celsius squared; cm, centimeter; bcm, basin centimeters]
| Term | Name | Units | Value | Method for determining value |
|---|---|---|---|---|
| PA | Pond-area scaling factor | None | 0.1 | Map analysis and topographic survey |
| FE | PET scaling factor | bcm/°C2 | 0.00115 | Set so the sum of daily PET over calibration period approximates regional average annual pan evaporation rate of 120 cm per year1 |
| BO | Basin overflow threshold | bcm | 0 | Arbitrarily set at zero |
| PO | Pond overflow threshold | bcm | 0 | Arbitrarily set at zero |
| BAV | Basin water-availability threshold | bcm | -23.4 | Spatially and depth-weighted mean published values of available moisture capacities of major soil series in Sinking Pond basin |
| PAV | Pond water-availability threshold | bcm | -9.48 | Spillway volume of pond plus product of BXT multiplied by PA |
| KBR | Basin runoff rate coefficient | None | 0.2 | Manual calibration |
| KPR | Pond runoff rate coefficient | None | 0.5 | Manual calibration |
| KBG | Basin ground-water rate coefficient | None | 0.12 | Manual calibration |
| PG | Pond ground-water constant flux | None | 0.0762 | Manual calibration |
1U.S. Geological Survey, 1970, p. 96.
The model specifies four threshold parameters—specific values of the state variables that represent discontinuities in the behavior of the system. The basin overflow threshold, BO (in bcm), and the pond overflow threshold, PO (in bcm), are values of storage above which surface runoff flows from the basin and the pond, respectively. Both PO and BO were arbitrarily set at zero (table 9), but they differ in physical significance. The model assumes that the hydrologic response of the Sinking Pond drainage basin is dominated by the basin vadose zone, and that most of the basin storage is contained in the soil-root zone. Thus, the basin overflow threshold, BO, represents a soil-moisture condition, rather than a water-surface elevation. In contrast, the pond-overflow threshold, PO, represents spillway stage, with the corresponding volume of surface water in the pond equal to 2.39 x 105 m3 or 7.14 bcm, based on the stage-volume rating. Positive values of SP indicate that pond stage exceeds the spillway elevation, and negative values indicate stage is below the spillway, but do not necessarily mean that the pond is dry.
ET is driven by temperature but limited by the availability of water. The model addresses that limit by two threshold terms, BAV and PAV, which represent the volume of water, in bcm, available for ET in the basin and the pond, respectively. Because they are defined as storage values below overflow conditions, both water-availability thresholds are represented as negative storage terms or deficits. BAV is assumed to be dominated by soil water held in the basin vadose zone. PAV is assumed to include soil water, but it is dominated by surface water in the pond.
BAV was determined from published available water capacities (bcm water per cm soil) of the Guthrie, Purdy, Mountview, and Dickson soil series (Conner, 2000; Jenkins, 2000). Available water capacity is published as a range of values for three or four discrete depth increments to 152 cm below land surface for the Mountview, Dickson, and Guthrie soils (Jenkins, 2000) and to 165 cm below land surface for the Purdy soil (Conner, 2000). The maximum value for each depth increment was multiplied by that increment's thickness (in cm) to yield a high estimate of how much available water the increment could hold. The incremental estimates for each soil were then summed to give a high estimate of maximum available water for areas mantled by the different soil series. The series estimates ranged from 19.8 bcm for the Dickson soil to 30.8 bcm for the Purdy soil. Each series estimate was weighted by the proportion of the total basin area represented by that soil series in the most recent available soil map of AEDC (Edwin Roworth, Arnold Engineering and Development Center, written commun., 2002). The spatially-weighted series estimates were then summed to give the overall value of 23.4 bcm for BAV (table 8).
The pond water-availability threshold, PAV was determined by multiplying BAV (23.4 bcm; table 9) by the pond-area scaling factor (0.1; table 9) and adding the total surface volume for the pond at spillway stage (7.14 bcm from stage-volume rating) to yield the value 9.48 bcm (table 9).
The remaining four constant terms determine fluxes from or between the storage compartments. Three of the terms, K BR, K PR, and K BG, are rate coefficients, which specify the proportionality between storage in a given compartment and the flux leaving that compartment. The basin runoff (K BR) and pond runoff (K PR) coefficients determine runoff from the basin and pond as functions of SB and SP, respectively. The basin ground-water recharge coefficient, K BG, functions in an analogous way to determine ground-water losses from the basin vadose zone as a function of SB. Ground-water flux from the pond was assumed to be constant with a daily magnitude equal to the pond ground-water recharge constant-flux term, PG. Names and units of the constant terms used in the model and the magnitude of and basis for their assigned values are given in table 9.
The model combines 24 terms in 12 equations, of which 6 have 2 or more conditional versions and 6 are unconditional. Using daily precipitation (precip) and temperature (temp) data from climate records as input, the model calculates the 12 equations for each daily time step, i.
The two forcing functions, daily total precipitation and maximum potential ET are calculated by equations 4 and 5, respectively. Daily total precipitation is taken directly from the precipitation input data set:

where PT i is total precipitation for daily time step, and precip is the corresponding precipitation value from the input data set. Daily temperature determines calculation of maximum potential ET (eq. 5):
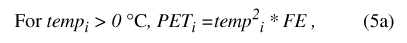
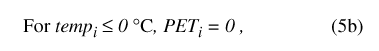
where for each daily time step i, temp i (°C) is daily mean temperature from the input data set, PET i(bcm) is maximum potential ET, and FE (bcm/°C2) is an empirically determined scaling factor (table 9).
Equations 6 through 10 represent routing of water through the Sinking Pond drainage basin. Equation 6 is the basin water-balance equation, rearranged from equation 2:
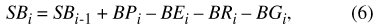
where SB i and SB i-1 represent the volume of water stored in the basin at time steps i and i-1, respectively; BP i is precipitation falling on the basin during time step i; and BE i, BR i, and BG i represent water leaving the basin during time step i as ET, surface runoff, and ground-water recharge, respectively.
Precipitation inputs to the Sinking Pond basin are represented by a simple, unconditional scaling equation:
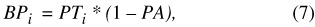
where BP i is precipitation falling on the basin exclusive of the pond during time step i, PT i is total precipitation for time step i (eq. 4), and PA is a dimensionless scaling factor representing the area of Sinking Pond relative to its drainage basin (table 9).
Water movement from the basin to the atmosphere (BE) is represented by one of three conditional expressions (eqs. 8a, 8b, and 8c), depending on the magnitude of basin water storage (SB) relative to the basin overflow and water-availability thresholds (BO and BAV, respectively; table 9). When basin storage equals or exceeds the basin overflow threshold, soil saturation is sufficient to permit PET to be fully realized throughout the drainage basin:
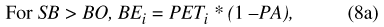
where SB (bcm) is basin storage, BO (bcm) is the basin overflow threshold, BE i (bcm) is ET loss from the basin for time step i, PET i (bcm) is maximum potential ET for time step i, and PA is the dimensionless pond-area scaling factor.
Negative basin storage (SB less than BO) indicates unsaturated soil-moisture conditions. The model assumes that ET continues for SB below BO in proportion to the difference between SB and the basin water-availability threshold BAV as long as SB exceeds BAV:
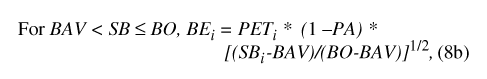
where BAV (bcm) is the basin water-availability threshold and the other terms are as defined previously. Once SB reaches or falls below the basin water-availability threshold, BAV, ET loss from the basin is assumed to cease (eq. 8c, fig. 18).
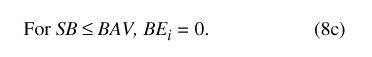
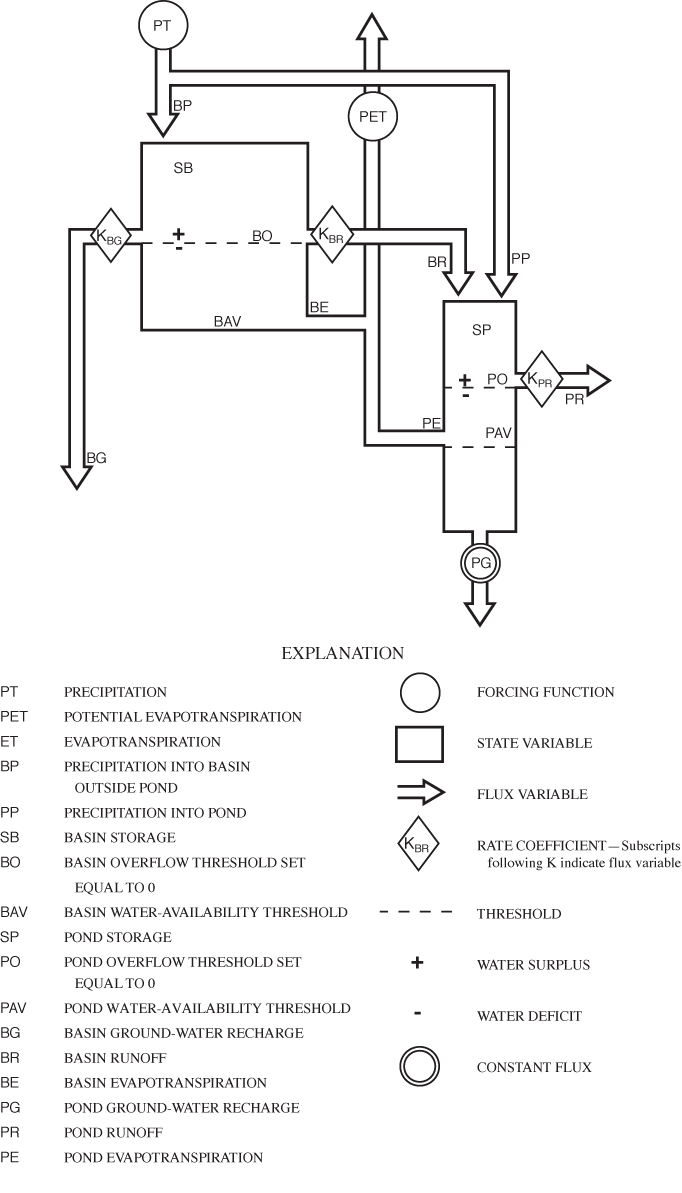
Figure 18. Schematic diagram of a parameterized water-balance model of Sinking Pond.
Surface runoff from the Sinking Pond basin (BR) is represented by either of two conditional equations (eqs. 9a and 9b). When SB exceeds BO, BR is proportional to the difference:
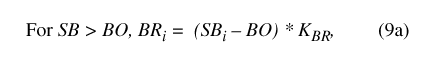
where K BR is a dimensionless coefficient (table 9). When SB equals or falls below the basin runoff threshold BO, the model assumes that runoff losses from the basin are zero:
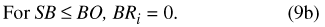
The equations representing ground-water losses from the basin (eqs. 10a and 10b) are identical in form and governing conditions to those representing runoff (eqs. 9a and 9b) discussed above:
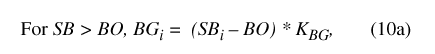
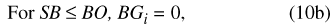
where SB i (bcm) is basin storage at time step i, BO (bcm) is the basin overflow threshold, and K BG is a dimensionless rate coefficient.
The daily water-balance equation for Sinking Pond (eq. 11) incorporates BR as a hydrologic input:
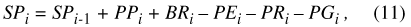
where SP i is the volume of water stored in Sinking Pond at time step i, SP i-1 is pond storage from the previous time step, PP i is the daily precipitation input to the pond, BR i is daily runoff moving from the basin to the pond, PE i is daily ET losses from the pond, PR i is daily runoff losses from the pond, and PG i is daily ground-water losses from the pond. All terms in equation 11 have units of bcm.
Daily precipitation inputs to Sinking Pond are calculated by equation 12:

where PP i (bcm) is precipitation entering the pond during time step i, PT i (bcm) is total precipitation for time step i, and PA is a dimensionless scaling factor (table 9).
Water movement from Sinking Pond to the atmosphere (PE) is represented by one of three conditional expressions (eqs. 13a, 13b, and 13c), depending on the magnitude of pond storage (SP) relative to the pond overflow and water-availability thresholds (PO and PAV, respectively; table 9). When pond storage equals or exceeds the pond overflow threshold, Sinking Pond is full, and maximum potential ET is fully realized for the entire pond area:
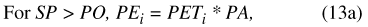
where SP (bcm) is pond storage, PO (bcm) is the pond overflow threshold, PE i is the ET loss from the pond for time step i, PET i is the maximum potential ET for time step i, and PA is a dimensionless scaling factor.
Negative pond storage (SP less than PO) indicates less-than-full conditions. The model assumes that ET continues for SP below PO in proportion to the difference between SP and the pond water-availability threshold PAV as long as SP exceeds PAV:
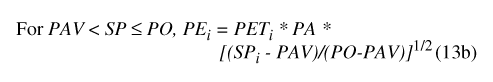
where PAV (bcm) is the pond water-availability threshold, and the other terms are as defined previously. Once SP reaches or falls below the PAV, ET loss from the basin is assumed to cease:
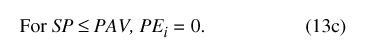
Surface runoff from Sinking Pond (PR) is represented by either of two conditional equations (eqs. 14a and 14b). When SP exceeds PO, calculated PR is proportional to the difference:
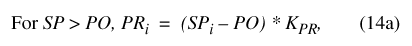
where K PR is a dimensionless coefficient (table 9). When pond storage SP equals or falls below the pond overflow threshold PO, the model assumes that runoff loss from the pond is zero:
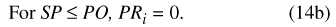
In contrast to surface runoff and ET, the model's treatment of ground-water recharge from Sinking Pond differs from that used for the basin. Previous studies documented the importance of antecedent ground-water conditions to the filling behavior of Sinking Pond (Wolfe, 1996a, b). However, details of the interaction between ground-water conditions and the hydraulic behavior of Sinking Pond are poorly understood. The simplest approach that maintains the interdependence between the local ground-water system and the pond is to assume that ground-water recharge from the pond is constant:

where PG i is ground-water loss from the pond for time step i, and PG is the pond ground-water constant flux in bcm (table 9).
As noted above, Sinking Pond storage (SP) was conceptualized as a karst window comprising surface-water and ground-water components. The model sets no lower limit on the magnitude of SP (fig. 18); deficits (negative values for SP) accumulate below the fully drained condition of the surface pond until reversed by the overall balance of hydrologic inputs and outputs.
Each value for SP has a corresponding value for pond volume and stage, which are calculated at every time step. At SP = 0 bcm, the modeled stage of Sinking Pond is defined to equal the spillway stage of 324.54 m, with a corresponding volume of water in the pond of 2.39 x 105 m3 or 7.14 bcm, based of the stage volume rating. For each time step i, the volume of water in Sinking Pond was calculated as:
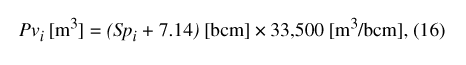
where Pv i is the modeled volume of water in Sinking Pond at time step i, in cubic meters. The modeled stage for each time step was determined from the modeled volume, based on the stage-volume rating.
The calibration data set drew from USGS records of Sinking Pond stage and streamflow in the Sinking Pond overflow channel at a culvert about 0.9 km south and downgradient of the Sinking Pond spillway (fig. 2; table 10). Wolfe (1996a) reported details on construction and instrumentation of these stations. The calibration period, February 11, 1993, through February 12, 1995, covers the period for which continuous data are available for both stations.
[d-m-s, degrees, minutes, seconds; NGVD 29, National Geodetic Vertical Datum of 1929; km2, square kilometer]
| Station name | USGS site identification number | Latitude and longitude (d-m-s) | Station type | Gage datum (NGVD 29) | Drainage area (km2) | Period of record used |
|---|---|---|---|---|---|---|
| Sinking Pond | 03596075 | 35° 24' 36" 86° 04' 11" | Continuous stage | 321.03 | 3.35 | Feb. 11, 1993 – Feb. 12, 1995; Oct. 1, 1999 – Sept. 30, 2002 |
| Sinking Pond outflow | 035960755 | 35° 24' 00" 86° 03' 41" | Continuous streamflow | 308.575 | 3.996 | Feb. 11, 1993 – Feb. 12, 1995 |
Calibration was based on graphical comparison of modeled and observed values for three closely related time series: (1) pond volume, (2) pond stage, and (3) pond runoff. Comparing the observed daily mean discharge at the Sinking Pond outflow gage with PR over the calibration period required converting the daily discharge values from cubic meters per second (m3/s) to daily basin centimeters of runoff. The gage drainage basin area is 399 ha (Wolfe, 1996a), about 15 percent larger than the Sinking Pond drainage basin. Each basin centimeter at the gage represents 39,900 m3 of water. Daily mean discharge in basin centimeters was calculated by multiplying the daily mean discharge by 86,400 seconds per day to determine cubic meters per day and then dividing by 39,900 m3/bcm.
As noted in the previous section and in table 9, values for 6 of the model's 10 constant terms, 2 scaling factors and 4 threshold parameters, were determined independently or assigned arbitrarily, leaving 4 constant terms, K BR, K PR, K BG, and PG, available for use in calibrating the model. The pond runoff coefficient, K PR, was partly constrained by the requirement that cumulative modeled runoff closely approximate the cumulative water yield 51 bcm observed at the Sinking Pond outflow gage during the period (Wolfe, 1996a). Within that constraint, values for the four rate coefficients were determined through an iterative process in which the computed values of daily pond stage, surface-storage volume, and runoff were compared to those of the calibration data set using different values of the various coefficients.
Given the close relation between stage and volume, comparison of the two time series may seem redundant, but the different shapes of the two hydrographs (fig. 19) made them complementary for calibrating the model. The pond-volume hydrograph rises and falls more gradually than the stage hydrograph because volume is determined by the balance of hydrologic inputs and outputs and thus is insensitive to the geometry of the pond. Because the model is based on volumetric water balance, comparison of positive values for observed and modeled volume hydrographs was the main tool used to adjust the general magnitude of the most sensitive parameters, PG, BR, and BG. Observed volume, based on the stage record, has zero as its lower limit, whereas, modeled volume, like the state variable SP on which it is based, can accumulate deficits lower than the fully drained condition of the surface pond (fig. 19). Only positive values for modeled and observed volume were used in calibration.
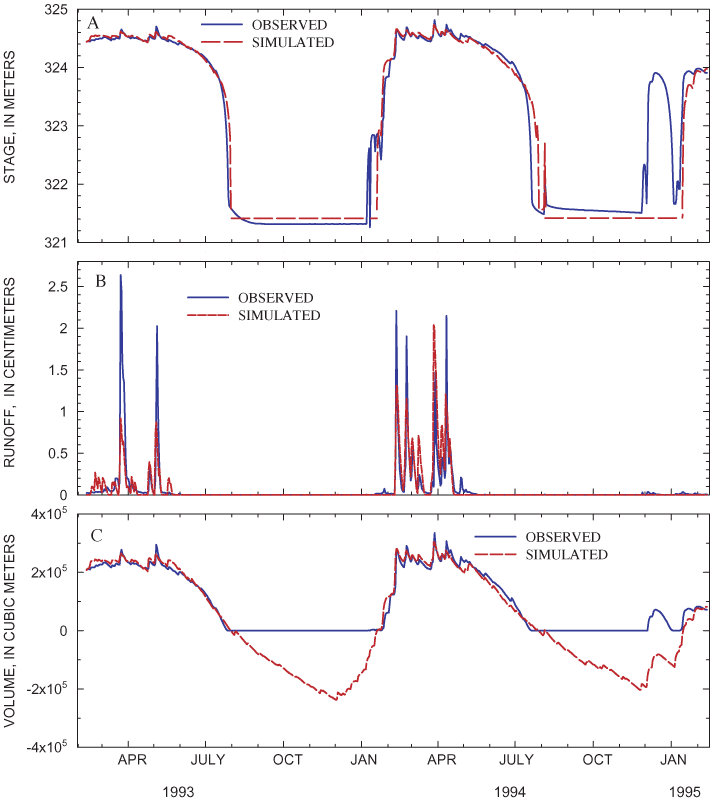
Comparison of the modeled and observed stage and outflow hydrographs were used to adjust BR to match storm peaks. The model produces reasonably good approximations of recessions and wet-season peaks in Sinking Pond for the calibration period, but these approximations tend to lag behind the observed values during early filling (fig. 19). This lag probably reflects the limited information available on ground-water storage directly beneath Sinking Pond and its interaction with the regional ground-water flow system. Adding terms or relaxing some of the model constraints might have reduced or eliminated this early-rise lag, but also would have overspecified the model.
A continuous stage recorder was re-established in Sinking Pond in October 1999 and operated through September 2002 (table 10), which provided data for three complete filling and draining cycles to verify the hydrologic model of Sinking Pond. As noted during calibration, the greatest departures of calculated from observed stage values occurred during the early filling stages, with modeled stage consistently lagging behind the observed rises (fig. 20). For the purpose of assessing broad temporal trends driven by climate, the verification indicates that the model produces an adequate simulation of the basin and pond's hydrologic response.
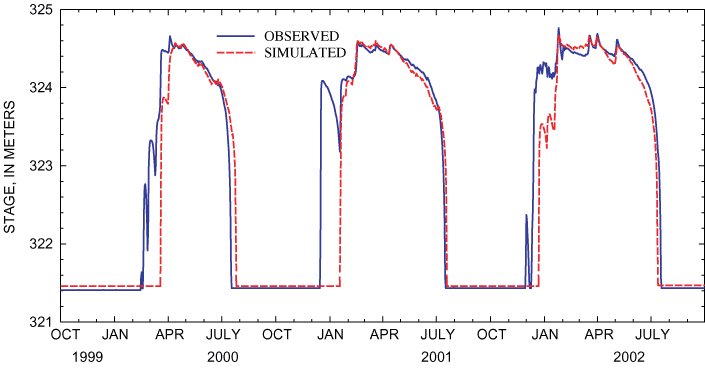
The model equations were applied to daily precipitation and temperature data covering the period beginning January 1, 1854, and ending September 30, 2002. As the initial condition, storage in Sinking Pond (SP) and the basin vadose zone (SB) were set at their respective overflow thresholds, PO and BO (table 9). Figure 21 shows the 148-year simulated hydrologic record for Sinking Pond. In general, the record shows increasing frequency and duration of ponding, beginning around 1945, which after 1970 reached levels unprecedented in the simulated record.
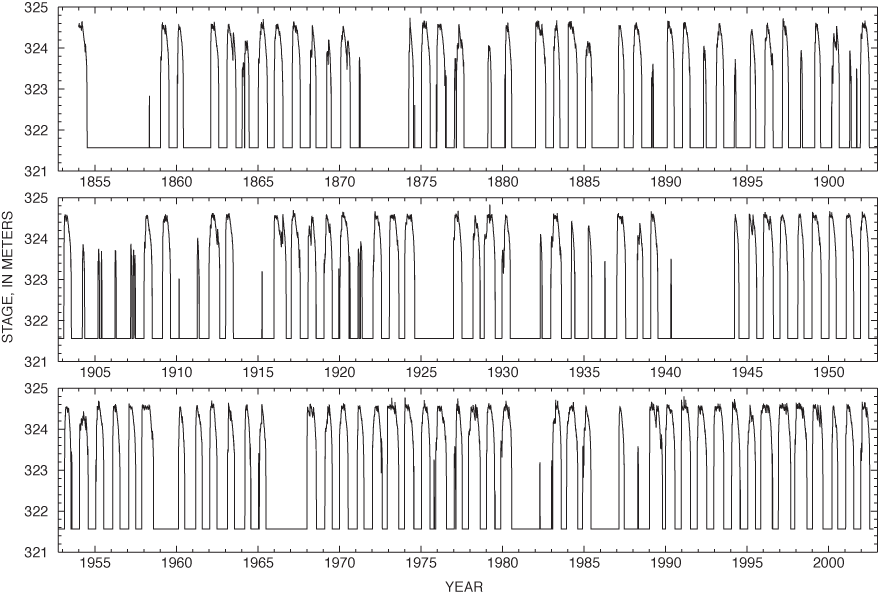
Figure 21. Simulated stage for Sinking Pond, January 1854 through September 2002.
One measure of ponding frequency is how often the water-surface elevation exceeds that of the spillway (324.53 m above NGVD 29). From 1855 through 1943, the simulated record shows flooding above the spillway occurring in 46 of 89 years, indicating that, on average, the pond had a 52 percent probability of overflowing in any given year. From 1944 through 2002, that probability increased to 86 percent, with flooding above the spillway occurring in 51 of 59 years (fig. 22).
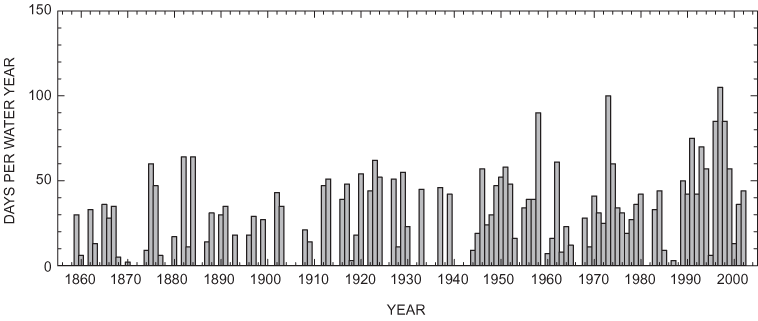
Simulated duration of ponding at or above the spillway elevation also increased after 1943. From 1855 through 1943, inundation of the spillway equaled or exceeded 50 days per water year in 9 years; the longest annual spillway inundation during this period was 64 days in 1882 and in 1884 (fig. 22). From 1944 through 2002, the spillway was flooded for 50 or more days per water year 15 times. Annual inundation of the spillway equaled or exceeded 70 days in 7 years, 5 of which had durations greater than 80 days (fig. 22).
The simulated hydrologic record shows increasing ponding duration for all three ponding-depth classes discussed in previous sections. Complete inundation of shallow areas (324 to 324.5 m above NGVD 29) corresponds to stages of 324.5 m or higher. Prior to 1944, the centered 5-year average annual inundation of shallow areas reached or exceeded 50 days per water year during only one 5-year period, 1921-25 (fig. 23). After 1944, the 5-year average annual inundation of shallow areas reached or exceeded 50 days during three distinct periods: water years 1947 through 1960, 1971 through 1978, and 1989 through 2000. After 1944, the simulated record shows a steady increase in the peak average annual inundation of shallow areas: 72 days per water year for the 5 years centered on 1950; 84 days for the period centered on 1974, and 95 days for the period centered on 1998 (fig. 23).
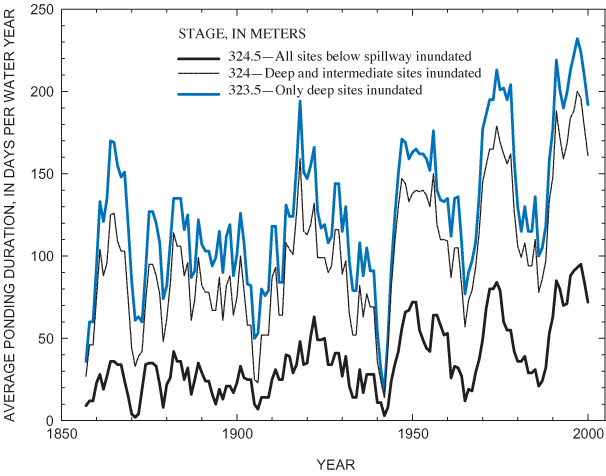
The simulation shows an increase in ponding duration of intermediate (323.5 to 323.99 m above NGVD 29) and deep (below 323.5 m above NGVD 29) areas beginning around 1970. Before 1970, the simulated hydrologic record shows average annual inundation of intermediate areas exceeding 150 days per water year for only one 5-year period, centered on 1918. For 1970 through 2000, the simulated 5-year average annual ponding duration of intermediate areas exceeds 150 days for 18 of 31 5-year periods (fig. 23). The average annual inundation of deep areas approaches 200 days only once before 1970, reaching 194 days for the 5-year period centered on 1918. During the periods 1971 through 1978 and 1991 through 2000, the simulated 5-year average annual inundation of deep areas remains above 185 days per water year, exceeding 200 days for 11 of 18 years (fig. 23).
Deep areas (below 323.5 m above NGVD 29) compose only about 20 percent of Sinking Pond's maximum flooded area. However, these areas encompass roughly two thirds of the pond's overall relief and contain the core of the overcup oak habitat (Wolfe, 1996a, b). More than half of the overcup oaks surveyed previously by Wolfe (1996a, b) and more than 60 percent of those surveyed in this study were found below 323.5 m above NGVD 29 (fig. 15). In both studies, more than 85 percent of surveyed overcup oaks were found at intermediate or deep ponding depths. The results of hydrologic modeling indicate that ponding durations in these areas have increased substantially since 1970, with important implications for the ecology of Sinking Pond.
Comparison of the density of overcup oak seedlings with that of surviving adults indicates that under pre-1970 hydrologic conditions, seedlings in Sinking Pond were subject to rigorous attrition. In 2001, overcup oak seedlings were present in 70 percent of the 4-m2 subplots in the surveyed area (fig. 7), with an average density of 10 seedlings per subplot. If, on average, 5 seedlings germinate on each subplot during each of the past 148 years, the period of hydrologic simulation would have seen more than 4 million seedlings germinated in the surveyed area, of which only about 400 seedlings survived through 2001, giving an attrition rate of 99.99 percent. Competition for light, herbivores, pathogens, fire, wind, lightning, ice, and all the other mortality factors operating in an oak forest contributed to this attrition, but the harsh hydrologic environment (Solomon, 1990) also constrained recruitment and survival. Despite such constraints, recuitment and survival were sufficient to maintain an apparently stable, closed-canopy stand of overcup oak in Sinking Pond for at least 150 years.
Hydrologic change in Sinking Pond since 1970 appears to have pushed the overcup oaks in deep and intermediate areas across a critical ecological threshold. The most striking changes in Sinking Pond's hydrologic regime is the pronounced increase in ponding duration, particularly the increased frequency of annual inundation exceeding 200 days per year in deep and even intermediate areas (fig. 23). The ecological threshold that has been crossed is the change from nearly total to total attrition of seedlings before they can recruit into the sapling stage.
Comparing the ring-count dates and elevations of sampled overcup oaks with the historical changes in the elevations subjected to prolonged ponding shows a strong relation between the overcup oak recruitment and ponding duration (fig. 24). Before 1970, model results indicate that inundation of any part of the pond for 200 days or more was relatively uncommon. Eighty-five percent of the sampled overcup oaks and 100 percent of the sampled trees in deep and intermediate areas (below 324 m) germinated during this period. After 1970, the frequency of simulated 200-day filling events increases sharply, with corresponding decreases in the temporal density and range of elevation of sampled trees. Hydrologic simulation indicates inundation of 200 days or more in 18 of 33 years from 1970 through 2002, compared with only 20 out of the previous 115 years (fig. 24).
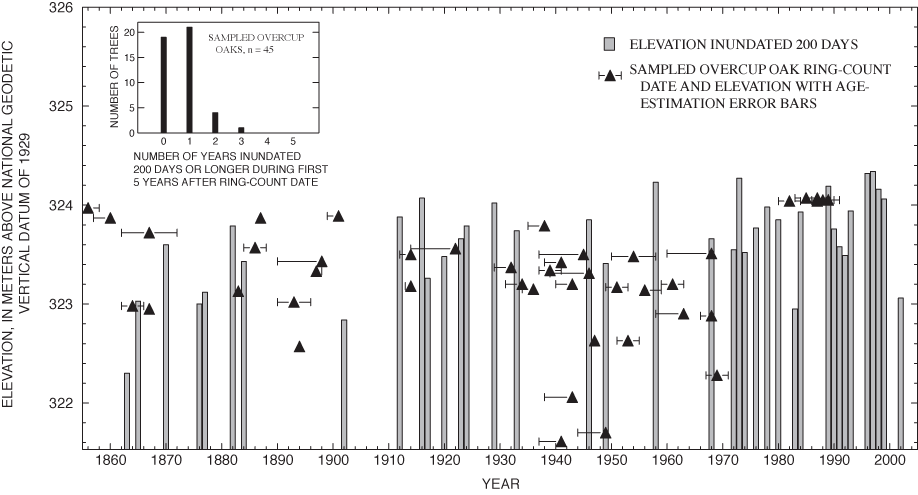
Before and after 1970, tree regeneration and survival appear to be most successful at times and elevations free from recurrent, prolonged ponding over several successive years. Survival of trees dated to the dry period from 1885 through 1900 (six trees from 15 years or 0.28 tree per year) is about twice that from a more recent wet period from 1910 through 1933 (three trees from 24 years or 0.125 tree per year). The densest concentration of sampled trees, including those at the lowest and third-lowest elevations in the sample, was dated to the dry period 1934-45 (fig. 24). The seven post-1970 trees in the sample have elevations tightly clustered near 324 m. Five of these seven trees have ring-count dates occurring in the drought years 1985-88, in which the simulated hydrologic record shows no 200-day ponding durations in the pond (fig. 24).
The strong relation between the 200-day ponding elevation and the dates and elevations of sampled trees supports the previous conclusion that prolonged inundation during the first few years after germination is a critical constraint on seedling survival and recruitment. The inset in figure 24 shows the distribution of 200-day inundation exceeding the elevations of sampled overcup oak during the first 5 years after their ring-count dates. Forty of 45 trees (89 percent) have ring-count dates and elevations that show 200 days of inundation in one or none of their first 5 years. Just one tree (2 percent) has an elevation and ring-count date that indicates 3 of the first 5 years with 200 days or longer of inundation, and no trees had more than 3 such years (fig. 24).
The simulated inundation histories of the sampled trees provide a more detailed picture of the relation between post-germination ponding and the recruitment and survival of overcup oaks. One measure of a tree's post-germination inundation history is the recruitment inundation, defined for this report as the median annual inundation, in days inundated per water year, for the 5-year period following germination. Recruitment inundation can be calculated for any tree for which elevation and germination date are known or estimated and for any site or group of sites in any set of 5-year recruitment periods. Recruitment inundations were determined for the sampled trees, based on each tree's elevation and ring-count date, ranked in ascending order, and plotted as a cumulative frequency distribution (fig. 25). A smooth curve through the points on the cumulative frequency plot (fig. 25A) provides an estimate of the cumulative distribution function (cdf) of recruitment inundation for the population from which the sample was drawn (Iman and Conover, 1983), in this case the surviving adult overcup oaks in the 2.3-ha sampled area.
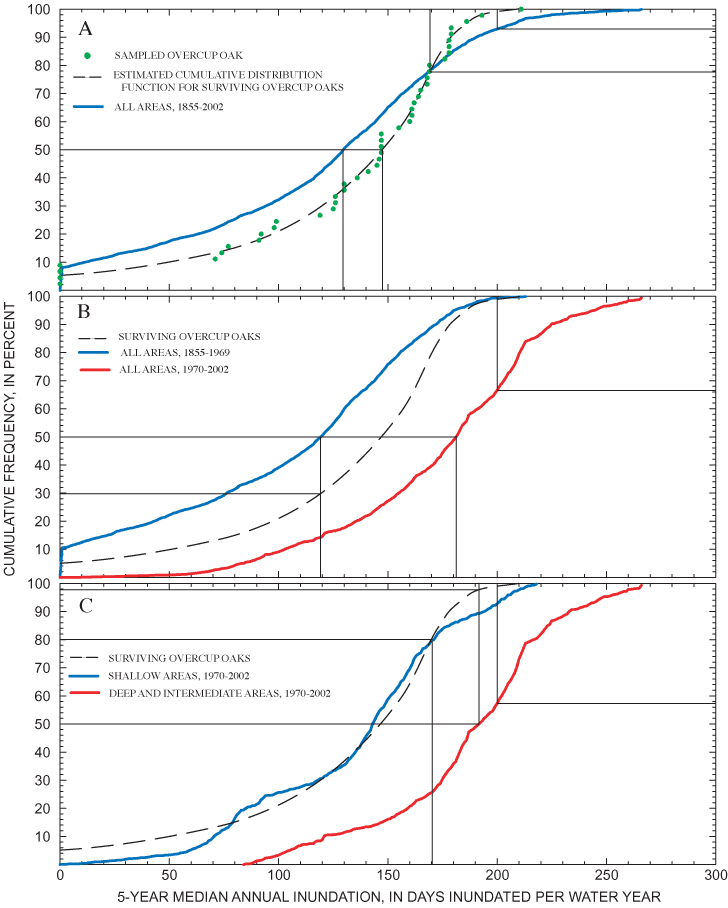
Similarly, recruitment inundation was calculated for the potential germination sites in the sampled area across the 148-year period of hydrologic simulation. The simulated hydrologic record contains 144 periods of 5 consecutive years, each of which represents a possible recruitment window, and each 4-m2 subplot in the sampled area is a potential germination site with known elevation. Multiplying 5,700 germination sites by all 5-year periods yields 820,800 possible recruitment opportunities in the sampled area during the period of hydrologic simulation.
The 820,800 calculated values of recruitment inundation were ranked in ascending order and plotted as a frequency distribution, which was then compared to the estimated cdf for surviving overcup oaks in the sampled area (fig. 25A). For the first 78 percent of the two distributions, the estimated cdf for surviving overcup oaks lies to the right (wetter) side of the distribution of all possible recruitment opportunities. For example, the 50th percentile for all recruitment opportunities is 130 days inundated per year, compared with 148 days for surviving overcup oaks (fig. 25A). The two distributions cross at the 78th percentile (about 170 days), and the upper limit for all recruitment opportunities exceeds that of the estimated cdf for overcup oaks by 55 days. Seven percent of all recruitment opportunities had recruitment inundations exceeding 200 days, approaching the upper limit (100th percentile) of the estimated overcup oak cdf (fig. 25A).
The effect of hydrologic change after 1970 on the frequency distribution of recruitment inundation in the 2.3-ha sampled area is shown in figure 25B. The distribution of recruitment inundation for pre-1970 recruitment opportunities lies to the left (drier) side of the estimated cdf for surviving overcup oaks, suggesting that in the past, prolonged inundation in the first few years was not a critical constraint on seedling survival and recruitment. The rightward (wetter) shift of the distribution of recruitment inundation after 1970 represents the increase in the ponding duration experienced by seedlings in their first few years. The median recruitment inundation for pre-1970 recruitment opportunities is 120 days, compared with 181 days for post-1970 recruitment opportunities and 148 days for the estimated cdf for surviving overcup oaks (fig. 25B). Nearly 32 percent of post-1970 recruitment opportunities have recruitment inundation exceeding 200 days (fig. 25B).
Under current (post-1970) hydrologic conditions, the concentration of successful overcup oak recruitment has shifted from deep and intermediate to shallow sites in Sinking Pond. The relation of recruitment inundation to this spatial shift is shown in figure 25C. In general, the recruitment-inundation distribution for shallow post-1970 recruitment opportunities lies close to the estimated cdf for surviving overcup oaks, indicating that post-1970 recruitment inundation of shallow sites approximates the recruitment-inundation regime that in the past permitted overcup oaks to recruit and survive. In contrast, the post-1970 recruitment inundation for deep and intermediate sites is much farther to the right (wetter) side of the cdf for surviving overcup oaks. The median post-1970 recruitment inundation for deep and intermediate sites is 192 days, which equals the 98th percentile of the estimated cdf for surviving overcup oaks; 42 percent of post-1970 deep and intermediate recruitment opportunities had recruitment inundation exceeding 200 days (fig. 25C).
As noted in this report and by McCarthy and Evans (2000), successful recruitment of overcup oak saplings and small adults currently is confined to areas with ponding depths less than 0.5 m, corresponding to elevations between 324 m and 324.5 m. The analysis of recruitment inundation confirms that current ponding regimes in these shallow areas are consistent with those that formerly permitted overcup oak recruitment in deeper parts of Sinking Pond—typically corresponding to recruitment inundation of 100 to 180 days (fig. 25). The apparent preference of surviving overcup oaks for this range of recruitment inundation does not reflect a physiological requirement for poorly drained soil or periodic inundation (Solomon, 1990). Rather, the overcup oaks in Sinking Pond are exploiting sites that may be barely tolerable for them but are even less tolerable for their competitors.
The spatial shift of suitable recruitment inundation from deep and intermediate to shallow sites represents a fundamental change in the ecological structure of Sinking Pond. In the past, overcup oak and associated tree species maintained a closed-canopy forest across most of the inundated area of Sinking Pond. Current patterns of recruitment and mortality indicate a transition towards a progressively more open canopy. Tree-ring analysis suggests that the former structure, a closed-canopy forest with the deepest areas dominated by overcup oak, has been relatively stable for at least 150 years. Neither the structural characteristics of the emerging ecosystem nor the climatic conditions under which it is developing seem likely to produce similar stability in the future.
Increased precipitation and streamflow in the eastern United States since 1970 is consistent with the results of global circulation models (GCMs) that simulate global climate response to greenhouse gas emissions (Karl and others, 1996; Burkett and others, 2001; McCabe and Wolock, 2002). Applying the GCM developed by the Hadley Climate Centre (United Kingdom) to the southeastern United States, Burkett and others (2001) projected regional mean annual temperature increasing by about 2.3 °C and precipitation increasing by about 20 percent between 2000 and 2100. The implications of this projection for Sinking Pond would be persistent prolonged (greater than 200 days per year) annual ponding into the foreseeable future.
Persistence of the wetting trend of the past several decades would sustain the spatial patterns of tree mortality and survival documented in this report. The former core of the overcup oak stand in Sinking Pond—deep and intermediate areas below 324 m above NGVD 29—would become progressively more open as adult trees die without being replaced. Shallow areas below the spillway elevation might sustain overcup oak as recruitment of competing species suffered under the new ponding regime. However, the resulting stand would have spatial and ecological characteristics markedly different from those of the historic stand.
The historic overcup oak stand, centered in the deep areas of Sinking Pond (about 20 percent of the maximum ponded area), was spatially compact and surrounded by buffer zones of intermediate to shallow ponding depth. When a canopy gap developed in the core area, overcup oak seedlings, saplings, or small adults were available to fill the gap. Gaps in the intermediate and shallow buffer zones provided additional opportunities for overcup oak release, especially during relatively wet years, when recruitment by competing species was suppressed. The spatial arrangement of a compact, monospecific core, surrounded by ecological buffer zones, favored structural stability and longevity for the overcup oak stand in Sinking Pond across the range of climatic fluctuations and other environmental stresses that occurred before 1970.
In contrast, the spatial characteristics of the emerging overcup oak stand are likely to exacerbate the stand's vulnerability to competition and environmental stress. Under current conditions, overcup oak regeneration in Sinking Pond is confined to areas with elevations between 324 and 324.5 m above NGVD 29, which compose about 30 percent of the maximum ponded area of Sinking Pond. These shallow areas currently have ponding regimes similar to those that characterized the deep core of the overcup oak stand before 1970. However, the shallow areas below the spillway elevation do not form a single compact core; rather these areas extend in a more or less narrow band around the pond's perimeter. Continued shifting of the overcup oak stand from the pond's interior to its perimeter would result in a progressively more dispersed, less concentrated seed source.
The emerging overcup oak stand would likely form a discontinuous linear swath between areas too wet for overcup oak recruitment and drier sites where overcup oak is at a competitive disadvantage. Even if shallow ponded areas were too wet for establishment of sweetgum and other competing species, upland trees near the pond margin could still compete with nearby overcup oaks for light. Because the spillway limits the elevation of prolonged inundation, further increases in ponding duration could make even shallow ponded areas untenable for overcup oak recruitment without creating new opportunities at higher elevations.
The broad structural change in the Sinking Pond ecosystem discussed above would likely be accompanied by many changes in ecological function, including changes in the interactions of fauna with the pond. Several hundred pairs of great blue herons currently nest in the deep and intermediate areas of Sinking Pond. Continued attrition without replacement of adult trees in these areas would steadily reduce the number of available nesting sites. Concurrently, loss of acorn production would reduce the value of Sinking Pond as a foraging site for deer, wood ducks, and other wildlife. On the other hand, upstream movement of fish into Sinking Pond from Crumpton Creek would likely increase with increased duration and magnitude of surface outflow. In response to increased fish numbers and residence time, snapping turtles and other aquatic predators might visit Sinking Pond in greater numbers and spend more time there.
Changes in hydrology and biota, such as those described above, would affect carbon and nutrient cycles in Sinking Pond. Breakdown of organic matter might slow down or proceed through different chemical pathways in response to longer inundation.
This report presents several lines of mutually supporting botanical and hydrologic evidence leading to the following conclusions:
(1) Areas of Sinking Pond with normal high-water ponding depths greater than 0.5 m have become untenable for overcup oak regeneration, even though these areas are dominated by adult overcup oaks; (2) Failure of tree regeneration in Sinking Pond occurs at the critical stage of recruitment from seedling to sapling size class; (3) Failure of tree regeneration in Sinking Pond began abruptly around 1970 and has persisted since then; (4) Prolonged (greater than 200 days per year) inundation during the first 5 years after germination is the proximate cause of tree-regeneration failure in Sinking Pond; (5) Increased ponding duration within Sinking Pond since 1970 has been driven by a step increase in precipitation, which occurred around that time; and (6) Increased rainfall since 1970 is not a local phenomenon but has been documented across much of the eastern United States.These conclusions have several implications for wetland management in and around the study area. One implication is that local management options for enhancing regeneration and survival of overcup oak in Sinking Pond are limited. Little or nothing can be done to reduce ponding duration in deep or intermediate areas. Even massive excavation of the spillway and outflow channel would leave deep areas of the pond unaffected (Appendix B). Selective thinning of red maple, sweetgum, and other trees may enhance light availability for overcup oak saplings and small adults in shallow areas of the pond. Such thinning would help maintain a remnant population of overcup oak around the pond margins but would not reverse broad changes in ecological structure.
A second implication of climate driven hydrologic change in Sinking Pond is that nearby wetlands have experienced similar increases in precipitation. The likely ecological response of nearby wetlands to increased precipitation would depend on both hydrologic and ecological factors. At present, little is known about ecological change in nearby wetlands, but previous studies (Wolfe, 1996a, b) and the results of this study suggest some hypotheses.
Wetlands whose hydrologic regimes are most responsive to seasonal ground-water conditions and whose biota are most sensitive to increased ponding duration would most likely experience the most pronounced ecological response to increased precipitation. For example, Willow Oak Swamp and Westall Swamp (fig. 2) are geomorphically and hydrologically similar to Sinking Pond in having maximum depths greater than 2 m, relatively efficient internal drains, and bottom elevation well within the normal range of seasonal water-table fluctuation (Wolfe, 1996a, b). Both of these depressions support forests dominated by species, such as willow oak and sweetgum, which are less tolerant of inundation than the overcup oaks of Sinking Pond. The combination of hydrologic similarity to Sinking Pond and biota with comparatively low tolerance for increased ponding duration would tend to make these wetland ecosystems vulnerable to the kind of climate-driven ecological change that has been documented in Sinking Pond.
In contrast, Tupelo Swamp is dominated by water tupelo, a species more tolerant of inundation than overcup oak, and has geomorphic and hydrologic characteristics that differ from those of Sinking Pond. Shallow (less than 1.5 m) maximum depth, lack of an efficient internal drain, and a bottom elevation higher than all but the highest recorded local water-table elevations make the hydrologic regime of Tupelo Swamp less dependent on ground-water conditions than that of Sinking Pond (Wolfe, 1996a). This hydrologic difference and the relatively high tolerance of its biota to inundation may tend to limit the ecological effect of increased precipitation in Tupelo Swamp and similar depression wetlands.
Hydrologic monitoring of Sinking Pond and other depression wetlands would be useful in testing the hypotheses summarized above. Such monitoring also could support refining the water-budget model described in this report and developing similar models for other depression wetlands, which could be used to project hydrologic conditions under different climatic scenarios. Ecological monitoring of Sinking Pond, for example permanently marking the 2.3-ha area used for spatial analysis and resurveying its vegetation at regular intervals, would improve understanding of the mechanics and trajectory of ecological change and may detect management opportunities that might otherwise be missed. Similarly, ecological monitoring of nearby depression wetlands would improve understanding of the extent, rates, and mechanism of wetland ecological response to changing climate at AAFB.
Beyond local management concerns, this study shows that current hydrologic conditions cannot be assumed to represent the ecological or historical norm without risk of substantial error. A 30-year hydrologic record is rare for most wetland settings, but even such a relatively long record would give a distorted view of the hydrologic conditions under which the overcup oak stand in Sinking Pond developed.
Carter and others (1979) listed five urgent research needs in wetland hydrology: (1) better hydrologic measurement techniques; (2) measurement of hydrologic inputs and outputs of representative wetlands with appropriate error analysis; (3) improved understanding of wetland soil-water-vegetation relations; (4) detailed long-term studies of representative wetlands in different environmental settings; and (5) predictive and analytical models of wetlands based on hydrologic data. Arguing that these research needs have been fully addressed during the 25 years since Carter and others pointed them out would be difficult. Nevertheless, the results of this study indicate the need for a sixth item on the list: comparison of current to past wetland hydrologic regimes and analysis of wetland response to climate change.
Many individuals representing numerous public agencies and private organizations contributed to this investigation and report. We owe particular thanks to Geoffrey Call, Mark Moran, Kevin Fitch, and Edward Roworth of Arnold Engineering Development Center (AEDC) for logistical support, background data, and intellectual exchange through the course of the investigation. Members of the AEDC Integrated Planning Core Team assisted with numerous questions, discussions, and criticisms at different stages of the study. We also thank Mary Priestley and Yolande Gottfried of the Sewanee Herbarium for compiling the plant species list for Sinking Pond (Appendix A) and for their assistance with data collection in the field.
Barclay, F.H., 1957, Natural vegetation of Johnson County, Tennessee: Ph.D. dissertation, University of Tennessee, 147 p.
Baskin, J.M., Baskin, C.C., and Chester, E.W., 1994, The Big Barrens region of Kentucky and Tennessee—further observations and considerations: Castanea, v. 59, p. 226-254.
Baskin, J.M., Baskin, C.C., and Chester, E.W., 1999, The Big Barrens region of Kentucky and Tennessee, chap. 11 in Anderson, R.C., Fralish, J.S., and Baskin, J.M. eds., Savannas, barrens, and rock outcrop plant communities of North America: Cambridge, U.K., Cambridge University Press, p. 190-205.
Bowen, Brian, and Pyne, Milo, 1995, May Prairie: Tennessee Conservationist, v. 61, no. 6, p. 19-23.
Brahana, J.V., and Bradley, M.W., 1986a, Preliminary delineation and description of the regional aquifers of Tennessee—The Highland Rim aquifer system: U.S. Geological Survey Water-Resources Investigations Report 82-4054, 38 p.
Brahana, J.V., and Bradley, M.W., 1986b, Preliminary delineation and description of the regional aquifers of Tennessee—The Central Basin aquifer system: U.S. Geological Survey Water-Resources Investigations Report 82-4002, 35 p.
Broadfoot, W.M., and Williston, W.H., 1973, Flooding effects of southern forests: Journal of Forestry, v. 71, no. 9, p. 584-587.
Burchett, C.R., 1977, Water resources of the upper Duck River basin, central Tennessee: Tennessee Division of Water Resources, Water Resources Series no. 12, 103 p.
Burchett, C.R., and Hollyday, E.F., 1974, Tennessee's newest aquifer [abs.]: Geological Society of America Abstracts with Programs, v. 6, no. 4, p. 338.
Burkett, Virginia, Ritschard, Ronald, McNulty, Steven, O'Brien, J.J., Abt, Robert, Jones, James, Hatch, Upton, Murray, Brian, Jagtap, Shrikant, and Cruise, Jim, 2001, Potential consequences of climate variability and change for the southeastern United States, chap. 5 of National Assessment Synthesis Team, U.S. Global Change Research Program, Climate change impacts on the United States: the potential consequences of climate variability and change—foundation: Cambridge, UK, Cambridge University Press, p. 137-164.
Call, Geoff, 1999, Integrated ecosystem management plan: U.S. Air Force, Arnold Air Force Base, Tenn., Arnold Engineering Development Center, 288 p.
Carter, Virginia, Bedinger, M.S., Novitsky, R.P., and Wilen, W.O., 1979, Water resources and wetlands, in Greeson, P.E., Clark, J.R., and J.E., eds., Wetland functions and values—the state of our understanding: American Water Resources Association, p. 344-376.
Chester, E.W., Noel, S.M., Baskin, J.M., Baskin, C.C., and McReynolds, M.L., 1995, A phytosociological analysis of an old-growth upland wet woods on the Pennyroyal Plain, southcentral Kentucky, USA: Natural Areas Journal, v. 15, no. 4, p. 297-307.
Chester, E.W., Wofford, B.E., Baskin, J.M., and Baskin, C.C., 1997, A floristic study of barrens on the southwestern Pennyroyal Plain, Kentucky and Tennessee: Castanea, v. 62, p. 161-172.
Conner, C.T., 2000, Soil survey of Jefferson County: U.S. Department of Agriculture, Natural Resources Conservation Service, 178 p.
Cowardin, L.M., Carter, Virginia, Golet, F.C., and LaRoe, E.T., 1979, Classification of wetlands and deepwater habitats in the United States: U.S. Fish and Wildlife Service, Biological Services Program, FWS/OBS 79/31, 131 p.
DeSelm, H.R., 1981, Characterization of some southeastern barrens with special reference to Tennessee, in Stuckey, R.L., and Reese, K.J., eds., The prairie peninsula—in the shadow of Transeau—Proceedings of the Sixth North American Prairie Conference, Aug. 12-17, 1978: Columbus, Ohio, The Ohio State University, Ohio Biological Survey, Biological Note no. 15, p. 86-88.
DeSelm, H.R., 1986, Natural forest openings on uplands of the Eastern United States, in Kulhavy, D.L., and Conner, R.N., eds., Wilderness and natural areas in the Eastern United States—a management challenge: Nacogdoches, Texas, Stephen F. Austin State University, School of Forestry, Center for Applied Studies, p. 366-375.
DeSelm, H.R., 1989, The barrens of Tennessee: Journal of the Tennessee Academy of Science, v. 64, p. 89-95.
DeSelm, H.R., Clebsch, E.E.C., Nichols, G.M., and Thor, E., 1974, Response of herbs, shrubs and tree sprouts in prescribed-burn hardwoods in Tennessee, in Komarek, E.V., Sr., ed., Proceedings, 13th Annual Tall Timbers Fire Ecology Conference: Tallahassee, Florida, Tall Timbers Research Station, p. 331-344.
Douglas, E.M., Vogel, R.M., and Kroll, C.N., 2000, Trends in floods and low flows in the United States—impact of spatial correlation: Journal of Hydrology, v. 240, p. 90-105.
Easterling, D.R., Evans, J.L., Groisman, P.Y., Karl, T.R., Kunkel, K.E., and Ambenje, P., 2000, Observed variability and trends in extreme climate events—a brief review: Bulletin of the American Meteorological Society, v. 81, no. 3, p. 417-426.
Eisenlohr, W.S., Jr., and others, 1972, Hydrologic investigations of prairie potholes in North Dakota, 1959-68: U.S. Geological Survey Professional Paper 585-A, 102 p.
Ellis, W.H., and Chester, E.W., 1989, Upland Swamps of the Highland Rim of Tennessee: Journal of the Tennessee Academy of Science, v. 64, no. 3, p. 97-101.
Ewel, K.C., and Odum, H.T., eds., 1984, Cypress swamps: Gainesville, University Presses of Florida, 472 p.
Fenneman, N.M., 1938, Physiography of Eastern United States: New York, McGraw-Hill, 714 p.
Gattinger, Augustin, 1901, The flora of Tennessee and a philosophy of botany—respectfully dedicated to the citizens of Tennessee: Nashville, Gospel Advocate Publishing Co., 296 p.
Gill, C.J., 1970, The flooding tolerance of woody species—a review: Forest Abstracts, v. 31, p. 671-688.
Greear, P.F., 1967, Composition, diversity, and structure of the vegetation of some natural ponds in northwest Georgia: Ann Arbor, Michigan, University microfilms (Ph.D. dissertation, University of Georgia), 183 p.
Groisman, P.Y., and Easterling, D.R., 1994, Variability and trends of total precipitation and snowfall over the United States and Canada: Journal of Climatology, v. 7, no. 1, p. 184-205.
Groisman, P.Y., Knight, R.W., and Karl, T.R., 2001, Heavy precipitation and high streamflow in the contiguous United States—trends in the 20th century: Bulletin of the American Meteorological Society, v. 82, no. 2, p. 219-246.
Hall, R.B.W., and Harcombe, P.A., 1998, Flooding alters apparent position of floodplain saplings on a light gradient: Ecology, v. 79, no. 3, p. 847-855.
Harms, W.R., Schreuder, H.T., Hook, D.D., Brown, C.L., and Shropshire, F.W., 1980, The effects of flooding on the swamp forest in Lake Ocklawaha, Florida: Ecology, v. 61, no. 6, p. 1412-1421.
Haugh, C.J., and Mahoney, E.N., 1994, Hydrogeology and simulation of ground-water flow at Arnold Air Force Base, Coffee and Franklin Counties, Tennessee: U.S. Geological Survey Water-Resources Investigations Report 93-4207, 69 p.
Haugh, C.J., Mahoney, E.N., and Robinson, J.A., 1992, Well-construction, water-level, geophysical, and water-quality data for ground-water monitoring wells for Arnold Air Force Base, Tennessee: U.S. Geological Survey Open-File Report 92-135, 88 p.
Helsel, D.R., and Hirsch, R.M., 1992, Statistical methods in water resources: New York, Elsevier, 522 p.
Helvey, J.D., 1974, A summary of rainfall interception by certain conifers of North America, in Biological Effects on the Hydrological Cycle—International Seminar for Hydrology Professors, 3rd, Purdue University, July 1971, Proceedings: West Lafayette, Indiana, Purdue University, p. 103-113.
Hemond, H.F., 1980, Biogeochemistry of Thoreau's Bog, Concord, Massachusetts: Ecological Monographs, v. 50, no. 4, p. 507-526.
Hendricks, E.L., and Goodwin, M.H., Jr., 1952, Water-level fluctuations in limestone sinks in southwestern Georgia, in Contributions to the hydrology of the United States, 1948-52: U.S. Geological Survey Water-Supply Paper 1110-E, p. 157-246, 1 pl., appendix.
Hewlett, J.D., and Hibbert, A.R., 1961, Increases in water yield after several types of forest clearing: Hydrological Sciences Journal, v. 6, p. 5-17.
Heyward, Frank, and Barnette, R.M., 1934, Effect of frequent fires on chemical composition of forest soils in the longleaf pine region: Florida Agricultural Experiment Station Bulletin 265, 39 p.
Hibbert, A.R., 1967, Forest treatment effects on water yield, in Sopper, W.E., and Lull, H.W., eds., Proceedings—International Symposium on Forest Hydrology: Oxford, Pergamon, p. 527-543.
Hook, David, 1984, Waterlogging tolerance of lowland tree species of the south: Southern Journal of Applied Forestry, v. 8, p. 136-149.
Hornbeck, J.W., Pierce, R.S., and Federer, C.A., 1970, Streamflow changes after forest clearing in New England: Water Resources Research, v. 6, p. 1124-1132.
Hosner, J.F., and Boyce, S.G., 1962, Tolerance to water saturated soil of various bottomland hardwoods: Forest Science, v. 8, no. 2, p. 180-186.
Huenneke, L.F., and Sharitz, R.R., 1986, Microsite abundance and distribution of woody seedlings in a South Carolina cypress-tupelo swamp: American Midland Naturalist, v. 115, no. 2, p. 328-335.
Iman, R.I., and Conover, W.J., 1983, A modern approach to statistics: New York, John Wiley and Sons, 497 p.
Jenkins, J.C., 2000, Soil survey of Cheatham County: U.S. Department of Agriculture, Natural Resources Conservation Service, 135 p.
Johnson, R.D., 1964, Water relations of pinus radiata under plantation conditions: Australian Journal of Botany, v. 12, p. 111-124.
Jones, R.H., Sharitz, R.R., Dixon, P.M., Segal, D.S., and Schneider, R.L., 1994, Woody plant regeneration in four floodplain forests: Ecological Monographs, v. 64, no. 3, p. 345-367.
Karl, T.R., and Knight, R.W., 1998, Secular trends of precipitation amount, frequency, and intensity in the USA: Bulletin of the American Meteorological Society, v. 79, no. 2, p. 231-241.
Karl, T.R., Knight, R.W., Easterling, D.R., and Quayle, R.G., 1996, Indices of climate change for the United States: Bulletin of the American Meteorological Society, v. 77, no. 2, p. 279-292.
Kartesz, J.T., and Meacham, C.A., 1999, Synthesis of the North American flora [computer program]. Version 1.0: Chapel Hill, North Carolina Botanical Garden.
Killebrew, J.B., and Safford, J.M., 1874, Introduction to the resources of Tennessee: Spartanburg, S.C., The Reprint Group, 1204 p.
Kral, Robert, 1973, Some notes on the flora of the southern states, particularly Alabama and Middle Tennessee: Rhodora, v. 75, p. 366-410.
Leitman, H.M., Sohm, J.E., and Franklin, M.A., 1983, Wetland hydrology and tree distribution of the Apalachicola River flood plain, Florida: U.S. Geological Survey Water-Supply Paper 2196-A, 52 p.
Lettenmaier, D.P., Wood, E.F., and Wallis, J.R., 1994, Hydro-climatological trends in the continental United States, 1948-88: Journal of Climatology, v. 7, no. 4, p. 586-607.
Lins, H.F., and Slack, J.R., 1999, Streamflow trends in the United States: Geophysical Research Letters, v. 26, no. 2, p. 227-230.
Lollar, E.H., 1924, Geologic structure of the "Barrens" of the Hollow Springs Quadrangle in Cannon and Coffee Counties, Tennessee: Nashville, Tenn., Vanderbilt University, M.S. Thesis, 69 p.
Love, T.R., Williams, L.D., Proffitt, W.H., Epley, I.B., and Elder, John, 1959, Soil survey of Coffee County, Tennessee: U.S. Department of Agriculture, Soil Conservation Service, Series 1956, no. 5, 112 p., 37 pls.
McCabe, G.J., and Wolock, D.M., 2002, A step increase in streamflow in the conterminous United States: Geophysical Research Letters, v. 29, no. 24, p. 2185-2189.
McCarthy, Sarah, and Evans, J.P., 2000, Population dynamics of overcup oak (Quercus lyrata) in a seasonally flooded karst depression: Journal of the Torrey Botanical Society, v. 127, no. 1, p. 9-18.
McCracken, Michael, Barron, Eric, Easterling, David, Felzer, Benjamin, and Karl, Thomas, 2001, Scenarios for climate variability and change, chap. 1 of National Assessment Synthesis Team, U.S. Global Change Research Program, Climate change impacts on the United States: the potential consequences of climate variability and change—foundation: Cambridge, UK, Cambridge University Press, p. 13-71.
Miller, R.A., 1974, Geologic history of Tennessee: Tennessee Division of Geology Bulletin 74, 63 p.
Mitsch, W.J., and Gosselink, J.G., 1993, Wetlands (3rd ed.): New York, Van Nostrand Reinhold, 722 p.
National Oceanic and Atmospheric Administration, 1900-2003, Climatological data annual summary, Tennessee: Asheville, N.C., National Climate Data Center, published annually.
Patterson, W.B., 1989, Vegetation and soils of the Sinking Pond area, Coffee County, Tennessee: Knoxville, University of Tennessee, M.S. Thesis, 105 p.
Pereira, H.C., 1973, Land use and water resources in temperate and tropical climates: London, Cambridge University Press, 246 p.
Phipps, R.L., 1985, Collecting, preparing, cross-dating, and measuring tree increment cores: U.S. Geological Survey Water-Resources Investigations Report 85-4148, 48 p.
Pritchett, W.L., 1979, Properties and management of forest soils: New York, Wiley, 500 p.
Pyne, Milo, 2000, Biogeographic study of The Barrens of the southeastern Highland Rim of Tennessee: Durham, N.C., The Nature Conservancy, 170 p.
Reily, P.W., and Johnson, W.C., 1982, The effects of altered hydrologic regime on tree growth along the Missouri River in North Dakota: Canadian Journal of Botany, v. 60, p. 2410-2423.
Rutter, A.J., 1972, Water consumption by forests, in Kozlowski, T.T., ed., Water deficits and plant growth: New York, Academic Press, v. 3, p. 23-84.
Schupp, E.W., 1995, Seed-seedling conflicts, habitat choice, and patterns of plant recruitment: American Journal of Botany, v. 82, no. 3, p. 399-409.
Smalley, G.W., 1983, Classification and evaluation of forest sites on the eastern Highland Rim and Pennyroyal: New Orleans, U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station, General Technical Report SO-43, 123 p.
Soil Conservation Service, 1991, State soil geographic data base (STATSGO) data user's guide: U.S. Department of Agriculture, Soil Conservation Service Miscellaneous Publication no. 1492.
Solomon, J.D., 1990, Quercus lyrata Walt., in Burns, R.M., and Honkala, B.H., technical coordinators, Silvics of North America, v. 2, Hardwoods: U.S. Department of Agriculture, Forest Service, Handbook 654, p. 681-685.
Springer, M.E., and Elder, J.A., 1980, Soils of Tennessee: University of Tennessee Agricultural Experiment Station Bulletin 596, 66 p.
Stanhill, Gerald, 1970, Water flux in temperate forests—precipitation and evapotranspiration, in Reichle, D.E., ed., Analysis of temperate forest ecosystems: Ecological studies, analysis, and synthesis, v. 1, New York, Springer-Verlag, p. 255-256.
Streng, D.R., Glitzenstein, J.S., and Harcombe, P.A., 1989, Woody seedling dynamics in an east Texas floodplain forest: Ecological Monographs, v. 59, no. 2, p. 177-204.
Svenson, H.K., 1941, Notes on the Tennessee flora: Journal of the Tennessee Academy of Science, v. 16, p. 111-160.
Swank, W.T., and Douglas, J.E., 1974, Streamflow greatly reduced by converting deciduous hardwoods to pine: Science, v. 85, p. 857-859.
Swank, W.T., Goebel, N.B., and Helvey, J.D., 1972, Interception loss in loblolly pine stands of the South Carolina Piedmont: Journal of Soil and Water Conservation, v. 26, p. 160-163.
Taylor, J.R., Cardamone, M.A., and Mitsch, W.J., 1990, Bottomland hardwood forests—their functions and values, in Gosselink, J.G., Lee, L.C., and Muir, T.A., eds., Ecological processes and cumulative impacts—illustrated by bottomland hardwood wetland ecosystems: Chelsea, Michigan, Lewis Publishers, p. 13-86.
Tennessee Department of Environment and Conservation, 2002, Rare species of Coffee County, accessed May 5, 2002, at http://www.state.tn.us/environment/nh/species/coffee.htm
Trousdell, K.B., and Hoover, M.D., 1955, A change in ground-water level after clearcutting of loblolly pine in the coastal plain: Journal of Forestry, v. 53, p. 493-498.
Tryon, B.W., 1990, Bog turtles (Clemmys muhlenbergii) in the south—a question of survival: Bulletin of the Chicago Herpetological Society, v. 25, no. 4, p. 57-66.
Twilley, R.R., 1982, Litter dynamics and organic carbon exchange in black mangrove (Avicennia germinans) basin forests in a southwest Florida estuary: University of Florida, PhD Dissertation, 262 p.
U.S. Geological Survey, 1970, The national atlas of the United States of America: Washington, D.C., U.S. Department of the Interior, Geological Survey, 417 p.
Wada, N., and Ribbens, E., 1997, Japanese maple (Acer palmatum var. Matsumurae, Aceraceae) recruitment patterns: seeds, seedlings, and saplings in relation to conspecific adult neighbors: American Journal of Botany, v. 84, no. 9, p. 1294-1300.
White, W.B., 1988, Geomorphology and hydrology of karst terrains: New York, Oxford University Press, 464 p.
Wilde, S.A., 1958, Forest soils: New York, Ronald Press, 537 p.
Wilson, C.W., Jr., 1976, Geologic map and mineral resources summary of the Manchester quadrangle, Tennessee: Tennessee Division of Geology, MRS 86-NE, scale 1:24,000.
Wolfe, W.J., 1996a, Hydrology and tree-distribution patterns of karst wetlands at Arnold Engineering Development Center, Tennessee: U.S. Geological Survey Water-Resources Investigations Report 96-4277, 46 p.
Wolfe, W.J., 1996b, Karst wetlands of The Barrens—geomorphic control of wetland hydrology and plant distribution on the Highland Rim of Tennessee: Ann Arbor, Michigan, University microfilms (Ph.D. dissertation, Clark University), 147 p.
Wolfe, W.J., Haugh, C.J., Webbers, Ank, and Diehl, T.H., 1997, Preliminary conceptual models of the occurrence, fate, and transport of chlorinated solvents in karst regions of Tennessee: U.S. Geological Survey Water-Resources Investigations Report 97-4097, 80 p.
Appendix A. Flora of Sinking Pond.
The following table lists plants observed within the seasonally inundated perimeter of Sinking Pond. Scientific and common names are from Kartesz and Meacham (1999).
| Family | Genus | Species | Common Name | ||
|---|---|---|---|---|---|
| Ferns | |||||
| Aspleniaceae | Asplenium | Platyneuron | (L.) B.S.P. | Ebony spleenwort | |
| Dryopteridaceae | Athyrium | filix-femina | (L.) Roth | Subarctic lady fern | |
| Dryopteridaceae | Polystichum | acrostichoides | (Michx.) Schott | Christmas fern | |
| Ophioglossaceae | Botrychium | sp. | Sw. | Grape fern, Moonwort | |
| Osmundaceae | Osmunda | regalis var. spectabilis | (Willd.) Gray | Royal fern | |
| Polypodiaceae | Pleopeltis | polypodioides ssp. polypodioides | (L.) Andrews & Windham | Resurrection fern | |
| Thelypteridaceae | Phegopteris | hexagonoptera | (Michx.) Fée | Broad beech fern | |
| Thelypteridaceae | Thelypteris | noveboracensis | (L.) Nieuwl. | New York fern | |
| Monocots | |||||
| Cyperaceae | Carex | debilis var. debilis | Michx. | White-edge sedge | |
| Cyperaceae | Carex | folliculata | L. | Northern-long sedge | |
| Cyperaceae | Carex | gracilescens | Steud. | Slender loose-flower sedge | |
| Cyperaceae | Carex | intumescens | Rudge | Greater bladder sedge | |
| Cyperaceae | Carex | joorii | Bailey | Cypress-swamp sedge | |
| Cyperaceae | Carex | lupulina | Muhl. ex Willd. | Hop sedge | |
| Cyperaceae | Carex | tribuloides | Wahlenb. | Blunt broom sedge | |
| Cyperaceae | Carex | typhina | Michx. | Cat-tail sedge | |
| Cyperaceae | Cyperus | pseudovegetus | Steud. | Marsh flat sedge | |
| Cyperaceae | Rhynchospora | corniculata | (Lam.) Gray | Short-bristle horned beak sedge | |
| Cyperaceae | Scirpus | cyperinus | (L.) Kunth | Cottongrass bulrush | |
| Juncaceae | Juncus | brachyphyllus | Wieg. | Short-leaf rush | |
| Juncaceae | Juncus | bufonius | L. | Toad rush | |
| Juncaceae | Juncus | coriaceus | Mackenzie | Leathery rush | |
| Juncaceae | Juncus | effusus | L. | Lamp rush | |
| Liliaceae | Maianthemum | racemosum | (L.) Link | Feathery false Solomon's-seal | |
| Poaceae | Andropogon | virginicus | L. | Broom-sedge | |
| Poaceae | Brachyelytrum | erectum | (Schreb. ex Spreng.) Beauv. | Bearded shorthusk | |
| Poaceae | Chasmanthium | latifolium | (Michx.) Yates | Indian wood-oats | |
| Poaceae | Chasmanthium | laxum | (L.) Yates | Slender wood-oats | |
| Poaceae | Danthonia | spicata | (L.) Beauv. ex Roemer & J. A. Schultes | Poverty wild oat grass | |
| Poaceae | Echinochloa | crus-galli | (L.) Beauv. | Large barnyard grass | |
| Poaceae | Microstegium | vimineum | (Trin.) A. Camus | Nepalese browntop | |
| Poaceae | Panicum | rigidulum | Bosc ex Nees | Red-top panic grass | |
| Poaceae | Panicum | verrucosum | Muhl. | Warty panic grass | |
| Poaceae | Saccharum | baldwinii | Spreng. | Narrow plume grass | |
| Poaceae | Sphenopholis | obtusata | (Michx.) Scribn. | Prairie wedgescale | |
| Smilacaceae | Smilax | ecirrata | (Engelm. ex Kunth) S. Wats | Upright carrion flower | |
| Smilacaceae | Smilax | rotundifolia | L. | Horsebrier | |
| Dicots | |||||
| Aceraceae | Acer | rubrum | L. | Red Maple | |
| Anacardiaceae | Rhus | copallinum | L. | Winged sumac | |
| Anacardiaceae | Rhus | glabra | L. | Smooth sumac | |
| Anacardiaceae | Toxicodendron | radicans | (L.) Kuntze | Eastern poison-ivy | |
| Annonaceae | Asimina | triloba | (L.) Dunal | Common Pawpaw | |
| Apiaceae | Cryptotaenia | canadensis | (L.) DC. | Canadian honeworth | |
| Apiaceae | Ptilimnium | nuttallii | (DC.) Britt | Laceflower | |
| Apocynaceae | Trachelospermum | difforme | (Walt.) Gray | Climbing-dogbane | |
| Aquifoliaceae | Ilex | opaca | Ait. | American holly | |
| Asteraceae | Bidens | frondosa | L. | Devil's-pitchfork | |
| Asteraceae | Erechtites | hieraciifolia | (L.) Raf. ex DC. | American burnweed | |
| Asteraceae | Erigeron | strigosus | Muhl. ex Willd. | Prairie fleabane | |
| Asteraceae | Eupatorium | capillifolium | (Lam.) Small | Dog-fennel | |
| Asteraceae | Eupatorium | serotinum | Michx. | Late-flowering thoroughwort | |
| Asteraceae | Helenium | autumnale | L. | Fall sneezeweed | |
| Asteraceae | Pluchea | camphorata | (L.) DC. | Plowman's-wort | |
| Asteraceae | Solidago | ulmifolia | Muhl. ex Willd. | Elm-leaf goldenrod | |
| Asteraceae | Symphyotrichum | lateriflorum var. lateriflorum | (L.)A.&D.Love | Farewell-summer | |
| Asteraceae | Symphyotrichum | pilosum var. pilosum | (Willd.) Nesom | White oldfield American-aster | |
| Asteraceae | Xanthium | strumarium | L. | Rough cockleburr | |
| Berberidaceae | Podophyllum | peltatum | L. | May-apple | |
| Betulaceae | Betula | nigra | L. | River birch | |
| Betulaceae | Carpinus | caroliniana | Walt. | American hornbeam | |
| Betulaceae | Corylus | cornuta | Marsh. | Beaked hazelnut | |
| Bignoniaceae | Campsis | radicans | (L.) Seem. ex Bureau | Trumpet-creeper | |
| Calycanthaceae | Calycanthus | floridus | L. | Eastern sweetshrub | |
| Campanulaceae | Lobelia | cardinalis | L. | Cardinal-flower | |
| Caprifoliaceae | Lonicera | japonica | Thunb. | Japanese honeysuckle | |
| Caprifoliaceae | Symphoricarpos | orbiculatus | Moench | Coral-berry | |
| Celastraceae | Euonymus | americana | L. | American strawberry-bush | |
| Clusiaceae | Hypericum | crux-andreae | (L.) Crantz | St. Peter's-wort | |
| Clusiaceae | Hypericum | hypericoides | (L.) Crantz | St. Andrew's-cross | |
| Clusiaceae | Triadenum | walteri | (J. G. Gmel.) Gleason | Greater marsh-St. John's-wort | |
| Cornaceae | Cornus | florida | L. | Flowering dogwood | |
| Cornaceae | Cornus | foemina | P. Mill. | Stiff dogwood | |
| Cornaceae | Nyssa | aquatica | L. | Water tupelo | |
| Cornaceae | Nyssa | sylvatica | Marsh. | Black tupelo | |
| Ericaceae | Rhododendron | canescens | (Michx.) Sweet | Mountain azalea | |
| Ericaceae | Rhododendron | viscosum | (L.) Torr. | Clammy azalea | |
| Ericaceae | Vaccinium | corymbosum | L. | Highbush blueberry | |
| Ericaceae | Vaccinium | fuscatum | Ait. | Black blueberry | |
| Euphorbiaceae | Chamaesyce | maculata | (L.) Small | Spotted sandmat | |
| Fabaceae | Desmodium | nudiflorum | (L.) DC. | Naked-flower tick-trefoil | |
| Fabaceae | Lespedeza | frutescens | (L.) Hornem. | Shrubbery bush-clover | |
| Fagaceae | Quercus | lyrata | Walt. | Overcup oak | |
| Fagaceae | Quercus | nigra | L. | Water oak | |
| Fagaceae | Quercus | palustris | Muenchh. | Pin oak | |
| Fagaceae | Quercus | phellos | L. | Willow oak | |
| Fagaceae | Quercus | rubra | L. | Northern red oak | |
| Fagaceae | Quercus | velutina | Lam. | Black oak | |
| Gentianaceae | Gentiana | sp. | L. | Gentian | |
| Grossulariaceae | Itea | virginica | L. | Virginia sweetspire | |
| Haloragaceae | Proserpinaca | palustris | L. | Marsh mermaidweed | |
| Hamamelidaceae | Liquidambar | styraciflua | L. | Sweet-gum | |
| Juglandaceae | Carya | alba | (L.) Nutt. ex Ell. | Mockernut hickory | |
| Lamiaceae | Lycopus | rubellus | Moench | Taper-leaf water horehound | |
| Lamiaceae | Pycnanthemum | tenuifolium | Schrad. | Narrow-leaf mountain-mint | |
| Lamiaceae | Scutellaria | lateriflora | L. | Mad dog skullcap | |
| Lauraceae | Sassafras | albidum | (Nutt.) Nees | Sassafras | |
| Lemnaceae | Lemna | valdiviana | Phil. | Pale duckweed | |
| Magnoliaceae | Liriodendron | tulipifera | L. | Tuliptree | |
| Menispermaceae | Cocculus | carolinus | (L.) DC. | Carolina coralbead | |
| Moraceae | Morus | rubra | L. | Red mulberry | |
| Oleaceae | Fraxinus | americana | L. | White ash | |
| Onagraceae | Ludwigia | alternifolia | L. | Seedbox | |
| Onagraceae | Ludwigia | palustris | (L.) Ell. | Marsh primrose-willow | |
| Phytolaccaceae | Phytolacca | americana | L. | American pokeweed | |
| Platanaceae | Platanus | occidentalis | L. | American sycamore | |
| Polygonaceae | Polygonum | virginianum | L. | Jumpseed | |
| Pyrolaceae | Chimaphila | maculata | (L.) Pursh | Striped Prince's-pine | |
| Rhamnaceae | Frangula | caroliniana | (Walt.) Gray | Carolina false buckthorn | |
| Rosaceae | Amelanchier | arborea | (Michx. f.) Fern. | Downy service-berry | |
| Rosaceae | Photinia | pyrifolia | (Lam.) Robertson & Phipps | Red chokeberry | |
| Rosaceae | Prunus | serotina | Ehrh. | Black cherry | |
| Rosaceae | Rubus | argutus | Link | Saw-tooth blackberry | |
| Rubiaceae | Cephalanthus | occidentalis | L. | Common buttonbush | |
| Rubiaceae | Diodia | virginica | L. | Virginia buttonweed | |
| Rubiaceae | Galium | circaezans | Michx. | Licorice bedstraw | |
| Rubiaceae | Galium | obtusum | Bigelow | Blunt-leaf bedstraw | |
| Rubiaceae | Mitchella | repens | L. | Partridge-berry | |
| Scrophulariaceae | Gratiola | neglecta | Torr. | Clammy hedge-hyssop | |
| Scrophulariaceae | Gratiola | virginiana | L. | Round-fruit hedge-hyssop | |
| Solanaceae | Solanum | americanum | P. Mill. | American black nightshade | |
| Styracaceae | Styrax | americanus | Lam. | American snowbell | |
| Ulmaceae | Ulmus | rubra | Muhl. | Slippery elm | |
| Urticaceae | Boehmeria | cylindrica | (L.) Sw. | Small-spike false nettle | |
| Vitaceae | Parthenocissus | quinquefolia | (L.) Planch. | Virginia-creeper | |
| Vitaceae | Vitis | sp. | L. | Grape | |
Three hypothetical anthropogenic alterations of hydrologic conditions in Sinking Pond were considered: (1) direct alteration of the spillway or main internal drain, (2) land-use change in the Sinking Pond drainage basin, and (3) waterfowl impoundments in three adjacent small watersheds north, west, and east of the Sinking Pond drainage basin.
Previous geomorphic and hydrologic investigations of the Sinking Pond area found no evidence for human modification of the Sinking Pond spillway or internal drain (Wolfe, 1996a, b). The spillway was traversed several times without noting evidence for disturbance. The broad, flat topography and mature hardwood vegetation of the Sinking Pond spillway contrast sharply with constructed berms in nearby basins (fig. B1). Level notes and discharge measurements made between 1992 and 1995 (Wolfe, 1996a) indicate a channel elevation at the Sinking Pond outflow gage of 323.6 m above NGVD 29 and typical wet-season water-surface elevations of 323.9 m—less than 1 m below the current spillway and more than 2 m above the internal drain; dredging the entire outflow channel to the level of the culvert would affect only sites in the highest one third of Sinking Pond's internal relief. Water levels in Sinking Pond and an adjacent well (fig. 3) indicate that the current internal drain is hydraulically efficient. Artificial blockage of the internal drain appears an improbable mechanism for increasing hydroperiods in Sinking Pond.

Additional field observations from 2000 through 2002 confirm that the spillway and internal drain of Sinking Pond remain in a basically natural condition. Soil borings in both areas reveal gray silty clay or silt loam with distinct soil horizons, hydromorphic features, and a fragipan at 0.5-1 m below land surface. Similar soil profiles are found throughout the pond and are consistent with the mapped Guthrie and Purdy soil series (Love and others, 1959; Edwin Roworth, Arnold Engineering Development Center, written commun., 2001).
Land-use patterns in the Sinking Pond area were mapped using aerial photographs that spanned a 60-year period from 1938 to 1997. For the purposes of this investigation, the Sinking Pond area includes the 335 ha drained by Sinking Pond and the 65 ha downgradient of the Sinking Pond spillway that drains to the Sinking Pond outflow gage (fig. 2). The objective of this work was to evaluate the possible relation between major changes in land use in the Sinking Pond area and hydrologic change associated with tree-regeneration failure documented in this report. Of particular interest were land-use activities, such as ground clearing, soil disturbance, and major changes in ground cover that could affect the basin's hydrologic response.
During the 60-year study period, management of the Sinking Pond catchment was transferred from private ownership (pre-1941) to State ownership (1941-1949), then to Federal ownership (1949-present). During the period of State ownership, the site was heavily utilized for military training exercises associated with Camp Forrest and extensive timber harvesting was conducted by the State caretakers (Jack Bennett, Archeology Associates, written commun., 2002). In 1954, much of the Sinking Pond catchment was designated a Wildlife Management Area, and in 1976 the site was designated a National Natural Landmark by the U.S. Park Service. In the 1960s, the AEDC forestry program began establishing pine plantations on former agricultural fields in the catchment (David Bynum, Arnold Air Force Base, written commun., 1997).
Changes in land use in the Sinking Pond area were determined by creating and analyzing a land-use/land-cover GIS database. This process involved the following steps: (1) acquiring imagery, (2) scanning and rectifying imagery, (3) digitizing land-use/land-cover polygons, and (4) assigning land-use/land-cover categories to each polygon across the 60-year temporal sequence. Table B1 shows the dates and sources of aerial photographs used in this analysis. The 1938 and 1943 imagery was received in a usable, georeferenced digital format. The 1950-1992 imagery was obtained in hard copy form from the Tennessee Valley Authority. Each image was scanned using a UMAX Powerlock 2100xl (at 400 dpi) and rectified to the USGS 1997 Manchester digital orthophoto quadrangle (DOQ) using ArcView Image Analysis software.
| Source | Date |
|---|---|
| National Archives1 | April 1938 |
| National Archives1 | June 18, 1943 |
| Tennessee Valley Authority | February 16, 1950 |
| Tennessee Valley Authority | April 1, 1960 |
| Tennessee Valley Authority | February 26, 1969 |
| Tennessee Valley Authority | November 3, 1971 |
| Tennessee Valley Authority | March 22, 1976 |
| Tennessee Valley Authority | March 22, 1984 |
| Tennessee Valley Authority | December 13, 1992 |
| Tennessee Valley Authority | March 29, 1997 |
1National Archives materials obtained through Arnold Air Force Base.
The perimeter of the Sinking Pond outflow basin was scanned from Wolfe (1996a) and rectified against the 1997 Manchester DOQ. This area represents the 335-ha catchment of Sinking Pond plus an additional 65 ha drained by the Sinking Pond outflow channel below the pond's spillway. The perimeter boundary then was digitized and used to define the area for land-use determinations in each image. A total of 100 land-use polygons, which maintained relative homogeneity throughout the period of analysis, were identified and digitized. Each polygon was examined in each of the 10 sequential images and was assigned to one of seven land-use/land-cover classes (table B2) for each of the 10 years. Prior to classification of the polygons, the seven land-use/land-cover classes were field checked using the 1997 photos.
Table B2. Land-use/land-cover classes used in land-use analysis of the Sinking Pond area.| Land-use/land-cover class | Definition |
|---|---|
| Hardwood, 70-100 percent forest cover | Hardwood forest with 70 to 100 percent continuous canopy cover; forest habitat with minimal evidence of recent human disturbance. |
| Hardwood, 40-70 percent forest cover | Hardwood forest with 40 to 70 percent continuous canopy cover; low-density canopy such as found in savanna-like habitat or recent selective logging activity; some disturbance of soil surface may be present (less than 10 percent mineral soil exposed). |
| Disturbed ground, 10-40 percent forest cover | Forest with 10 to 40 percent continuous canopy cover; early successional habitat (mixture of tree, shrubs, and herbaceous cover); strong evidence of vehicular or other disturbance of soil surface (10 to 40 percent mineral soil exposed). |
| Evergreen succession | Early- to mid-successional habitat; dominant canopy species is red cedar; some disturbance of soil surface may be present (less than 10 percent mineral soil exposed). |
| Pine plantation | Various stages of loblolly pine plantation development with varying degrees of pine canopy cover; varying degrees of soil disturbance, depending on stand age. |
| Grassland | Predominantly herbaceous vegetation with less than 10 percent tree cover; strong evidence of vehicular or other disturbance of soil surface (10 to 40 percent mineral soil exposed). |
| Bare ground | Cleared land with 40 to 100 percent mineral soil exposed; lands in crop rotation, in preparation for pine plantation, or receiving high off-road vehicular traffic such as traffic associated with military training exercises. |
The GIS analysis of the land-use patterns over the 60-year study period indicates that human disturbance within the Sinking Pond area reached a maximum level around 1943, with less than 25 percent of the land area having a hardwood forest cover of 40 percent or greater (fig. B2). During this same time, 57 percent of the landscape was in a heavily disturbed and erosion-prone condition (cleared bare and disturbed 10-40 percent) compared to 23 percent in 1938. Visual inspection of the imagery indicates that this increase can be attributed to heavy clearcutting in and around Willow Oak Swamp and around the perimeter of Sinking Pond and to off-road vehicle traffic and explosion impacts on the west side of the drainage.
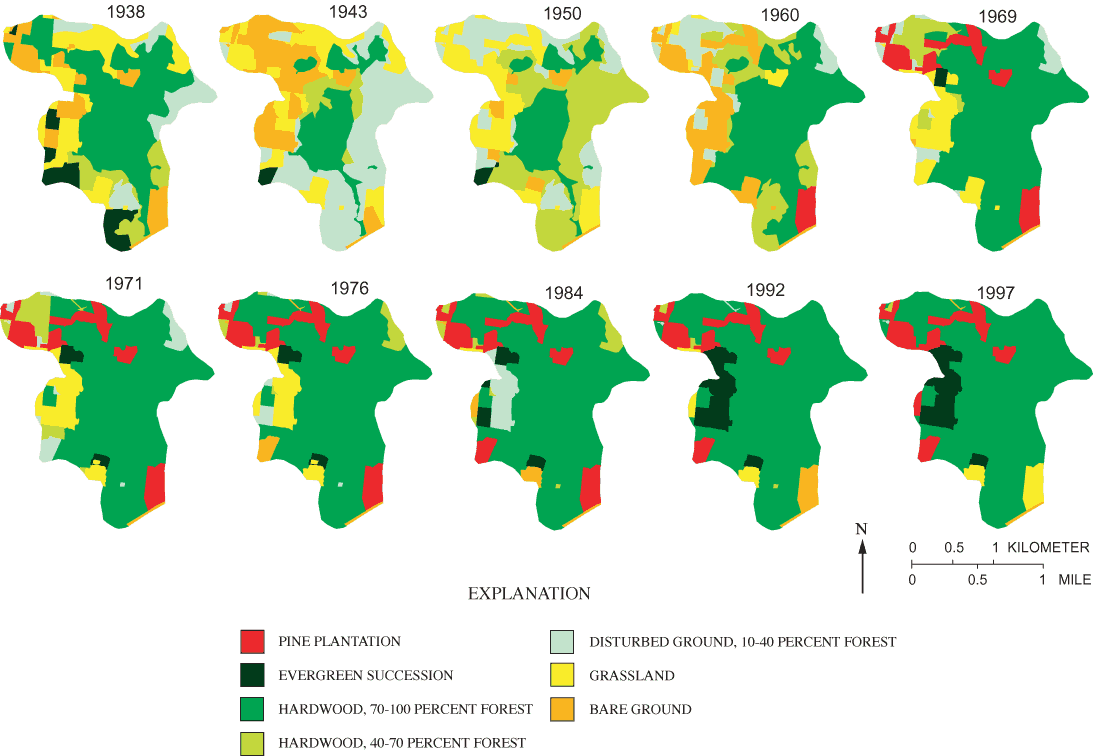
Figure B2. Changes in land cover in the area drained by the Sinking Pond outflow gage, 1938-1997.
From 1950 to 1997, there was a steady increase in the percent of landscape with continuous forest cover and a decrease in overall soil disturbance. In 1960, there was a brief increase in cleared-bare area as former agricultural fields were converted to pine plantation. By 1971, however, 85 percent of the land area had hardwood forest cover greater than 40 percent or contained established pine plantations, and areas of bare ground represented less than 1 percent of the basin (fig. B2). None of the photos indicate direct disturbance of the ponded area of Sinking Pond during the 60-year period.
The general relation of vegetation structure to the hydrologic cycle in humid, mid-latitude settings is well established from many studies (Wilde, 1958; Pereira, 1973; Pritchett, 1979). Other factors being equal, forests have higher infiltration and soil-water retention (Heyward and Barnette, 1934; Pereira, 1973; Pritchett, 1979), less runoff (Hibbert, 1967), greater interception and evapotranspiration losses (Johnson, 1964; Rutter, 1972; Stanhill, 1970), lower water yields (Hewlett and Hibbert, 1961; Hibbert, 1967; Hornbeck and others, 1970), and lower ground-water levels (Trousdell and Hoover, 1955) than areas under row crops, pasture, savanna, or bare ground. Stands of evergreen conifers exhibit similar hydrological differences in comparison to deciduous hardwoods (Johnson, 1964; Stanhill, 1970; Helvey, 1974; Swank and others, 1972; Swank and Douglas, 1974).
Land-use changes in the Sinking Pond basin from 1943 through 1997—a steady increase in forested area and establishment of pine plantations (fig. B2)—could be expected to lower the water table, reduce runoff and throughflow, and increase evapotranspiration. These hydrologic effects would reduce, not increase, the hydroperiod in Sinking Pond resulting from a given annual rainfall. Neither the altered patterns in tree regeneration indicated by the botanical evidence discussed above nor the hydrologic changes that apparently have driven those patterns can be attributed to land-use change.
In the 1960s, berms were built across Huckleberry Creek, Dixie Flats, and Westall Swamp, directly to the north, east, and west of the Sinking Pond basin, respectively (fig. 2). The berms were intended to provide habitat and resting places for waterfowl and are equipped with control structures to permit seasonal draining (Mark Moran, Arnold Engineering Development Center, written commun., 2001). The hypothesis was considered that the berms might potentially increase ground-water recharge or compete with Sinking Pond for drainage. An extreme example of such an effect would be the case of two basins each having highly conductive internal drains leading to the same karst conduit. In such a case, artificially ponding water in one basin would pressurize the other basin's internal drain, creating an effect similar to that of a backed up drain in a bathtub.
The hydraulic connection between the Sinking Pond basin and one of the neighboring impounded basins was examined through a simple impoundment-draining experiment. Continuous water-level recorders were installed behind the Huckleberry Creek berm, at a seasonally filled vertical shaft near the divide between the Huckleberry Creek and Sinking Pond basins, and in three wells north of Sinking Pond (fig. 2). The control structure at Huckleberry Creek dam was closed, and the area behind the dam was allowed to fill. The control structure was opened on February 13, 2002, and the water-level response in Sinking Pond, the wells, and the vertical shaft was monitored as the pond drained.
Water levels at Huckleberry Creek dam, Sinking Pond, and the monitored wells and vertical shaft are shown in figure B3. There is no indication that draining Huckleberry Creek dam had any effect on the other stations. Short-term recessions from minor rainfall on February 7, 2002, began before Huckleberry Creek dam was drained and continued with no obvious change in slope through the drainage event.
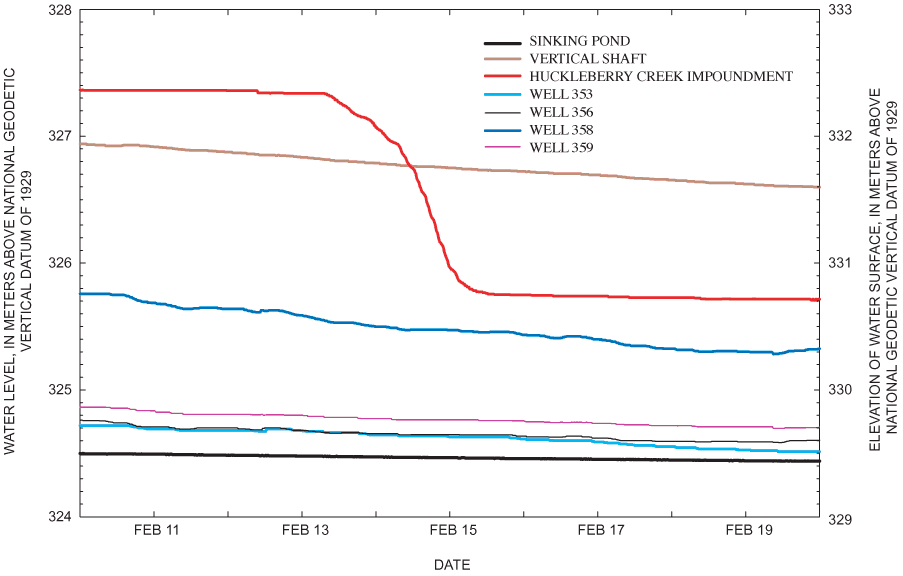
Lack of a strong signal does not mean that the berms have had no effect on the Sinking Pond hydroperiod. Even after drainage, some water remained ponded behind Huckleberry Creek dam. Similarly, the berms at Dixie Flats and Westall Swamp retain water even with the control structures open. The artificially ponded water imposes hydrostatic pressure on the local ground-water system and may marginally increase seasonal recharge (C.J. Haugh, U.S. Geological Survey, Nashville, Tenn., written commun., 2002). The effects of such changes across basin divides, however, cannot be demonstrated and appear to fall short of the abrupt, high-magnitude hydrologic change implied by the ecological and tree-ring data for Sinking Pond.
| Basin centimeter | The volume of 1 centimeter of precipitation distributed over a drainage basin. |
| Biodiversity | The number, variety, and variability of species in an ecosystem. |
| Canopy | The cover formed by the branches and leaves of forest trees. |
| Cohort | A group of organisms of the same species and age, for example, a group of overcup oak trees that germinated in a single year. |
| Disjunct | An isolated population of plants or animals far from their typical geographic range. |
| Endemic | Native to or typical of a particular region or ecosystem. |
| Fragipan | A subsurface soil layer of relatively high density and low permeability to water. |
| Hydroperiod | The amount of time a wetland is covered by or saturated with water. |
| Karst | A landscape in which chemical dissolution has enlarged joints, fractures, bedding planes, or other openings in soluble, underlying bedrock; may be characterized by sinkholes, caves, and disappearing streams. |
| Karst conduit | A discrete, solution-enlarged opening in karst bedrock large enough to permit turbulent flow, typically greater than 5 millimeters across. |
| Recruitment | The transition of an individual organism into one of several successive developmental stages, such as acorn to seedling, seedling to sapling, or sapling to mature tree. |
| Regeneration | The maintenance of a biological population through the reproduction and survival of new individuals. |
| Vadose zone | The zone between the land surface and the dry-season water table. |
| Water budget | The balance between water inputs, outputs, and changes in storage during a fixed time interval. |
Document Accessibility: Adobe Systems Incorporated has information about PDFs and the visually impaired. This information provides tools to help make PDF files accessible. These tools convert Adobe PDF documents into HTML or ASCII text, which then can be read by a number of common screen-reading programs that synthesize text as audible speech. In addition, an accessible version of Acrobat Reader 6.0, which contains support for screen readers, is available. These tools and the accessible reader may be obtained free from Adobe at Adobe Access.
| AccessibilityFOIAPrivacyPolicies and Notices | |
 |
|