 View figure 3. or
download the postscript file.
View figure 3. or
download the postscript file.Black Rock core stratigraphy plotted against the magnetostratigraphy, macrofossil occurrences, and magnetic susceptibility. BR = Brunhes, J = Jaramillo, C = Cobb Mountain, O = Olduvai, R = Réunion, X = Anomaly X, K = Kaena.
Sediments in the Black Rock core are primarily calcareous mud that typically has dry colors of light gray to pale yellow (Fig. 3). At the time of drilling the mud was largely unoxidized and was greenish gray to black. The mud is massive, to finely bedded (beds between 5 and 8 mm thick and separated by thin silt or very fine sand partings), to laminated (laminae less than 5 mm thick). Soft-sediment deformation is common throughout the length of the core in intervals less than about one foot in thickness. Except for laminated zones, most bedding is wavy. Laminated intervals are several inches to several feet in thickness, and the laminations are laterally discontinuous. Mud samples from the upper 400 feet of the core average approximately 40% carbonate minerals and 60% mud, with small amounts of sand in some samples. Carbonate content is generally inversely correlated with mud content, and carbonate ranges from about 30 to 80%. Preliminary X- ray diffraction studies indicate that the carbonate is in the form of dolomite and calcite.
There are three sandy intervals in the core, although thin sand beds or laminae are present throughout the core. The three sandy intervals are approximately from 0 to 100 ft, from 350 to 450 ft, and from 750 ft to the bottom of the core at 897 ft (Figure 2). Most of the sand is fine to very fine and is flat laminated to low-angle ripple-laminated, to finely cross-bedded. The sand is medium-grained in the upper 35 ft. Sand throughout the core is uncemented except for four thin intervals, each about 2 inches thick, between 29 and 36 ft, where it is well cemented with calcite. In the lower part of the core, from 865 to 875 ft, the fine sand is cemented with gypsum. Between depths of 858 and 861 ft the sand is composed of rounded gypsum grains that were probably transported to the depositional site by wind or some other mechanism. Euhedral selenite crystals and selenite veins are common throughout the core. Most of the sand in the core is poorly sorted, interlaminated with mud, and less than one inch thick, except for the medium sand in the upper part of the core. Analyses of samples from the upper 400 feet indicate that sand content ranges from 0 to over 80% in the upper 100 feet, but ranges between 0 and 40% below 100 feet. Sand percentage is zero in many samples, especially between 100 and 350 ft (Figure 2).
No evidence of soil development or extended periods of drying was observed anywhere in the core. A few features interpreted as mud cracks are poorly developed and probably indicate minor hiatuses. The sandy to silty, finely-bedded sediments suggest shallow-water deposition. In addition, vertical, irregular tube-shaped color stains, which are interpreted as root traces, probably from aquatic plants such as Ruppia, also suggest shallow water. Ruppia seeds are commonly present in the same intervals as the root traces and are distributed throughout much of the core, especially in the mud intervals (Fig. 3). Impressions of filamentous leaves, possibly of Ruppia, were logged at about 495 and 817 ft, and gastropods were observed between 51 and 63 ft.
Taken together, the observations suggest that a shallow lake existed at the Black Rock site for the entire period recorded by the core. Shifts in sand content are related to changes in the sediment influx rate, which may have been caused by changes in the depositional sites of rivers or streams that entered the lake, or by changes in lake level or tectonics. Uplift in the vicinity of Cove Creek dome, 18 km east of the Black Rock site, is known to have occurred in late Pliocene time (Crecraft and others, 1981), and possibly as recently as the late Pleistocene (Oviatt, 1991). Uplift, delta progradation, or lake-level lowering would each cause an increase in sand deposition.
The magnetic polarity stratigraphy obtained using the data interpretation procedures outlined above is presented in Figure 4 and in Appendix 2. As mentioned above, the Bishop ash (0.759 Ma) is preserved in silts that were deposited in hollows on the surface of the Black Rock lava flow. The normal polarity directions obtained from the lacustrine record immediately below the Black Rock basalt are therefore interpreted as representing the lowermost part of the Brunhes Chron (< 0.78 Ma). This interpretation is inconsistent with K-Ar age determinations of 0.97 � 0.25 (Condie and Barsky, 1972) and 1.32 � 0.09 Ma (Crecraft et al., 1981) on the Black Rock basalt. However, the paleomagnetic stratigraphy is internally consistent and it is possible that the radiometric age determinations may be in error. A thick zone of reversed-polarity dominance is recorded between 53 and 658 ft, which we interpret to represent the Matuyama Chron. Several zones of normal polarity are recorded in this interval. These zones are interpreted to represent the Jaramillo, Cobb Mountain, Olduvai and Réunion subchrons, respectively. A thick zone of normal polarity from 658 to 896 ft is interpreted to represent the Gauss normal Chron. A short interval of normal polarity (626 to 635 ft) immediately above the Gauss/Matuyama boundary may represent the so-called "X-anomaly" (2.42 to 2.41 Ma). However, there is only weak magnetostratigraphic evidence for the existence of the X-anomaly (Cande and Kent, 1992). Therefore we do not correlate this zone in our magnetostratigraphic scheme. A short zone of reversed polarity near the bottom of the Black Rock core is tentatively identified as the Kaena subchron.
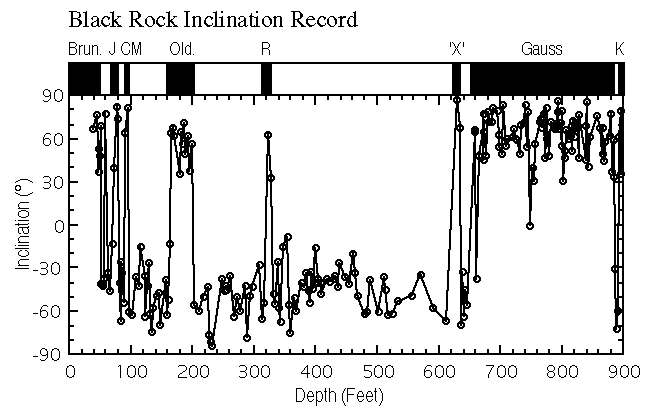 View figure 4 or
download the postscript file.
View figure 4 or
download the postscript file.Black Rock core paleomagnetic inclination record. Brun. = Brunhes, J = Jaramillo, CM = Cobb Mountain, Old. = Olduvai, R = Réunion, 'X' = Anomaly X, and K = Kaena.
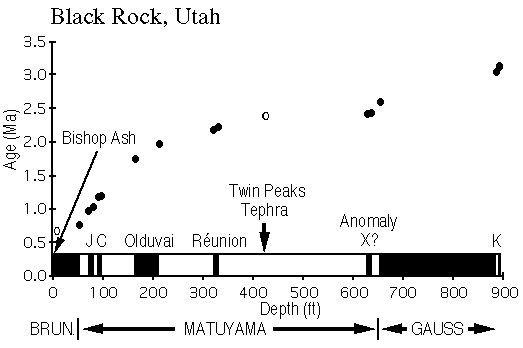 View figure 5 or
download the postscript file.
View figure 5 or
download the postscript file.Black Rock core depths versus age. Paleomagnetically-based age estimates are shown as filled circles and tephra as an open circle.J = Jaramillo, C = Cobb Mountain, K = Kaena.
Pollen data from the Black Rock core are illustrated in Figures 6 and 7, and are listed in Appendix 3. As discussed above, no core was recovered from the upper ~26 ft of sediment at basalt at the Black Rock site. Pollen samples from ~26 to ~147 ft are either barren or have extremely low pollen concentrations. The pollen assemblages recovered from this interval are probably highly altered by diagenesis and may not accurately reflect the vegetation growing at the time the sediments were deposited. Consequently, these samples are ignored in the following discussions.
The pollen flora present throughout the Black Rock record is indistinguishable from that of the modern eastern Great Basin. The extremely rare occurrences of elm (Ulmus), Fremontodendron, and a few other minor types may be the only holdovers of earlier Tertiary floras, or they may represent rebedded grains. The Black Rock pollen record overall suggests that taxa dominant today at slightly higher and cooler sites, such as pine, juniper, and sagebrush, were present near the site throughout much of the period from ~3.1 to ~1.6 Ma (Figure 6). The modern pollen rain at the site (as shown by the two uppermost samples of the profile) is dominated by Chenopodiaceae/Amaranthus, which reflects the abundance of shadscale and other Chenopodiaceae living at the site. With the exceptions of samples in the 300 - 340 ft and 415 - 430 ft core intervals, the Black Rock pollen record does not include Chenopodiaceae/Amaranthus pollen percentages approaching those of today, and we conclude that conditions were slightly wetter than today throughout most of the middle and late Pliocene and early Pleistocene. During this time, the climate was never wet enough to support abundant spruce, hemlock, or oak, and it was never apparently warm or dry enough that desert taxa dispersed into the region from the south.
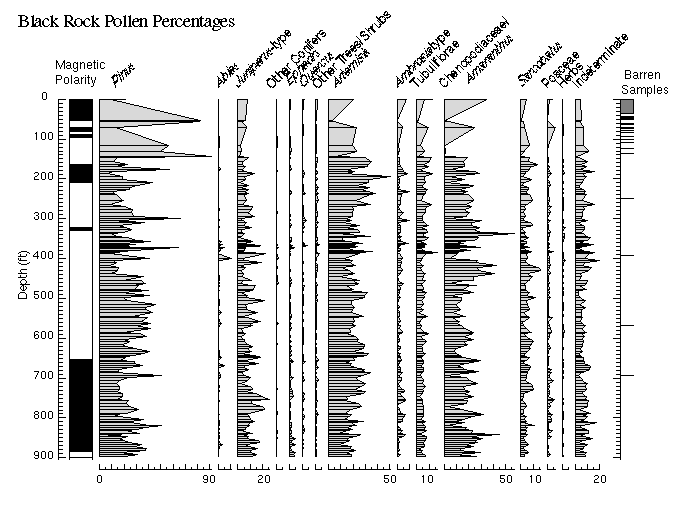 View figure 6 or
download the postscript file.
View figure 6 or
download the postscript file.The Black Rock core terrestrial pollen record plotted by depth (magnetostratigraphy shown on left). Cross-hatched area in furthest right-hand column indicates no core recovery.
To properly view the detail of this figure it is recommended that the postscript file be downloaded and printed.
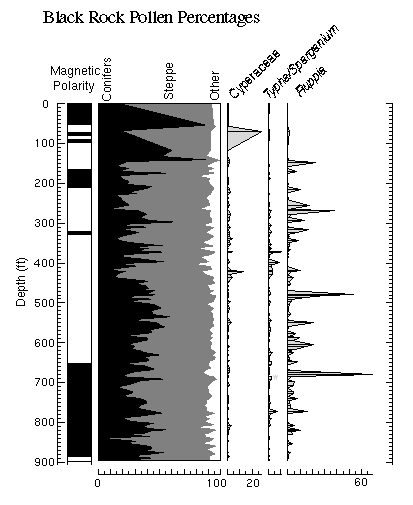 View figure 7 or
download the postscript file.
View figure 7 or
download the postscript file.Summary diagram (on left) of conifers vs. steppe plants vs. other taxa from the Black Rock core pollen record plotted against aquatic pollen types.
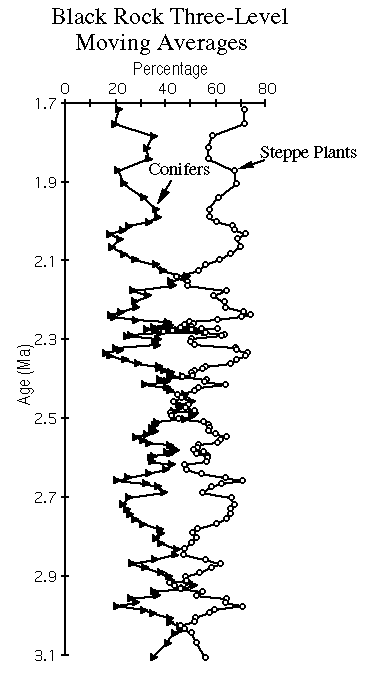 View figure 8 or
download the postscript file.
View figure 8 or
download the postscript file.Three-level moving averages of conifer and steppe pollen from the Black Rock record plotted against time.
Ostracodes and other calcareous microfossils were extracted from samples from the Black Rock core at approximately 1 ft (30 cm) intervals. Two sub-samples were processed from each sample, one for ostracode counts and curation, and the second to provide ostracode and other carbonate for stable and radiogenic isotopic analyses. Approximately 900 sample pairs have been processed thus far, and although quantitative studies have not yet been conducted on these samples, preliminary paleoenvironmental interpretations are possible from visual scans of the ostracode slides.
The ostracode assemblages from the lower portion of the Black Rock core indicate a cyclic and somewhat predictable expansion and contraction of a saline, probably shallow, lake to wetland complex. In contrast, the upper part of the core contains ostracodes that suggest irregular and unpredictable sequence of fresh to saline wetlands and shallow lakes. The older cycles have three primary components. The first involves a shallow, and typically saline, wetland to spring environment similar to the Clear Lake wetlands present today 50-km northeast of the Black Rock site. These ostracode assemblages imply waters enriched in calcium and depleted in carbonate, which in turn may indicate water/rock interactions involving sedimentary rocks. The dissolved-ion chemistry of these waters was similar to that of ocean water, and accordingly these assemblages contain estuarine as well as continental ostracodes. In other instances, the saline ostracode assemblage is represented by a single continental saline species (Limnocythere staplini), which was common in the Stansbury phase of the Bonneville lake cycle. Deep-basin ground waters and perhaps limited stream input probably supported this environment.
The second step in the environmental sequence, a step that may be absent in some sections of the core, involves the complete loss of the first assemblage and the appearance of an assemblage dominated by Elkocythereis bramlettei, an extinct ostracode that is the probable ancestor of the modern Limnocythere bradburyi. The presence of Elkocythereis bramlettei (and associated species) implies that the major dissolved ion chemistry of the lake has shifted from a calcium-enriched and carbonate-depleted environment to a calcium-depleted and carbonate-enriched environment. Waters having the latter dissolved-ion chemistry are often derived from water/rock interactions involving volcanic rocks rather than sedimentary rocks. Thus the change from the first to the second assemblage implies a change in source waters and may indicate increased stream input.
The third step in the environmental sequence involves the replacement of the Elkocythereis bramlettei assemblage with a Limnocythere ceriotuberosa assemblage, which suggests similar water chemistry but either the lake was deeper (and thus colder) or the salinity was above the tolerance range of E. bramlettei. A larger lake with colder bottom waters is the more probable explanation, but the data are insufficient to select among one of these or other possibilities.
A similar transition from the first to the third assemblage occurred during the late Pleistocene rise of Lake Bonneville from the Stansbury phase toward levels coincident with the Provo shoreline. The Bonneville sequence, however, also included a change from the Limnocythere ceriotuberosa assemblage to a specific kind of candonid assemblage. Candona remains are rare to non-existent in the samples examined thus far from the Black Rock core, which indicates that the Black Rock lake did not become fresh enough for most types of candonas to survive.
The ostracodes examined to date from the lower part of the Black Rock core imply a somewhat predictable climatic oscillation from a dry climate where lake waters are derived from local basin sources, to a wetter climate where various kinds of saline lakes are supported by streams entering the basin. The details of these transitions await completion of ostracode species counts.
The upper portion of the core contains an array of taxa that live in the region today and are often associated with wetlands located adjacent to streams. However, too few samples have been examined from this part of the core to characterize it further.
Twenty-four samples from the Black Rock core between depths of 54 and 876 ft were examined for diatoms in water mounts at a magnification of 400X. Only 6 samples were productive. Diatoms are rare to common and fairly well preserved at 236, 514, 562, 619.85, 713, and 736 ft. The diatom assemblages are characterized by distinctive species such as Surirella striatula, Campylodiscus clypeus, Navicula peregrina, N. subinflatoides, N. pygmaea, Scoliopleura peisonis, Nitzschia granulata, N. hustedtii, and Amphora coffaeiformis that indicate shallow, alkaline and chloride-rich saline water. Some levels have diatoms of coastal marine affinities such as Melosira dubia and Hyalodiscus scoticus that indicate shallow brackish water dominated by Na+ and Cl- ions. In general, the samples productive for diatoms are characterized by large percentages of Artemisia pollen, possibly indicating cool climates with some summer moisture. Barren samples were either dominated by pollen of Chenopodiaceae/Amaranthus or Pinus.
No samples examined contain large numbers of planktic, freshwater diatoms that would suggest deep, through-flowing lacustrine environments, although freshwater benthic diatoms (Epithemia turgida, Cocconeis placentula) are occasionally present that were probably introduced to the lake from marginal freshwater marsh environments associated with incoming streams.
Shallow, alkaline to saline water diatom assemblages are also found in well cuttings in the upper 300 m of drill holes in the northern part of the Great Salt Lake basin (~ 41.5° N, 112.6° W). Although correlations to the Black Rock core at similar depths are only speculative, they are consistent with a predominance of saline, shallow water environments throughout the Great Salt Lake basin in early Quaternary and Pliocene time.
Continue to Pit of Death Core Record