Open-File Report 01-415
2001
Pollen and dinocysts from marine and marginal-marine sediments of two stratigraphic test holes in Jackson County, Mississippi were studied for biostratigraphic and paleoenvironmental interpretations. In both the Ocean Springs Police Firing Range No. 1 core and the Belle Fontaine No. 1 core, the studied sediments range from Pliocene to Holocene age. Pleistocene pollen assemblages from these two cores are analogous to modern pollen assemblages from the Mississippi-Alabama Coastal Plains, and dinocyst assemblages generally are similar to modern assemblages from Mississippi Sound. Another probable Pleistocene-age sample from BelF No. 1 has a pollen assemblage similar to those of late-glacial sediments and only reworked dinocysts. Pliocene assemblages were identified by the presence of Pterocarya pollen and the dinocyst Impagidinium fenestroseptatum. Pliocene pollen assemblages in BelF No. 1 are similar to modern assemblages from the Mississippi coastal plain. Those from OSPD No. 1 have no close modern analogs from the Gulf Coast region but are similar to assemblages from interglacial deposits of Florida and North Carolina. Pliocene dinocyst assemblages from these cores are similar to those from other subtropical and tropical environments with evidence for some freshwater input.
Two stratigraphic test holes in Jackson County, Mississippi recovered fossiliferous sediments that include Pliocene, Pleistocene, and Holocene sediments that represent marine, marginal-marine, and fluvial paleoenvironments (Gohn and Reinhardt, this volume). Pleistocene and Holocene pollen assemblages from terrestrial deposits of the Atlantic Coastal Plain have been studied extensively (Delcourt and Delcourt, 1985; Watts, 1971, 1980; Whitehead, 1965), revealing patterns of vegetational change in response to glacial and interglacial intervals. The Jackson County cores provide an opportunity to determine whether these same patterns are reflected in the shallow-marine record of the central Gulf Coastal Plain.
Miocene to Holocene dinocysts from a core in the Gulf of Mexico have been studied by Wrenn and Kokinos (1986) and Holocene dinocysts from nearshore sediments of the western North Atlantic and adjacent areas have been documented by Wall (1967) and Wall and others (1977). Subtropical to tropical dinocyst assemblages from Quaternary sediments have been detailed by Rossignol (1961, 1962), Morzadec-Kerfourn (1983, 1988, 1992), Bradford and Wall (1984), Bint (1988), Turon and Londeix (1988), and McMinn (1990, 1991), among others. The Jackson County cores add to our knowledge about subtropical dinocyst populations and how they were affected by Pliocene and Pleistocene climatic fluctuations. In the following, we discuss the results of a reconnaisance study aimed at determining whether pollen and dinoflagellate cysts are sufficiently abundant in these cores for further study, at determining the age of these samples using pollen and dinocyst biostratigraphy, and at interpreting climatic and ecological changes at these sites in the Coastal Plain of Mississippi.
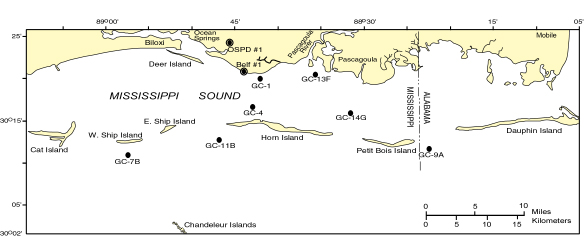
Samples for palynological analysis were collected from two cores in Jackson County, Mississippi (fig. 1). Descriptions and stratigraphic analyses of these cores are presented in Gohn and others (1992), Gohn and Reinhardt (this volume), Gohn and others (this volume), and Otvos (this volume). The core designated Ocean Springs Police Firing Range No. 1 (OSPD No. 1) (NE 1/4, NW 1/4, SW 1/4, Sec. 35, T7S, R8W, Ocean Springs 7.5 minute quadrangle) has a total depth of 170 feet. Two reconnaisance samples were collected from OSPD No. 1, one at 18.9 feet and another at 87.4 feet. The core designated Belle Fontaine No. 1 (BelF No. 1) (SW 1/4, NE 1/4, Sec. 13, T8S, R8W, Gautier South 7.5 minute quadrangle) has a total depth of 175 feet. Eight samples were collected from BelF No. 1, ranging in depth from 31.75 feet to 173.3 feet. Samples were processed using standard palynological procedures (see Edwards and Willard, this volume) and assigned USGS Paleobotanical Numbers R4331A-B (OSPD No. 1 Core) and R4224AA-AE, B, and C (BelF No. 1 core). Three hundred pollen grains and up to 600 dinoflagellates (where present) were counted from each sample.
For paleoecological analyses, pollen assemblages were compared to modern assemblages (see Edwards and Willard, this volume; Litwin and Andrle, 1992) using the modern analog technique (MAT) of dissimilarity coefficients, which compares assemblages statistically to identify the closest modern analog for each fossil assemblage. Squared chord distance (SCD) was used as the dissimilarity coefficient, and all samples with SCD = 0.15 were considered to have close modern ecological analogs (see Overpeck and others, 1985, 1992). Representative specimens of biostratigraphically or paleoecologically important taxa are illustrated on plate 1. England Finder coordinates are given for pollen illustrations, and slide coordinates for dinocyst illustrations are those of L.E. Edwards' Olympus Vanox microscope.
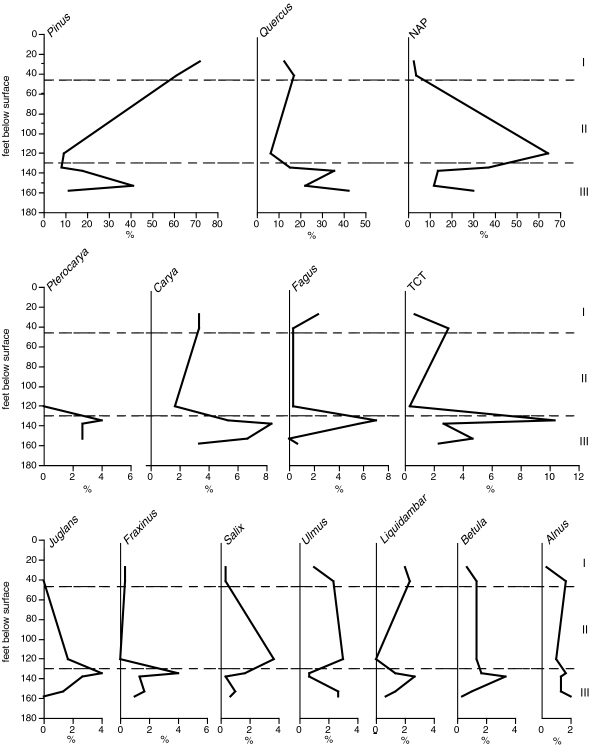
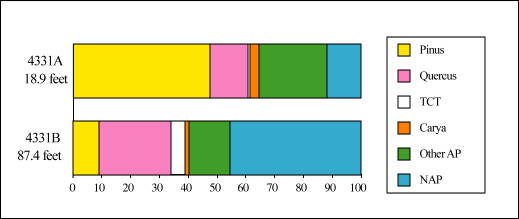
Pollen assemblages from sample R4331A (18.9 feet depth) and sample R4331B (87.42 feet depth) in OSPD No. 1 differ strikingly. Sample R4331A is from the upper part of unit 4 of Gohn and Reinhardt (this volume), and sample R4331B is from unit 2 of those authors. The higher sample is dominated by Pinus (47.5%) (pl. 1, fig. 5) and Quercus (13.4%) (pl. 1, fig. 7) pollen, with non-arboreal pollen (NAP) comprising 13.8% of the assemblage (fig. 2). Other tree pollen present include Acer, Betula, Carya (pl. 1, fig. 3), Castanea, Corylus, Fagus, Fraxinus (pl. 1, fig. 6), Liquidambar, and Ulmus (table 1), although none comprise more than 3% of the assemblage. Sample R4331B, the lower sample, is dominated by NAP (45.3%), with pollen of Quercus (25%) subdominant , and Pinus and TCT pollen (which includes members of the Taxodiaceae, Cupressaceae, and Taxaceae; pl. 1, fig. 2) comprising 9.3% and 4.7% of the assemblage, respectively. Other tree and shrub pollen commonly present include Betula, Carya, Fagus, Fraxinus, Ilex, Nyssa, and Salix, all of which individually comprise less than 2% of the assemblage (Table 1). Pterocarya (pl. 1, fig. 1), which is unknown from post-Pliocene strata in the middle Atlantic region of North America (Groot and others, 1990; Frederiksen, 1984) also is present in the assemblage from R4331B. Its presence, along with the absence of other exotic taxa typically found in Miocene sediments of the Coastal Plain (for example, Cyrilla, Engelhardia, Gordonia, Podocarpus, Symplocos (Frederiksen, 1984; Groot and others, 1990)), suggests a Pliocene age for this sample.
| Core: | . | OSPD | OSPD | BelF | BelF | BelF | BelF | BelF | BelF | BelF |
|---|---|---|---|---|---|---|---|---|---|---|
| R Number: | . | 4331A | 4331B | 4424AA | 4424AB | 4424AC | 4424B | 4424C | 4424AD | 4424AE |
| Depth (feet): | . | 18.9 | 87.4 | 31.75 | 46.4 | 132.3 | 148 | 152 | 168.25 | 173.3 |
| Arboreal Pollen | . | . | . | . | . | . | . | . | . | . |
| Scientific Name | Common Name | . | . | . | . | . | . | . | . | . |
| Acer | Maple | 0.98 | 0 | 0 | P | P | 0.33 | 0.67 | 0 | 0 |
| Betula | Birch | 2.95 | 1.33 | 0.67 | 1.33 | 1.33 | 1.67 | 3.33 | 1 | 0.33 |
| Carya | Hickory | 2.95 | 1.33 | 3.33 | 3.33 | 1.67 | 5.33 | 8.33 | 6.67 | 3.33 |
| Castanea | Chestnut | 2.95 | 0 | 0 | 0 | 0.67 | 0 | 0 | 0 | 0.33 |
| Corylus | Hazlenut | 1.31 | P | 0.67 | 1 | 1.67 | P | 0.67 | P | P |
| Fagus | Beech | 1.31 | 1.33 | 2.33 | 0.33 | 0.33 | 7 | 5.67 | 0 | 0.67 |
| Fraxinus | Ash | 1.31 | 0.67 | 0.33 | 0.33 | P | 4 | 1.33 | 1.67 | 1 |
| Juglans | Walnut | 0 | P | P | 0 | 1.67 | 0 | 0 | 1.33 | P |
| Liquidambar | Sweetgum | 0.98 | 0.33 | 2 | 2.33 | P | 1.33 | 2.67 | 1.33 | 0.67 |
| Myrica | Bayberry | 0 | 0 | 0.33 | 0 | 0 | 0 | 0 | 0 | P |
| Nyssa | Watergum | 3.21 | 2 | 0.33 | 1 | 0 | 0 | 0 | 0 | P |
| O/C* | Hophornbeam | 0.64 | 0 | 0.33 | 0.33 | 0.33 | 0.67 | 0 | 0 | 0 |
| Palmae | Palm | 0 | 0 | 0 | 0 | 0.33 | 0 | 0.67 | 0 | 0.33 |
| Picea | Spruce | 0 | 0 | 0 | 0 | 0.33 | P | 0 | P | 0 |
| Pinus | Pine | 47.54 | 9.33 | 72 | 61.33 | 9.33 | 8.33 | 18 | 41.33 | 11.67 |
| Pterocarya | Wingnut | 0 | 0.67 | 0 | 0 | 0 | 4 | 2.67 | 2.67 | 1.33 |
| Quercus | Oak | 13.44 | 25 | 12.67 | 17 | 6.33 | 15.33 | 35.67 | 22.33 | 42.33 |
| Salix | Willow | 0.64 | 1.33 | 0.33 | 0.33 | 3.67 | 1.67 | 0.33 | 1 | 0.67 |
| TCT** | 0.64 | 4.67 | 0.67 | 3 | 0.33 | 10.33 | 2.67 | 4.67 | 2.33 | |
| Tilia | Basswood | 0 | P | P | 0 | 0 | P | 0.33 | P | 0 |
| Ulmus | Elm | 1.31 | 0.33 | 1 | 2.33 | 3 | 0.67 | 0.67 | 2.67 | 2.67 |
| . | . | . | . | . | . | . | . | . | . | . |
| * Ostrya/Carpinus | . | . | . | . | . | . | . | . | . | . |
| ** Taxodiaceae, Cupressaceae, Taxaceae | . | . | . | . | . | . | . | . | . | . |
| . | . | . | . | . | . | . | . | . | . | . |
| Non-Arboreal Pollen | . | . | . | . | . | . | . | . | . | . |
| Scientific Name | Common Name | . | . | . | . | . | . | . | . | . |
| Alnus | Alder | 2.88 | 0.67 | 0.33 | 1.67 | 1.00 | 1.67 | 1.33 | 1.33 | 2.00 |
| Ambrosia | Ragweed | 1.60 | 9.33 | P | 0 | 9.67 | 6.67 | 0.33 | 0.67 | 4.33 |
| Artemesia | Sage | 0.98 | 1.33 | P | 0 | 15.67 | 5.00 | 3.33 | 1.67 | 2.33 |
| Aster | Aster | 0.98 | 3.00 | 0.67 | 0.67 | 14.67 | 4.00 | 1.00 | 2.00 | 2.33 |
| Asteraceae indet. | Aster | 0.32 | 0.33 | 0 | 0 | 0.33 | 0.33 | 0 | 0 | 0.33 |
| Caryophyllaceae | Pink family | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | P |
| Chenopods | Pigweed family | 0.98 | 2.67 | 0.33 | 0.33 | 9.67 | 0.67 | 0.33 | 0.33 | 1.00 |
| Cyperaceae | Sedge family | 0.98 | 5.67 | 0.33 | 0.67 | 2.00 | 9.67 | 0.67 | 3.67 | 2.33 |
| Ephedra | Ephedra | 0 | P | 0 | 0 | 0.33 | 0 | 0 | 0 | 0 |
| Ericaceae | Heath family | P | 0 | P | P | 0.67 | 0.67 | P | 0.33 | P |
| Ilex | Holly | 2.24 | P | 0 | 0.33 | P | 0.67 | 0.67 | 0.33 | 0 |
| Leguminosae | Legume family | 0.64 | P | P | 0.33 | 0.67 | P | 1.00 | P | P |
| Liguliflorae | Legume family | 0 | P | 0 | 0 | 0.33 | P | 0 | P | P |
| Poaceae | Grass family | 3.53 | 27.67 | 1.33 | 2.00 | 11.67 | 9.33 | 7.67 | 3.00 | 17.33 |
| Ranunculaceae | Buttercup family | 0 | 0 | 0 | P | 0.33 | 0 | 0 | 0 | 0 |
| Umbelliferae | Parsley family | 1.92 | 1.00 | P | P | 2.00 | 0.67 | P | P | 0.33 |
Comparison of assemblages from OSPD No. 1 with modern assemblages shows that sample R4331A has close modern analogs (SCD = 0.15) in the modern dataset from sites located off the Mississippi coast between Biloxi Bay and Pascagoula Bay (sites GC-1, GC-13F; fig. 1). Sample R4331B has no close modern analogs, but the most similar modern assemblage (SCD=0.45) is sample GC-13F from near the mouth of the Pascagoula River (fig. 1), which has relatively low percentages of Pinus and higher percentages of Quercus and NAP. Other samples from the same area have slightly higher SCD values, as do two samples collected from the New Jersey and Massachusetts coasts, which reflect pollen assemblages representative of the hickory-oak-pine forests that dominate those areas (Küchler 1985). However, these modern assemblages all have much lower abundances of NAP than R4331B.
Both samples from OSPD No. 1 contain very sparse dinocyst assemblages. Sample R4331A (18.9 feet depth) contains the subtropical to tropical, nearshore forms Polysphaeridium zoharyi (pl. 1, fig. 18) and Lingulodinum machaerophorum, (pl. 1, fig. 17) and the freshwater alga Pediastrum (table 2). In addition, the reworked Tertiary forms Homotryblium vallum (Neogene?), Muratodinium fimbriatum, and Wetzeliella sp. (Paleogene) are present. In contrast, sample R4331B (87.42 feet depth) is dominated by Spiniferites spp. (pl. 1, figs. 19,20) and Operculodinum spp. (pl. 1, fig. 16). It also contains several specimens of the dinocyst Impagidinium fenestroseptatum (pl. 1, figs. 13,14). The only previous report of this form is from the early Pliocene calcareous nannofossil zone NN12 of the Labrador Sea (Head and others, 1989). The genus Impagidinium generally is considered to be indicative of offshore environments. The freshwater alga, Pediastrum, also is present (table 2).
| Core: | OSPD | OSPD | BelF | BelF | BelF | BelF | BelF | BelF | BelF |
|---|---|---|---|---|---|---|---|---|---|
| R Number: | 4331A | 4331B | 4424AA | 4424AB | 4424AC | 4424B | 4424C | 4424AD | 4424AE |
| Depth (feet): | 18.9 | 87.4 | 31.75 | 46.4 | 132.3 | 148 | 152 | 168.25 | 173.3 |
| . | . | . | . | . | . | . | . | . | . |
| Achomosphaera sp. | . | . | . | . | . | . | . | 10.23 | . |
| Impagidinium fenestroseptatum | . | 10.14 | . | . | . | . | . | 2.27 | . |
| Lingulodinium machaerophorum | P | 2.90 | 15.83 | 34.33 | . | . | . | 1.14 | . |
| Multispinula quanta | . | . | . | . | . | . | . | 1.14 | . |
| Nematosphaeropsis labyrinthus | . | . | 1.33 | 1.00 | . | . | . | 1.14 | . |
| Operculodinium centrocarpum s.l. | . | 0.67 | 2.00 | . | . | . | . | 1.14 | . |
| Operculodinium israelianum | . | 1.45 | 0.33 | 1.00 | . | . | . | 13.64 | . |
| Operculodinium spp. | P | 28.99 | . | . | . | . | . | . | . |
| Polysphaeridium zoharyi | P | 1.45 | 70.00 | 48.00 | . | . | . | . | . |
| Selenopemphix sp. | . | . | . | . | . | . | . | 1.14 | . |
| Spiniferites spp. | . | 42.03* | 10.00 | 9.33 | . | . | . | 56.82 | . |
| Trinovantedinium sp. | . | . | . | . | . | . | . | 1.14 | . |
| Tuberculodinium vancampoae | P | 11.59 | 1.33 | 1.67 | . | . | . | 9.10 | . |
| Indeterminate dinocysts | . | 1.45 | 0.33 | 2.67 | . | . | . | 1.14 | . |
| Reworked older dinocysts | P | . | P | . | P | P | P | . | . |
| Nondiagnostic dinocysts | . | . | . | . | . | . | . | . | 100.00 |
| . | . | . | . | . | . | . | . | . | |
| Total specimens counted: | . | 69 | 600 | 300 | . | . | . | 88 | 3 |
| * includes S. mirabilis 7.24% | . | . | . | . | . | . | . | . | . |
Seven samples from BelF No. 1 range in depth from 31.7-173.3 feet, but the sample at 173 feet is barren of pollen and is not discussed. The compositions of the pollen assemblages vary down the core (fig. 3), and three pollen associations are recognizable. Association I, which consists of samples at 31.75 feet and 46.4 feet, represents unit GB of Gohn and Reinhardt (this volume) and the Biloxi Formation of Otvos (this volume). This association is dominated strongly by Pinus pollen (61-72%), with Quercus subdominant (12.7-17%) (Fig. 3). Betula, Carya, Liquidambar, TCT, and Ulmus pollen are commonly present, and NAP is relatively rare (<5%) (table 1). Close modern analogs for pollen association I are found in many samples collected off the coasts of Mississippi and Alabama (including all offshore sites illustrated in fig. 1) and from the east coast of the U.S. ranging from central North Carolina south to Key West.
Pollen Association II is found in only one sample, from 132.3 feet depth, in unit 3 of Gohn and Reinhardt (this volume). It differs strongly from all others in the strong dominance of NAP (64.7%) (fig. 3). Pinus and Quercus are less common than in the other samples, comprising 9.3% and 6.3% of the assemblage, respectively. Other tree and shrub pollen present in the count include Betula, Carya, Corylus, Ilex (pl. 1, fig. 8), Juglans, Picea, Salix, and Ulmus (table 1). The NAP component is dominated by members of the Asteraceae (40.3%), with chenopod and grass pollen also common (9.7% and 11.7%, respectively) (pl. 1, figs. 9-12). This association has no close modern analog in the existing dataset, but the most similar assemblages in the modern database (SCD=0.24-0.25) are from the coasts of North Carolina, Maryland, and New Jersey.
Pollen Association III includes three samples from 148 feet to 168.25 feet depth that represent unit 2 of Gohn and Reinhardt (this volume). This association is differentiated by lower abundances of NAP than in the other associations, highest abundance of Quercus, and presence of Pterocarya. These samples are dominated by Quercus (two samples) or Pinus (one sample) (fig. 3). Tree pollen present in the counts includes Betula, Carya, Fagus, Fraxinus, Liquidambar, Pterocarya, Salix, Tilia (pl. 1, fig. 4), and Ulmus (table 1). Most of the NAP are Asteraceae, Cyperaceae, and Poaceae, with relatively small contributions by the chenopods. A Pliocene age is suggested for this association by the presence of Pterocarya. Only one sample in Association III (R4424AD) has close modern analogs, and these are from offshore sites near West Ship Island (Sites GC-4 and GC-7; Fig. 1). The most similar modern assemblages to the other two samples (SCD=0.27-0.64) are from site GC-7B (fig. 1) and from off the New Jersey coast, where oak-pine forests dominate the vegetation (Greller ,1988; Küchler, 1985).
Of the eight samples studied from BelF No. 1, only two contained well preserved dinofloras. Four others contained only Pediastrum and dinocysts that obviously were reworked from older sediments, and two contained sparse dinocysts. The two highest samples (31.75 feet and 46.4 feet depth; unit GB of Gohn and Reinhardt, this volume) contain well-preserved dinocyst floras dominated by Polysphaeridium zoharyi. In both samples, Spiniferites spp., Tuberculodinium vancampoae, Nematosphaeropsis labyrinthus (pl. 1, fig. 15) and Operculodinium sp. are minor components, and Pediastrum (freshwater alga) is present. The lower of these samples also contains a significant proportion of Lingulodinium machaerophorum. The three samples ranging from 132.3 feet to 152 feet depth (units 2 and 3 of Gohn and Reinhardt, this volume) are presumably nonmarine or estuarine. They contain poorly preserved dinocysts and algae, including Pediastrum and reworked Cretaceous and Tertiary dinocysts. The sample at 168.25 feet depth is marine and contains a sparse dinocyst flora, dominated by Spiniferites spp. Impagidinium fenestroseptatum is a rare but conspicuous component, and the freshwater alga, Pediastrum, is present. The age is definitely pre-Holocene and most likely is Pliocene, based on the presence of Impagidinium fenestroseptatum. The environment of deposition is similar to that in the lower sample from the OSPD No. 1 core. The lowest sample studied from the BelF No. 1 core (R4424AE, 173.3 feet depth) contains only three nondiagnostic dinocysts.
Based on palynological evidence, both cores extend into Pliocene sediments and record changes in both terrestrial plant communities and dinocyst assemblages that correspond to climatic changes and varying proximity to paleo-shorelines. In OSPD No. 1, both the pollen and dinocyst assemblages from the upper sample are analogous to modern assemblages from the same area. Presently, pine flatwoods are the dominant vegetation type throughout much of the Mississippi Coastal Plain (Christensen, 1988), and the sparse dinocyst assemblage appears to represent an environment similar to nearshore areas of the present Mississippi Sound.
The lower sample from OSPD No. 1 (R4331B) apparently is Pliocene in age, based on the presence of Pterocarya pollen and the dinocyst Impagidinium fenestroseptatum. This pollen assemblage, dominated by non-arboreal pollen, has no close modern analogs, and the most similar assemblages are those from organic muds deposited during the Sangamon interglacial and modern postglacial times in the Coastal Plain. Sangamon pollen assemblages from North Carolina and Florida exhibit intervals dominated by NAP (Ambrosia, Gramineae, other herb pollen) with abundant Quercus pollen and smaller proportions of Pinus, Carya, and other mesic trees (Frey, 1953; Watts, 1975, 1980). These are interpreted as representing mixed scrub oak/prairie vegetation that would require relatively warm, dry conditions and that have no modern analogs (Watts and Stuiver, 1980; Watts, 1980). This assemblage differs strikingly from Pliocene assemblages from southwestern Florida, which are dominated consistently by Pinus with subdominant Quercus throughout much of the Pliocene (Willard and others, in press). The presence of plant communities dominated by NAP in the Pliocene may indicate that these sediments are younger than about 2.5 Ma, when the amplitude and frequency of climatic fluctuations changed and Northern Hemipshere ice growth began (see Raymo and others, 1990). The dinocysts also show a very different assemblage from the upper sample and are dominated by Spiniferites spp. and Operculodinum spp. The species Impagidinium fenestroseptatum indicates a more offshore environment than the upper sample.
The sample set studied from BelF No. 1 provides more complete coverage of that core than is available for the OSPD No. 1 core and includes samples ranging in age from Pliocene to Pleistocene. As with OSPD No. 1, the Pliocene age assignment of the samples ranging from 148 feet to 170 feet depth is based on the presence of Pterocarya and Impagidinium fenestroseptatum. No age estimates are made for the lowermost sample (173.3 feet depth), which is barren of pollen and has only nondiagnostic dinocyst species.
Pollen Association I occurs in late Pleistocene, nearshore-marine sediments with abundant calcareous macro- and microfossils (see Cronin, this volume; Poag, this volume; Wingard, this volume). This association is representative of the long-leaf/slash pine communities that presently dominate much of the coastal plain from southern Florida to North Carolina. The dinocyst assemblages from this interval are well preserved and may indicate two somewhat different environments, both dominated by P. zoharyi but with the lower sample including significantly higher numbers of L. machaerophorum . The overall dinocyst assemblages in this interval suggest tropical to subtropical, very nearshore environments. Both samples contain higher percentages of L. machaerophorum (16-34%) than any of our modern samples from the Mississippi Sound and Gulf of Mexico (Edwards and Willard, this volume; maximum percentage is 5.2). In the Atlantic Ocean, Wall and others (1977) found only a few samples that contained more than 10 percent L. machaerophorum , and only three of these contained more than 20 percent P. zoharyi. These samples are all from very shallow, protected "sounds" in Bermuda, the Bahamas, and Puerto Rico.
Pollen Association II, with the dominance of NAP, low abundances of Quercus and Pinus pollen, and presence of Picea pollen, is most similar to assemblages deposited during late-glacial intervals. Picea has been identified from late Wisconsinan deposits of east-central Louisiana (Delcourt and Delcourt 1977), and the pollen assemblage from Association II in BelF No. 1 also is similar in the dominance of NAP and abundance of Quercus and mesic taxa. Therefore, pollen Association II probably also represents a late glacial deposit, but further study and dating of the sediments are required to assign the proper glacial stage. Only reworked dinocysts were observed in this sample, and, as has been suggested by other workers (Wrenn and Kokinos, 1986; de Vernal and others, 1987), an increase in the numbers of reworked dinocysts may correlate to glacial intervals.
Pollen Association III is Pliocene in age, based on the presence of Pterocarya pollen and the dinocyst Impagidinium fenestroseptatum. Only the sample from 168.25 feet depth has a close modern analog, from near West Ship Island (GC7B; fig. 1), but this site also is the closest analog for the other two samples in pollen Association III. Although the presence of one modern analog indicates that Pliocene conditions may have been similar to those in Mississippi today, the existence of higher than present percentages of Quercus and the presence of Picea in the fossil assemblages suggests that these sediments may have been deposited during a relatively cool part of the Pliocene. The sparse dinoflora is similar to that found in the lower sample in the Ocean Springs core (87.4 feet), and the depositional environment probably had some freshwater influence.
Pollen and dinocyst assemblages from two cores in Jackson County, Mississippi provide a partial record of vegetational and paleoceanographic changes from the Pliocene to the late Pleistocene. Palynomorphs from late Pleistocene samples indicate environments and climatic conditions similar to today. The pollen assemblage composition and the presence of only reworked dinocysts in sediments from 130 feet depth in BelF No. 1 suggest that these sediments may have been deposited during late-glacial conditions of the Pleistocene. Pliocene sediments also are present in both cores, based on the presence of Pterocarya pollen and the dinocyst Impagidinium fenestroseptatum. Pollen assemblages from Pliocene samples in both OSPD No. 1 and BelF No. 1 represent a different flora than presently found in the region and suggest cooler conditions than today. Dinocysts from these Pliocene samples support deposition in a more offshore environment than is suggested for the stratigraphically higher samples. The stratigraphic range encompassed by both cores and the relatively high abundance of palynomorphs, particularly pollen, makes this core ideal for further, higher resolution study. Further research should provide more detailed evidence of vegetational changes in response to glacial/interglacial periods in the southeastern United States and allow assessment of the response of subtropical dinocysts to glacial cycles.
Acknowledgements. We thank N. J. Durika (USGS) for processing the samples and V.A.S. Andrle, T. Servais, and D.B. Weaver (all USGS) for rendering the illustrations. We also thank R.J. Litwin (USGS) and G.S. Gohn (USGS) for reviews and discussions of the manuscript.
Bint, A.D., 1988, Recent dinoflagellate cysts from Mermaid Sound, northwestern Australia: Memoir of the Association of Australasian Palaeontologists, v. 5, p. 329-341.
Bradford, M.R., and Wall, D.A., 1984, The distribution of Recent organic-walled dinoflagellate cysts in the Persian Gulf, Gulf of Oman, and northwestern Arabian Sea: Palaeontographica: Abt. B., v. 192, p. 1-84.
Christensen, N.L., 1988, Vegetation of the southeastern coastal plain, in Barbour, M.G.and Billings, W.D., ed., North American Terrestrial Vegetation, p. 317-364.
Delcourt, P.A. and Delcourt, H.R., 1977, The Tunica Hills, Louisiana-Mississippi: Late Glacial locality for spruce and deciduous forest species: Quaternary Research, v. 7, p. 218-237.
Delcourt, H.R. and Delcourt, P.A., 1985, Quaternary palynology and vegetational history of the southeastern United States, in Bryant, V.M., Jr. and Holloway, R.G., ed., Pollen records of late-Quaternary North American sediments, p. 1-37.
de Vernal, A., Hillaire-Marcel, C. , Aksu, A.E., and Mudie, P.J., 1987. Palynostratigraphy of Baffin Bay deep sea cores: climatostratigraphic implications: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 61, P. 97-105.
Frederiksen, N.O., 1984, Stratigraphic, paleoclimatic, and paleobiogeographic significance of Tertiary sporomorphs from Massachusetts: U. S. Geological Survey Professional Paper, v. 1308, p. 1-25.
Frey, D.G., 1953, Regional aspects of the late-glacial and post-glacial pollen succession of southeastern North Carolina: Ecological Monographs, v. 23, no. 3, p. 289-313.
Gohn, G. S., Reinhardt, J. and Garrison, J.A., Jr., 1992, Preliminary lithologic logs for three stratigraphic test holes in Jackson County, Mississippi: U.S. Geological Survey Open-File Report 92-394, 23 p.
Greller, A.M., 1988, Deciduous Forest, in Barbour, M.G. and Billings, W.D., ed., North American Terrestrial Vegetation, p. 287-316.
Groot, J.J., Ramsey, K.W. and Wehmiller, J.F., 1990, Ages of the Bethany, Beaverdam, and Omar Formations of southern Delaware: Delaware Geological Survey Report of Investigations, v. 47, p. 1-19.
Küchler, A.W., 1985, Potential Natural Vegetation-National Atlas of the United States of America: U. S. Geological Survey, 2nd.
Litwin, R.J. and Andrle, V.A.S., 1992, Modern palynomorph and weather census data from the U. S. Atlantic coast (Continental Margin Program samples and selected NOAA weather stations): U. S. Geological Survey Open-File Report, v. 92-263, p. 1-31.
McMinn, A., 1990, Recent dinoflagellate cyst distribution in eastern Australia. Review of Palaeobotany and Palynology, v. 65, p. 305-310.
McMinn, A., 1991, Recent dinoflagellate cysts from estuaries on the central coast of New South Wales, Australia. Micropaleontology, v. 37, no. 3, p. 269-287.
Morzadec-Kerfourn, M.-T., 1983, Intérêt de kystes de dinoflagellés pour l'établissement de reconstitution paléogéographique: exemple du Golfe de Gabès: Cahiers de Micropaléontologie, v. 4, p. 15-22.
Morzadec-Kerfourn, M.-T., 1988, Paléoclimats et paléoenvironnement, du Tardiglaciaire au Récent, en Méditerranée orientale, à l'est du delta du Nil; l'apport des microfossiles à membrane organique: Bulletin des centres de Recherches Exploration-Production Elf-Aquitaine, v. 12, no. 1, p. 3267-275.
Morzadec-Kerfourn, M.-T., 1992, Upper Pleistocene and Holocene dinoflagellate cyst assemblages in marine environments of the Mediterranean Sea and the northwest Atlantic coast of France, in Head, J.J., and Wrenn, J.H., eds., Neogene and Quaternary Dinoflagellate Cysts and Acritarchs: American Association of Stratigraphic Palynologists Foundation, Salt Lake City, p. 121-132.
Overpeck, J.T., Webb, R.S., and Webb, T. III, 1992, Mapping eastern North American vegetation change of the past 18 ka: No-analogs and the future: Geology, v. 20, p. 1071-1074.
Overpeck, J.T., Webb, T., III, and Prentice, I.C., 1985, Quantitative interpretation of fossil pollen spectra: dissimilarity coefficients and the method of modern analogs: Quaternary Research, v. 23, p. 87-108.
Raymo, M. E., Rind, D., and Ruddiman, W.F., 1990, Climatic effects of reduced Arctic sea ice limits in the GISS II General Circulation Model: Paleoceanography, v. 5, p. 367-382.
Rossignol, M., 1961, Analyse pollinique de sédiments marins quaternaires en Israël. I: Sédiments récents: Pollen et Spores, v. 3 no. 2, p. 303-324.
Rossignol, M., 1962, Analyse pollinique de sédiments marins quaternaires en Israël. I: Sédiments Pleistocénes: Pollen et Spores, v. 4 no. 1, p. 121-148.
Turon, J.-L., and Londeix, L., 1988, Les assemblages de kystes de dinoflagellés en Méditerranée occidentale (mer d'Alboran): Mise en évidence de l'évolution des paléoenvironnements depuis le dernier maximum glaciaire: Bulletin des centres de Recherches Exploration-Production Elf-Aquitaine, v. 12, no. 1, p. 313-344.
Wall, D., 1967, Fossil microplankton in deep-sea cores from the Caribbean Sea: Palaeontology, v. 10, no. 1, p. 95-123.
Wall, D., Dale, B., Lohmann, G.P., and Smith, W.K., 1977, The environmental and climatic distribution of dinoflagellate cysts in modern marine sediments from regions in the North and South Atlantic Oceans and adjacent seas: Marine Micropaleontology, v. 2, p. 121-200.
Watts, W.A., 1971, Postglacial and interglacial vegetation history of southern Georgia and central Florida: Ecology, v. 52, no. 4, p. 676-690.
Watts, W.A., 1975, A late Quaternary record of vegetation from Lake Annie, south-central Florida: Geology, v. 3, p. 344-346.
Watts, W.A., 1980, The Late Quaternary vegetation history of the southeastern United States: Annual Review of Ecology and Systematics, v. 11, p. 387-409.
Watts, W.A. and Stuiver, M., 1980, Late Wisconsin climate of northern Florida and the origin of species-rich deciduous forest: Science, v. 210, p. 325-327.
Whitehead, D.R., 1965, Palynology and Pleistocene phytogeography of unglaciated eastern North America, in Wright, H.E., Jr., and Frey, D.G., ed., The Quaternary of the United States, p. 417-432.
Willard, D.A., Cronin, T.M., Ishman, S.E. and Litwin, R.J., 1993, Terrestrial and marine records of climatic and environmental changes during the Pliocene in subtropical Florida: Geology, v. 21, p. 689-692.
Wrenn, J. H., and Kokinos, J. P., 1986. Preliminary comments on Miocene through Pleistocene dinoflagellate cysts from De Soto Canyon, Gulf of Mexico: in Wrenn, J.H., Duffield, S.L., and Stein, J. A., eds., Papers from the first symposium on Neogene dinoflagellate cyst biostratigraphy: American Association of Stratigraphic Palynologists Contributions Series Number 17, p. 169-225, 20 pl.
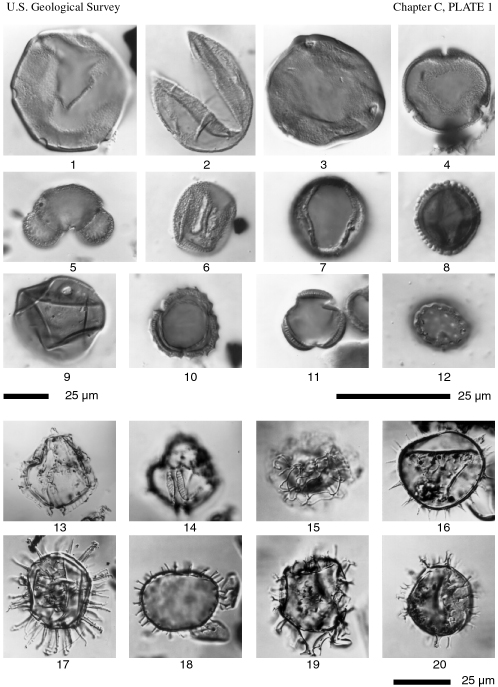
Figure 1. Pterocarya pollen, sample R4424B (1). BelF No. 1 at 148 feet. England Finder coordinates D42/4.
Figure 2. TCT pollen (Taxodiaceae, Cupressaceae, Taxaceae), sample R4424B (1). BelF No. 1 at 148 feet. England Finder coordinates E38/2.
Figure 3. Carya pollen, sample R4424B (1). BelF No. 1 at 148 feet. England Finder coordinates N42/3.
Figure 4. Tilia pollen, sample R4424AA (1). BelF No. 1 at 31.75 feet. England Finder coordinates F31/3.
Figure 5. Pinus pollen, sample R4424AA (1). BelF No. 1 at 31.75 feet. England Finder coordinates K27/4.
Figure 6. Fraxinus pollen, sample R4424AD (1). BelF No. 1 at 168.25 ft. England Finder coordinates D41/4.
Figure 7. Quercus pollen, sample R4424AA (1). BelF No. 1 at 31.75 feet. England Finder coordinates J27/1.
Figure 8. Ilex pollen, sampleR4424AD (1). BelF No. 1 at 168.25 ft. England Finder coordinates X14/0.
Figure 9. Poaceae pollen, sample R4424AD (1). BelF No. 1 at 168.25 ft. England Finder coordinates F39/0.
Figure 10. Ambrosia pollen, sample R4424B (1). BelF No. 1 at 148 feet. England Finder coordinates F29/4.
Figure 11. Artemesia pollen, sample R4424B (1). BelF No. 1 at 148 feet. England Finder coordinates G34/2.
Figure 12. Chenopodiaceae/Amaranthaceae pollen, sample R4424AD (1). BelF No. 1 at 168.25 feet. England Finder coordinates F41/1.
Figures 13, 14. Impagidinium fenestroseptatum Head et al. Ventral views of optical section and dorsal surface, sample R4431B (2). OSPD No. 1 at 87.42 feet. Edwards slide coordinates, 27.1x96.4.
Figure 15. Nematosphaeropsis labyrinthus (Ostenfeld) Reid. Orientation uncertain, high focus, sample R4424AA (4). BelF No. 1 at 31.75 feet. Edwards slide coordinates, 20.9x88.2.
Figure 16. Operculodinium israelianum (Rossignol) Wall. Ventral view of optical section, sample R4431B (2). OSPD No. 1 at 87.42 feet. Edwards slide coordinates, 36.8x85.5
.Figure 17. Lingulodinium machaerophorum (Deflandre & Cookson) Wall. Apical view of optical section, sample R4424AA (4). BelF No. 1 at 31.75 feet. Edwards slide coordinates, 35.1x103.8.
Figure 18. Polysphaeridium zoharyi (Rossignol) Bujak et al. Antapical view of optical section, sample R4424AA (4). BelF No. 1 at 31.75 feet. Edwards slide coordinates, 20.9x88.2.
Figure 19. Spiniferites mirabilis (Rossignol) Sarjeant. Oblique dorsal view of optical section, sample R4431B (2). OSPD No. 1 at 87.42 feet. Edwards slide coordinates, 22.0x80.5.
Figure 20. Spiniferites sp. B of Wrenn and Kokinos (1986). Ventral view of optical section, sample R4431B (2), OSPD No. 1 at 87.42 feet. Edwards slide coordinates, 36.4x75.0.

![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: http://pubsdata.usgs.gov/pubs/of/2001/of01-415/chap3txt.htm
Page Contact Information: USGS Publications Team
Page Last Modified: Wednesday, 07-Dec-2016 18:56:35 EST