Ecologists are concerned with interrelations between biological communities and their habitat. A variety of biological, chemical, and physical factors control the abundance, distribution, and productivity of aquatic organisms (Gordon and others, 1992, p. 17). Competition for preferred habitat, predation, water chemistry, nutrient inputs, the presence of waterfalls or dams, and streamflow variability are important factors for stream biota (Gordon and others, 1992, p. 17). The preceding section of this report introduced chemical water quality and summarized one of its constituents. In this section, selected physical and biological features of stream ecology in the YRB is discussed.
The physical character of rivers within the study unit changes markedly from headwater streams to lowland streams. Upstream reaches are characterized by turbulent flows, steep gradients, cold water temperatures, coarse substrates, and well-oxygenated water, whereas lowland reaches are typically characterized by warmer water temperatures (especially during summer), gentle gradients, turbidity, sediment deposition, fine substrates, and smaller concentrations of dissolved oxygen.
The physical factors that are of greatest ecological significance include streamflow (discharge and velocity), channel shape, channel substrate, and water temperature (Gordon and others, 1992). Stream discharge, including the ecologically important periods of floods and droughts, was discussed in a previous section of this report. Streamflow velocities are a function of flow depth and gradient or slope in the downstream direction. To allow comparison of streamflow velocities among different sites or streams, a discharge of approximately constant frequency of occurrence is referenced at all sites; commonly, the mean annual discharge is used.
Leopold and Maddock (1953) tabulated mean streamflow velocities at mean annual discharge for 46 streamflow-gaging stations in the YRB, and several summary observations may be made from those data, as follows. Average velocity increases in the downstream direction along the upper through middle reaches of the principal rivers, as it does typically due to increasing flow depths. But velocities decrease in the lower reaches of the Bighorn, Powder, and Yellowstone Rivers. The Wind/Bighorn River mainstem has an average velocity between 0.9 and 1.1 m/s for much of its length upstream from Bighorn Lake, but on the Great Plains the Bighorn and Little Bighorn Rivers have average velocities of about 0.7 m/s. Also on the Great Plains, average velocities in the several forks of the Powder River near their confluences with the mainstem are less than 0.6 m/s. In a middle section of the Powder River mainstem, average velocity increases to about 1.0 m/s, but then decreases to less than 0.9 m/s in its lower reaches. In contrast, average velocity in the Yellowstone River at Corwin Springs, Mont., is 1.1 m/s, and increases to 1.5 m/s at Billings, Mont., but then decreases to just less than 1.0 m/s at Sidney, Mont. (Leopold and Maddock, 1953).
The streamflow velocities actually encountered by biota are more relevant than average velocity. But large local variability produces a mosaic of velocity patterns that support species with differing preferences (Gordon and others, 1992, p. 20). The velocity patterns, and related turbulence of flows, are difficult to quantify for an individual river, much less over a large drainage such as the YRB.
Channel shape is a function of the width and depths of streamflow. As related to flow depth, it influences water temperature, light penetration, and fish migration (where depth is too shallow for passage). As related to width, it influences the degree of shading by overhanging bank vegetation, instream photosynthesis, and input of organic matter (Gordon and others, 1992, p. 21). The width-to-depth ratio (W:D) commonly is used as an index of channel shape.
Osterkamp and Hedman (1982) presented channel geometry measurements, including active-channel width and average bankfull depth, for 21 stream stations in the YRB. For three streams with drainages primarily in the Wyoming Basin Province, W:D was 11.1 or less, indicative of incised channels. Seven cobble-bed streams with primarily mountainous drainages had W:D averaging 29.4 and ranging from about 20 to 51. On the Great Plains, six sand- or gravel-bed streams had W:D averaging 31.2 and ranging from 24 to 43 (Osterkamp and Hedman, 1982). Both of the sand-bed sites were on the Powder River. For comparison, sand-bed streams of some other Great Plains river systems (Platte and Republican Rivers) commonly have W:D greater than 100. The sand-bed channels of the Powder River contain more silt and clay in their beds than do those of the other two river systems, and such cohesive materials tend to provide greater resistance to erosion. The interrelations among physical habitat characteristics become apparent when seeking to understand the variation in these factors.
Stream substrate, or the particle-size distribution of bed material, is associated with streamflow velocity and, as implied above, influences channel shape. Substrate is an important factor controlling the occurrence of benthic (bottom-dwelling) fauna, and the complex mixture of coarse particles found in riffles provides the richest habitat for aquatic insects (Gordon and others, 1992). Although both the average and range of particle sizes, and their arrangement on the streambed, are ecologically important, data are most readily available for the median particle size (d50).
In addition to substrate data for 19 stream stations in Osterkamp and Hedman (1982), the authors measured d50 for 15 stations while conducting a reconnaissance of YRB streams in 1997. These measurements were based on pebble counts performed using the half-largest-particle grid interval method of sampling (Wohl and others, 1996). Data from both sources were considered together to analyze the overall pattern of average particle sizes.
Three stations draining the Wyoming Basin Province have an average d50 of 6 mm, but substrates range from silt to coarse gravel. The exposed soil area around sparse vegetation in drier parts of the Wyoming Basin allows sheet-and-rill erosion to wash fine sediment into the stream channels.
Particle size generally decreases in the downstream direction, and data for the YRB streams are mostly consistent with this generalization. Among 15 stations with predominantly mountainous drainages, the average d50 is 125 mm, with substrates ranging from coarse gravel to medium boulders. Five stations that are transitional between mountains and plains drainages have median particle diameters ranging from 38 to 58 mm. On the Great Plains, the average d50 for 11 YRB stations is 12 mm, with substrates ranging from very fine sand to very coarse gravel. An exception to the trend of downstream fining of bed material occurs in the lower Powder River between Arvada, Wyo., and Locate, Mont., where d50 coarsens from 0.16 to 0.35 mm (Osterkamp and Hedman, 1982).
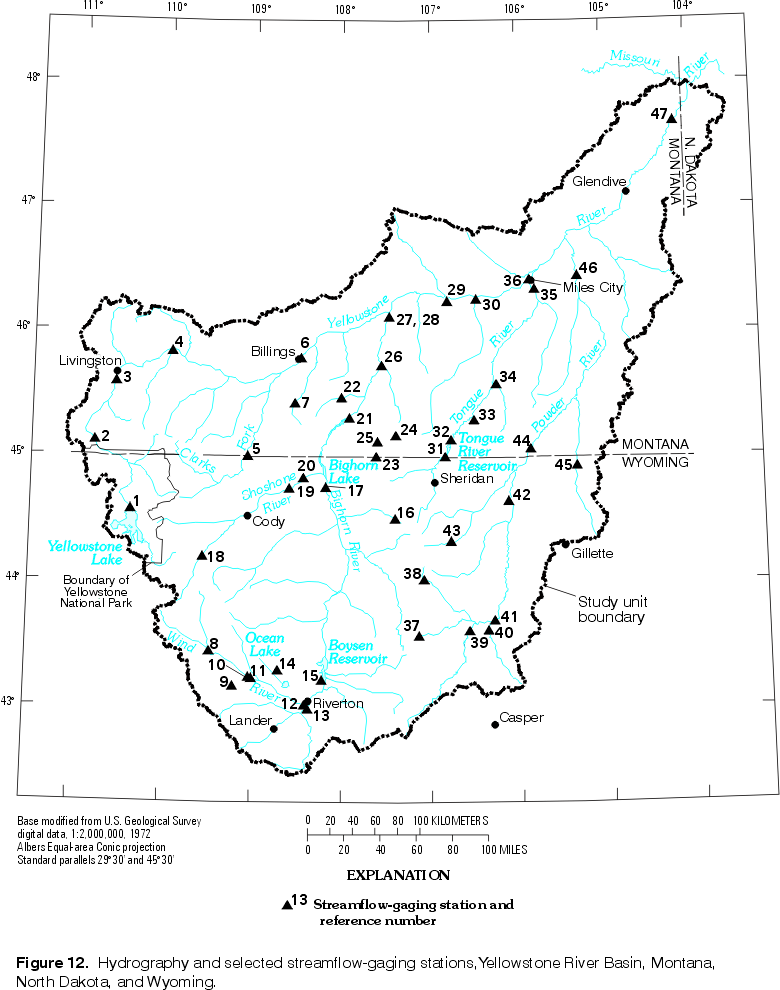
Water temperature varies both seasonally and daily in concert with air temperature, but it generally increases in the downstream direction as the cooler, high-elevation climate is displaced by the lowland climate. This pattern is seen in YRB streams, particularly with respect to mean water temperature in the warmest months. Available USGS data for the 14 stream stations listed in table 6, and for the Gardiner River near Mammoth in Yellowstone National Park, were summarized by month. The period of record used generally was 1970-94, although the period was slightly different for a few stations; however, for the Gardiner River, the record extends from 1985-96. Mean water temperature is warmest in July, with five exceptions: August has warmer mean water temperature than July for sites 5, 6, 28, 46 (fig. 12), and for the Gardiner River site. Site 46 has equally warm mean water temperature in June and August; all other sites have a single peak on the curve of monthly mean temperatures. Mean water temperature in the warmest month ranges from about 16.5°C at sites 3 and 5 to about 23.5°C at sites 35, 41, and 42.
Mean water temperature is coolest in January, with five exceptions: February has cooler mean water temperature than January for sites 6, 28, 42, and 45, and December has the coolest mean water temperature for the Gardiner River site. Mean water temperature in the coolest month generally has a small range, from 0°C at sites 12, 13, 35, and 46 to about 1.3°C at sites 5, 6, and 28. In contrast, the geothermally influenced Gardiner River has warmer mean water temperatures for December through February, ranging from 10.3°C to 11.3°C. Cold winter water temperatures in the northeastern part of the YRB reflect a continental climatic gradient, whereas those in the Wind River Basin are indicative of its higher elevation.
Longitudinal zonation of species occurs primarily with respect to altitude, water depth, and substrate composition in relation to stream gradient. High-gradient headwater streams are termed "salmonid" (after Family Salmonidae--Trouts, includes whitefishes, trouts, salmons, chars, and graylings) or rhithron zones, whereas lowland streams are termed "cyprinid" (after Family Cyprinidae--Minnows, includes minnows, shiners, dace, and chubs) or potamon zones. A transition zone, characterized by intermediate levels of both erosion and deposition, moderate gradients, alternating riffle-pool sequences, and sand-gravel substrates separates the two.
Biotic zonation was originally defined by the apparent relation between a stream's gradient and the fish species harbored therein. However, algae and invertebrate studies have since shown that the complete assemblage of fauna shows zonation, both in community composition and in numbers of species (Hynes, 1970). For example, Newell (1977) sampled macroinvertebrate communities at 20 stations along the Yellowstone River. He defined the salmonid zone to extend downstream to about 680 km upstream of the mainstem mouth, and the transition zone to extend 200 km further downstream to the confluence with the Bighorn River. Newell's (1977) downstream boundary for the salmonid zone roughly corresponds to the physiographic transition from mountains to plains.
Downstream variations in aquatic biota also have been explained using the river continuum concept (Vannote and others, 1980), relating stream order (Strahler, 1957), size of particulate organic matter, and the ratio of photosynthetic production to community respiration. Low-order streams (for example, first-order tributaries in the forested headwaters of the Yellowstone River) typically receive substantial terrestrial contributions of coarse particulate organic matter (CPOM) and have little photosynthetic production. Food chains in intermediate-sized streams are more dependent on photosynthetic production along with fine particulate organic matter (FPOM) from upstream shredding of CPOM by invertebrates (Newell, 1977). Greater turbidity in large rivers (such as downstream reaches of the Powder and Yellowstone Rivers) limits photosynthetic production such that FPOM becomes the principal base of the food chain.
Algae are ubiquitous, autotrophic, unicellular or multicellular organisms maintaining reproductive characteristics that distinguish them from liverworts, mosses, and vascular plants. Algae are central to aquatic ecosystems (Bahls and others, 1984) as producers of carbohydrates and proteins via photosynthetic processes and are an essential component of the riverine food chain (Round, 1973). Benthic species are most important when considering a stream's primary production and plant diversity, and may be of epilithic (attached to stones), epipelic (attached to mud or sand), epiphytic (attached to plants), or epizoic (attached to animals) habits (Bold and Wynne, 1985). In flowing water, planktonic (open water) algal species are of lesser biological significance because their presence depends largely upon flow regime, catastrophic drift, and tributary inoculation.
In a variety of ways, algae and other photosynthetic plants are indicative of stream water quality (Whitton, 1979). Historically, biologically based surveys of streams, as opposed to those of lakes and reservoirs, have used animals, both vertebrates and invertebrates, as indicators. However, as pollutant concentrations meeting standards formulated to protect fish may be lethal to other aquatic species, modern bioassays generally use at least three food-chain components: algae, invertebrates, and fish or amphibians (Whitton, 1979). Changes in algal community structure, population, biomass, photosynthetic and respiration rate, nitrogen-fixation rate, chemical composition, and morphology may all signify water-quality changes within a given reach. Similarly, water quality may be questionable if a widely distributed species is not found during sampling (Bahls and others, 1984). A stream's general water quality may best be indicated by benthic organisms.
Discharge, current velocity, substrate, turbidity, scour, suspended solids, nutrient status, dissolved oxygen, dissolved salts, light intensity, pH, hardness, toxicants, water temperature, and grazing animals are all factors influencing riverine algal communities. Community structure is strongly affected by the seasonal periodicity of species, with annuals achieving maximum development, abundance, and reproduction during specified seasons, perennials maintaining a continuous vegetative cycle, and ephemerals appearing, opportunistically, for short times during any season (Smith, 1950). Correspondingly, dispersal agents (such as wind, birds, amphibia) (Smith, 1950), catastrophic events, origin of water (spring, standing, or tributary), and riparian vegetation will vary community composition (Round, 1973; Bahls and others, 1984). Lastly, although limited algal species succession occurs, the traditional plant community concept as defined by terrestrial ecologists does not generally apply to streams; algal associations are difficult to delimit.
The divisions of algae most common in the YRB include:
Bacillariophyta (diatoms) -- Diatoms are the predominant algae in streams draining the montane portions of the YRB (Robinson and others, 1996). Diatoms are essentially cold-water organisms, common during spring and autumn, and live on substrates including rock, sand, mud, and water plants, or as epiphytic growth on other algae. Planktonic habits of these essentially unicellular algae also are known (Bold and Wynne, 1985).
Charophyta (stoneworts) -- These algae are usually found in freshwater, anchored on muddy or sandy substrates or hard limestone stream beds (Bold and Wynne, 1985). Many species become encrusted with carbonates, mainly of calcium and magnesium; hence suggesting their common names
(stoneworts or brittleworts). Some species prefer brackish water, but most thrive best in clear, hard water; aeration is not essential.
Chlorophycota (green algae) -- These algae are found in a variety of settings including standing, tranquil, and swiftly flowing water (cataracts, waterfalls, dam spillways). They inhabit water having a great range of salinity, and both benthic and planktonic species are known (Bold and Wynne, 1985).
Chrysophyta (golden-brown algae) -- These algae typically are restricted to unpolluted, cold streams and springs.
Cyanophycota (blue-green algae) -- These algae are generally more abundant in neutral or slightly alkaline water, but are found in water having a great range of salinity and temperature (Bold and Wynne, 1985). Both planktonic and benthic species are included in this division. Blue-green algae inhabit alkaline hot springs in Yellowstone National Park having water temperatures as warm as 74°C (Bold and Wynne, 1985).
Euglenophycota (euglenoids) -- These single-celled algae are widely distributed, being found in freshwater, brackish water, and on moist soils and mud (Bold and Wynne, 1985). Euglenoids usually are abundant in small pools rich in organic matter, especially those to which livestock have access.
Pyrrophycophyta (dinoflagellates) -- Although generally marine, freshwater dinoflagellates occur in pools, ditches, and small lakes with considerable vegetation. The unicellular algae of this division are widely known for the blooms known as "red tides" that they produce in some marine
locations.
Rhodophycota (red algae) -- Although generally marine, freshwater species are restricted to cold, well-aerated water of rapids, falls, and spillways
(Bahls and others, 1984). Many species do not appear reddish at all, and a full range of pigmentation occurs among freshwater species (Bold and Wynne, 1985).
Xanthophyta (yellow-green algae) -- In streams, habits of these algae are epiphytic; terrestrial members of this division may be found growing in dense stands on drying mud along stream banks.
Algae found in cold, mountain streams within the YRB are more distinctive, and include a larger percentage of species restricted to a particular habitat, than those found in warmer, lowland streams (Smith, 1950). Algae found in rapids and waterfalls usually are attached to stones (Smith, 1950). Additionally, rocks continuously moistened by spray from rapids and waterfalls, or where there is a continuous trickle of water, develop extensive feltlike or gelatinous algal masses upon rocky cliffs (Smith, 1950). Certain species of blue-green algae are the most frequently encountered algae on dripping cliffs (Smith, 1950). Algae of high-gradient streams are of two distinct morphological types: encrusting algae (diatoms, for example) and those in which the greater part of the thallus (undifferentiated body) trails in the current (Smith, 1950). Certain red algae and river mosses (Cladophora) are restricted to cold, high-gradient streams at elevations higher than approximately 3600 m (Smith, 1950).
The algal flora of lowland streams with fine-grained substrates are far less diverse than those of mountain streams (Smith, 1950). Most lowland flora are attached to mud or sand, and include diatoms, blue-green algae, green algae, and euglenoids. In contrast to encrusting and trailing algae upstream, these species are often motile.
As with algae, the composition of invertebrate communities varies longitudinally downstream within the YRB. Overall, factors influencing both distribution and abundance of aquatic invertebrates include current velocity, water temperature, substrate, stability of both aquatic and riparian vegetation, dissolved substances, competition, zoogeography, food, disturbance history, and human practices. Water temperature and chemistry exert great influence, but invertebrate morphological features and respiratory requirements, in relation to streamflow habitat, ultimately define macroinvertebrate community composition. Large, stable substrates such as boulders and cobbles support larger, more productive invertebrate populations than do unstable gravel and sand substrates. Consequently, a longitudinal decrease in substrate size and riffle frequency results in lower macroinvertebrate production and diversity downstream. Although deposition of organic sediment at slow current velocities may increase benthic production for midges (Chironomidae) and earthworms (Oligochaeta) that depend on allochthonous detritus (Baril and others, 1978), the filling of interstitial spaces with fine, inorganic sediment eliminates potential habitat.
Within the YRB, invertebrate fauna are likely be dominated by mayflies (Ephemeroptera , includes both mountain and prairie varieties) whose current-velocity preferences vary; for example, Baetis prefer rapid currents whereas Tricorythodes prefer slow velocities. Other prominent taxa include caddisflies (Trichoptera), many of whom, such as Hydropsyche , depend on rapid currents for proper net functioning (Baril and others, 1978); true flies (Diptera) (Newell, 1977); and stoneflies (Plecoptera), whose general preference is for rapid currents and whose fauna is diverse but not abundant (Newell, 1977). Another swift-current specialist is the riffle beetle (Elmidae). Invertebrates present in large numbers in transition-zone streams of the YRB generally are tolerant of a wide range of habitat conditions, whereas those abundant in prairie reaches are considered tolerant of turbid, silty conditions (Baril and others, 1978).
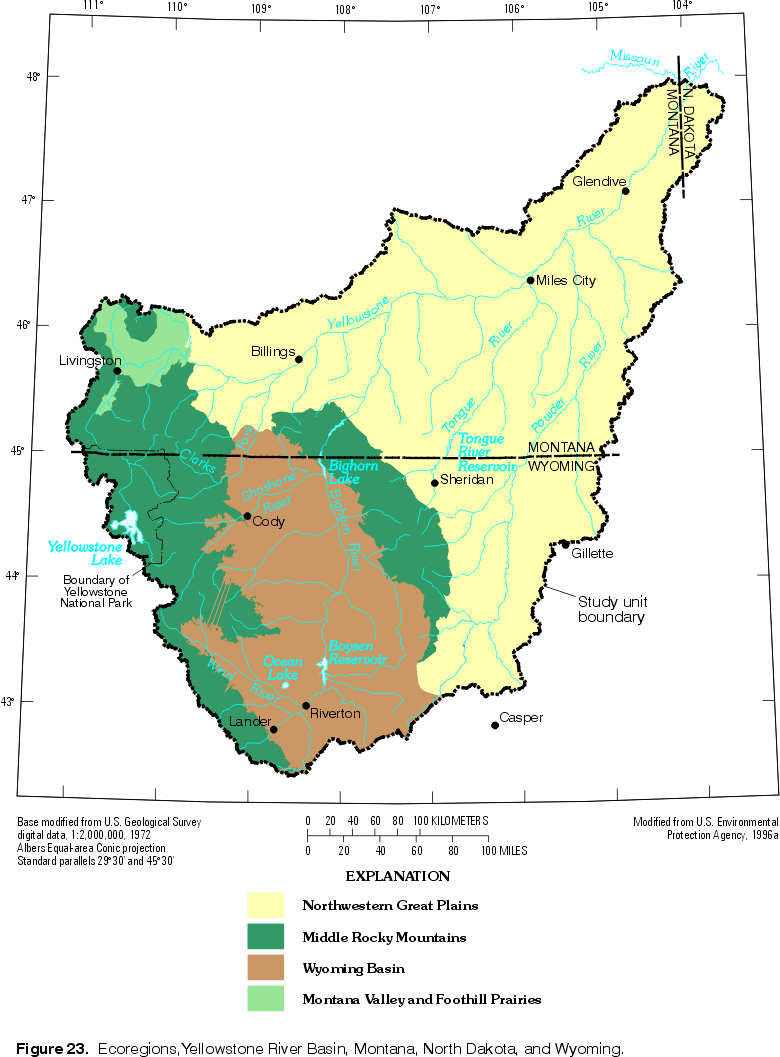
Macroinvertebrate distribution and abundance often vary widely through the year in response to seasonal flow variations. Dispersal is affected by floods through channel scouring and catastrophic drift that can transport invertebrates that would not drift otherwise (Peterson, 1990). Broadly, within each ecoregion of the YRB (described later and shown in fig. 23), invertebrate communities of all perennial streams will be similar; moreover, communities of ephemeral and intermittent streams will resemble one another (Peterson, 1990).
The distribution of fish within the YRB is influenced by elevation, drainage divides, and other physiographic features (Baxter and Stone, 1995). As with other aquatic communities, both abiotic and biotic factors control the occurrence, interspecific relationships, numbers, and growth rates of fish. Abiotic factors include dissolved oxygen, water temperature, current velocity, discharge fluctuation, dissolved salts, substrate, and turbidity. Information on the effect of biotic factors (numbers and growth rates of competitors) is much less available, although closely related species interact with one another in such a way as to divide habitat between them (Hynes, 1970). Of abiotic factors, water temperature is important to cold- and warm-water fish species and indirectly influences oxygen consumption; water chemistry seems to be of lesser importance in natural systems (Hynes, 1970).
Warm-water species considered native to the YRB include the goldeye, brassy minnow, fathead minnow, flathead chub, sturgeon chub, western silvery minnow, white sucker, shorthead redhorse, stonecat, burbot, channel catfish, and sauger. Of these, the stonecat may be found in high-gradient reaches while the goldeye, flathead chub, western silvery minnow, burbot, and sauger all prefer larger, sluggish, deeply turbid streams. The channel catfish prefers larger rivers with turbid habitats. The shorthead redhorse, white sucker, brassy minnow, fathead minnow, and sturgeon chub may all be found in transition zones that generally have less turbidity, some vegetation, and gravel substrates.
Cold-water species native to the study unit but also found on the western side of the Continental Divide include the mountain whitefish, cutthroat trout, arctic grayling, and mottled sculpin. Species common to cool-water, transition-zone streams include the lake chub, longnose dace, longnose sucker, mountain sucker (Brown, 1971; Baxter and Stone, 1995). Of these, the lake chub is usually found in smaller streams, and the mountain sucker inhabits a wide variety of habitats, including large rivers and mountain streams. The longnose dace prefers the swift-water environment of riffles, and the longnose sucker prefers clear water.
In the YRB, several species of fish are accorded special concern by the Federal or State governments. The pallid sturgeon, a Federally listed Endangered Species, occurs in the lower Yellowstone River. The sturgeon chub and arctic grayling currently are designated as Candidate species for Federal Endangered or Threatened status (Fertig, 1997). The Yellowstone cutthroat trout is designated a Sensitive Species by the U.S. Forest Service. Species of special concern to State governments include the pallid sturgeon, paddlefish, Yellowstone cutthroat trout, arctic grayling, sturgeon chub, sicklefin chub, northern redbelly x finescale dace (hybrid), and blue sucker. Most of these are warm-water species inhabiting the lower Yellowstone River, but the Yellowstone cutthroat trout and the arctic grayling are cold-water species.
Return to Environmental Setting of the Yellowstone River Basin, WRIR 98-4269
{kind=link}
{kind=link}