Open-File Report 01-415
Surface sediments from the shallow waters of Mississippi Sound and the Gulf of Mexico show patterns in the distributions of marine and nonmarine palynomorphs that can be related to the marine and terrestrial ecology and to transport history. Polysphaeridium zoharyi (Rossignol) Lentin & Williams dominates the dinocyst assemblages in all but the most offshore sediment samples and shows an inverse relationship with increasing water depth. This species shows relatively less dominance in samples near open passages between barrier islands than in samples from areas that are relatively sheltered behind islands. In the very shallow-water samples in this study, cysts of the family Congruentidiaceae show a direct relationship with water depth. Nearly all the cysts recovered in this study, regardless of taxonomic affinity, are round and spiny: Polysphaeridium, Multispinula, Spiniferites, Lingulodinium, Nematosphaeropsis, Operculodinium. Specimens within a taxon show wide variation in size, surface features, and spine length. The pollen assemblages represent the shallow marine record of the southern mixed forest, which consists primarily of pine-dominated forests with abundant oak and a variety of hardwoods and shrubs; such forests cover most of Florida, southern Mississippi and Alabama, and southeastern Georgia. The assemblages from these samples are very similar to each other, with the only notable variation the abundance of Pinus and Quercus pollen. Pinus pollen is most abundant in assemblages from deep-water sediments and Quercus shows no coherent pattern in the relatively small area studied.
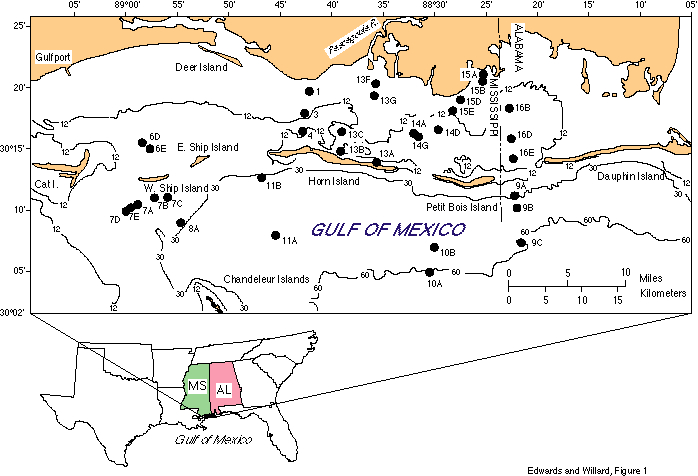
The study area (fig. 1) includes the coastal areas offshore of eastern Mississippi and westernmost Alabama. The area is adjacent to Biloxi Bay, the Pascagoula River, Mobile Bay, and numerous bayous. Samples were taken between the coast and the barrier islands (Mississippi Sound) and seaward of the barrier islands (Gulf of Mexico) and represent modern sedimentation. Our study of the dinoflagellate cysts and pollen from surface sediments in these shallow waters will help understand the distributions of these palynomorphs in older marine sediments preserved in the subsurface of coastal Mississippi.
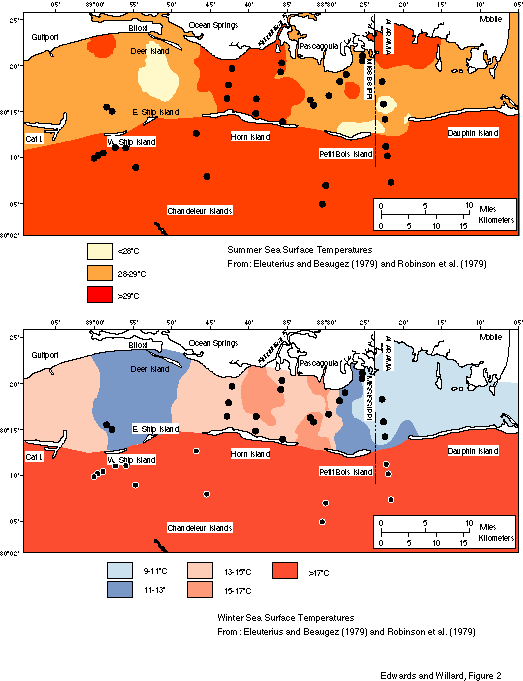
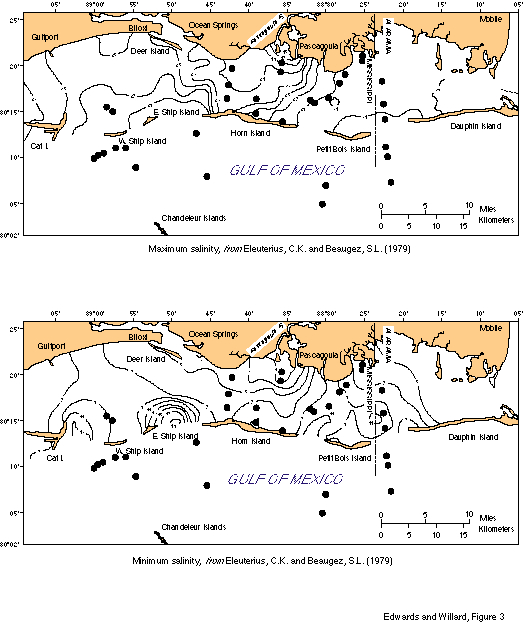
Climate in the study area is generally tropical to subtropical. Summer atmospheric temperatures average 28oC and winter atmospheric temperatures are 10-11oC (National Climatic Center, 1983; National Oceanographic and Atmospheric Administration, 1985). Summer sea-surface temperatures average 30oC; winter sea-surface temperatures are 12-15oC (fig. 2). Surface salinity is highly variable, both landward and seaward of the barrier islands (fig. 3). Landward, it varies from 1 o/oo in the spring to 30 o/oo in the fall. Even 25 km offshore, direct measurements of surface salinity in September were less than 30 o/oo. The bayous, rivers, and bays draining the adjacent land area bring in considerable sediment and freshwater. Hurricanes and tropical storms are a major influence in the maintenance of the barrier islands. Historic records (Eleuterius and Beaugez, 1979) indicate one hurricane strikes the area every four years, one tropical storm every three.
Studies of the distribution of dinocysts in modern marine sediments began with observations of the "hystrichospheres" in pollen studies. Muller (1959) noted patterns of abundance and morphological appearance of "Hystrix" in estuarine to shelf areas near the Orinoco delta off the coast of Venezuela. Rossignol (1961) reported absolute frequencies of hystrichospheres in recent sediments in the eastern Mediterranean Sea off the coast of Israel. In their study of palynomorph sedimentation in the Bahamas, Traverse and Ginsburg (1966) observed that the pattern of dinocyst abundance is related to both sedimentary factors and nutrient availability. Williams (1971) demonstrated that absolute frequency of dinocysts in sediments and distribution patterns of individual dinocyst taxa can be related to surface environmental conditions in the North Atlantic. His material included samples north of Cuba and in the Caribbean, and he used principle components analysis to recognize ten biofacies. Davey (1971) and Davey and Rogers (1975) studied absolute and relative abundances of dinocyst species in shelf sediments of the coast of South Africa and South West Africa. Wall and others (1977) presented quantitative treatment of dinoflagellate-cyst assemblages from the North and South Atlantic and adjacent seas. They recognized inshore to offshore trends and climatic trends in the distributions of individual taxa, in associations of species, in species diversity, and in absolute abundances of cysts. Other low-latitude studies of dinocysts in modern sediments include Morzadec-Kerfourn (1983), Turon and Londeix (1988) from Mediterranean Sea material, Bradford and Wall (1984) on material from the Persian Gulf, Gulf of Oman and Arabian Sea, Bint (1988) on material offshore of northwestern Australia, and McMinn (1990) on material from eastern Australia and New Guinea.
The distribution of pollen in marine sediments has been studied in several areas of the world, including the northwest Atlantic Ocean (Mudie, 1982; Heusser, 1983), western Atlantic Ocean (Litwin and Andrle, 1992), Great Bahama Bank region (Traverse and Ginsburg, 1966), the Orinoco delta region (Muller, 1959), eastern Atlantic Ocean (Hooghiemstra and others, 1986; Hooghiemstra, 1988), the Mediterranean coast of Israel (Rossignol, 1961), Gulf of California (Cross and others, 1966), and the northeast Pacific Ocean (Heusser and Balsam, 1977). These works show that pollen assemblages from marine sediments are representative of the nearby terrestrial vegetation (Groot and Groot, 1966; Litwin and Andrle, 1992), although the relative abundance of taxa in the marine record differs from that on land. Typically, bisaccate pollen such as that produced by Pinus increase in relative abundance with increasing distance from shore (Heusser and Balsam, 1977; Mudie, 1982), and other taxa decrease in relative abundance accordingly.
Acknowledgments. We thank Johan Groot (Delaware Geological Survey) and Anne de Vernal (GEOTOP, UniversitŽ du Quebec ˆ MontrŽal) for helpful comments on earlier versions of this manuscript. We also thank Martin Head, Barrie Dale, and other attendees of the 1993 Neogene workshop for comments on taxonomy and taxonomic problems. We express our appreciation to the Gulf Coast Research Laboratory, Ocean Springs, Mississippi, and especially to Captain George Cantrell, for everything that made the use of the R/V Bill Demoran a success.
During April and September of 1991, more than 50 box-core samples and 5 gravity cores were collected in Mississippi Sound and the Gulf of Mexico from the R/V Bill Demoran. Samples were taken in water depths from 1-60 ft along inshore-offshore transects, and around barrier islands. All sediment samples are from shallow, nearshore sites between 0.5 and 15 mi offshore. Sediment type is predominantly silt and clay. Attempts were made to take additional samples in sandy-bottomed areas, but these samples were lost from the coring device. Here we report on the results of our palynological investigations of 33 of those samples. Samples taken near recently dredged shipping channels were excluded. All samples were taken from the top 2-4 cm of box cores, except for three samples from gravity cores (the first three samples on table 1).
Samples were processed by dissolution in hydrochloric and hydrofluoric acids. No oxidation or heavy liquid was used. Slides were sieved on a 10µm screen. Most of the samples were dried and weighed, and known amounts of residue were counted for estimates of absolute abundances of dinocysts (cysts per gram). Material was examined under a light microscope, and a few samples were also examined using a scanning electron microscope.
Dinoflagellate cysts and pollen were counted from each of the 33 samples. The dinocyst results are summarized in Table 1. Pollen counts are shown in Table 2.
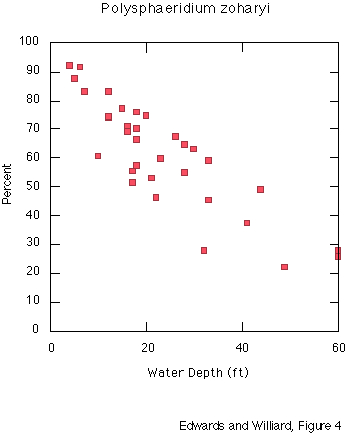
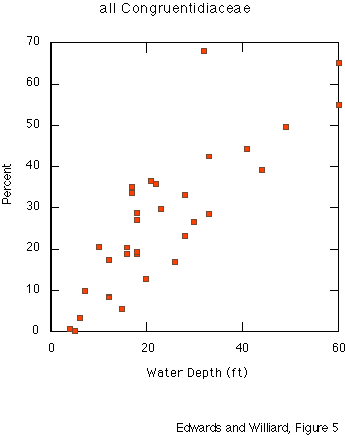
Dinocysts Polysphaeridium zoharyi (Rossignol) Lentin & Williams (thecate name: Pyrodinium bahamense Plate) dominates the dinocyst assemblages in all but eight offshore sediment samples. Cysts of peridinialean dinoflagellates (Family Congruentidiaceae) are present in some samples and nearly absent from others. P. zoharyi shows an inverse relationship with increasing water depth (fig. 4), whereas Congruentidiaceae cysts show a direct relationship with depth (fig. 5) in these shallow-water areas.
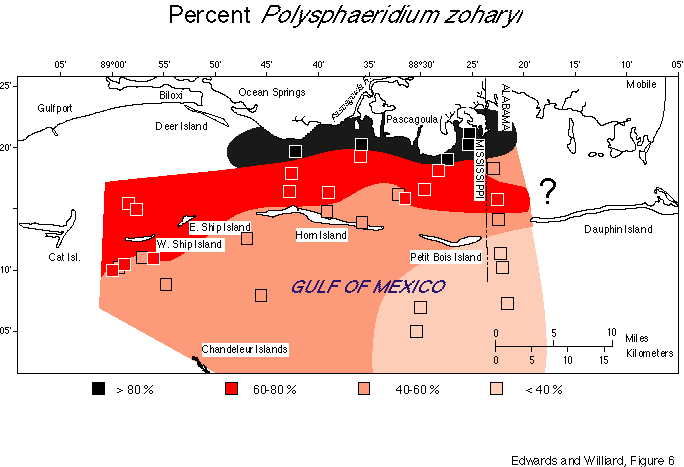
Figure 6 shows the general distribution pattern of P. zoharyi. Samples taken in water depths of less than 3 m (and less than 2 km offshore of the nearest land) were all strongly dominated by cysts of Polysphaeridium zoharyi (greater than 80 percent). Undifferentiated Spiniferites species and Multispinula cf. M. quanta Bradford made up less than 10 percent each. Only occasional specimens of Lingulodinium machaerophorum, Nematosphaeropsis rigida and Operculodinium israelianum were encountered.
The assemblages from the most offshore samples are dominated by Multispinula cf. M. quanta; P. zoharyi is second in abundance. These samples are from water depths up to 60 ft, and the furthest sample is 8 mi from the barrier island and 15 mi from the mainland. Other peridinialean forms comprise 2-16 percent of the dinocyst assemblages. Gonyaulacalean forms including Spiniferites spp., O. israelianum, L. machaerophorum, and Nematosphaeropsis rigida are present in small amounts. Samples collected between the most offshore and the most inshore sites contain 45-77 percent P. zoharyi. In general, samples near open passages between islands show relatively less dominance by P. zoharyi, and samples that are relatively sheltered behind islands show higher dominance.
The passage between Petit Bois Island and Dauphin Island is relatively shallow; samples along this transect have an intermediate character. Both P. zoharyi and M. cf. M. quanta show wide variation in size, surface features, and spine length; hence, end-members of a population may appear very different (Pl. 1, figs. 7-8; Pl. 2, figs. 2-9).
One further observation, which may be helpful in recognizing other similar environments in older sediments, is the fact that nearly all the cysts recovered in this study, regardless of taxonomic affinity, are round (spherical or discoidal) and spiny: Polysphaeridium, Multispinula, Spiniferites, Lingulodinium, Nematosphaeropsis, Operculodinium. Many of the specimens of Spiniferites spp. were poorly oriented or folded. Individual species were not differentiated. Plate 1, figs. 9-12 show two of these.
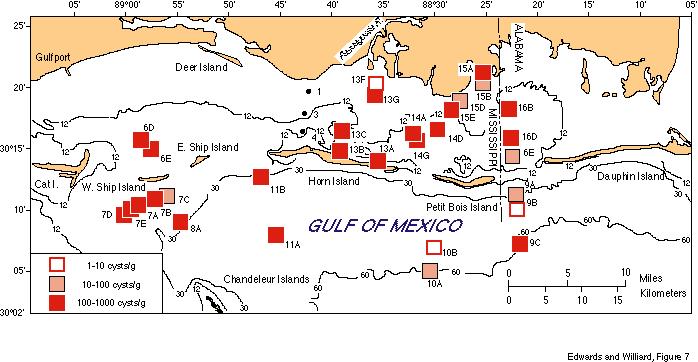
The number of dinocysts per gram of sediment was calculated for most of the counted samples (fig. 7). Counts varied by two orders of magnitude among samples and ranged from a low of 4 to a high of 570 cysts/g. Samples with low values are most typically from sandier sediments, or from areas of greater terrigenous input.
Reworked specimens of Cretaceous and early Tertiary age were encountered in a few samples. Two of these are shown in Plate 1, figs, 14 and 15.
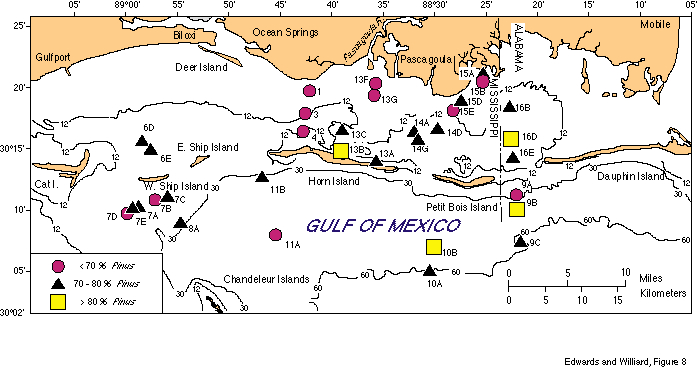
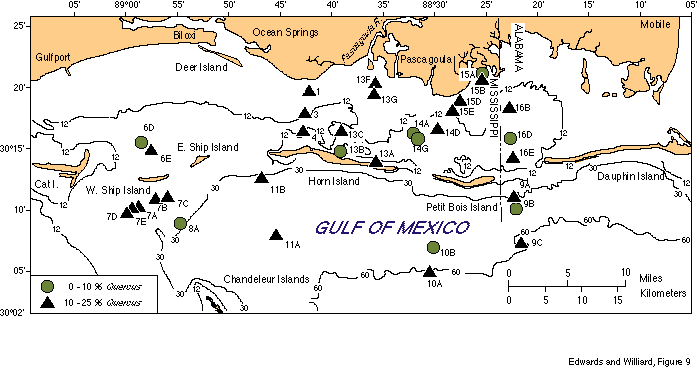
Pollen assemblages (Table 2) are dominated uniformly by Pinus pollen (55-90 percent) with Quercus pollen ranking second in abundance (2-23 percent). Other tree taxa commonly present in small amounts (less than 5 percent) include Carya, TCT (pollen of Taxodiaceae, Cupressaceae, Taxaceae), Ulmus, Juglans, Salix, Nyssa, and Betula. Non-arboreal pollen (NAP) is a relatively small component, comprising from 2-16 percent of the assemblages and consists primarily of members of the aster and grass families. Figures 8 and 9 show the percent abundances of Pinus and Quercus in the studied samples.
Assemblages from these samples are very similar to each other; the abundances of Pinus and Quercus pollen are the only notable variations. Few patterns in pollen distribution are evident, due to the relatively small area studied and the uniformity of vegetation onshore. In general, Pinus pollen is most abundant from deeper water sediments (Fig. 8). Quercus shows no coherent distribution pattern (Fig. 9), and pollen of other taxa are too rare (less than 5 percent) to track differences in their abundance throughout the area.
Dinocyst assemblages from the very nearshore sediments in Mississippi Sound are dominated by P. zoharyi. These assemblages are similar to ones reported from the Bahamas and Phosphorescence Bay, Puerto Rico (Wall and others, 1977), the Persian Gulf (Bradford and Wall, 1984), and coastal New Guinea (McMinn, 1990). Although P. zoharyi often is regarded as an indicator of highly saline conditions (Morzadec-Kerfourn, 1983, 1988), here it is apparently an indicator of abnormally low salinity, or perhaps of highly fluctuating salinity. Wall and others (1977) reported that the thecate stage is known to migrate vertically to avoid direct contact with incoming freshwater.
Dinocyst assemblages from shallow-water sediments in the Gulf of Mexico contain large numbers of Multispinula cf. M. quanta. Similar assemblages are not known. Perhaps this form is readily destroyed, either in the sediments or in the processing laboratory. The relative rarity of this form in the three gravity-core samples (taken below the top few centimeters) is noteworthy. Other peridinialean forms are conspicuous and comprise up to 16 percent of the assemblage.
Although P. zoharyi shows a strong negative correlation, and Multispinula cf. M. quanta shows a strong positive correlation, with water depth, the controlling factor is unlikely to be depth per se. All samples are quite shallow. More likely, the proximity to terrigenous freshwater input -- which relates to nutrient delivery and to salinity fluctuations -- is more important. The pattern of very high dominance of P. zoharyi in sheltered areas also suggests that currents play a role. Currents could deliver different nutrients or more saline water, but probably also deliver cysts from dinoflagellates that are more abundant elsewhere into areas where the cysts are not normally present.
Pollen assemblages from these shallow marine sediments are virtually indistinguishable from each other and from those collected along the east coast of Florida and Georgia (see Litwin and Andrle, 1992). All are dominated strongly by Pinus pollen with Quercus pollen subdominant, and pollen of various trees (Carya, Juglans, TCT, Ulmus) are present in consistently small amounts. These assemblages represent the shallow marine record of the southern mixed forest, which consists primarily of pine-dominated forests with abundant oak and a variety of hardwoods and shrubs (Christensen, 1988). The southern mixed forest covers most of Florida, southern Mississippi and Alabama, and southeastern Georgia (Küchler, 1985).
The shallow marine record described here, and the terrestrial pollen record of the southern mixed forest, show no notable compositional differences. Rather, the two records differ primarily in abundance of Pinus pollen and the pollen of deciduous trees. Pollen assemblages from lake deposits in Florida and Georgia have 45 to 65 percent Pinus pollen (Watts, 1969, 1970, 1975, 1980; Watts and Stuiver, 1980), whereas the shallow marine assemblages show 57 to 88 percent Pinus pollen. Quercus pollen comprises about 10 to 20 percent of both the terrestrial and shallow marine records, but many hardwoods are slightly more abundant in the terrestrial pollen record. For example, Carya typically comprises about 5 percent of the terrestrial assemblages but only up to 2.7 percent in shallow marine assemblages.
The dinocyst and pollen assemblages in the shallow marine sediments of Mississippi Sound and the Gulf of Mexico reflect both ecologic factors and transport history. Polysphaeridium zoharyi (Rossignol) Lentin & Williams dominates all but eight dinocyst assemblages and shows an inverse relationship with increasing depth. It is also relatively less abundant in samples near open passages between barrier islands than in samples that are relatively sheltered behind islands. Although this species is often regarded as an indicator of highly saline here it is most abundant in an area of abnormally low, and highly fluctuating, salinity. Cysts of the Congruentidiaceae are present in some samples and nearly absent from others. In these very shallow-water samples, these cysts, including Multispinula cf. M. quanta, show increasing abundance with increasing depth. Regardless of taxonomic affinity, most dinocysts in this study are round and spiny: Polysphaeridium, Multispinula, Spiniferites, Lingulodinium, Nematosphaeropsis, Operculodinium. P. zoharyi and M. cf. M. quanta show wide variation in size, surface features, and spine length. The pattern of dinocyst diversity near and between barrier islands suggests that currents deliver cysts to bottom sediments.
Pollen assemblages in the Mississippi Sound and Gulf of Mexico are dominated uniformly by Pinus pollen; Quercus pollen ranks second in abundance. Other tree taxa are present in small amounts and non-arboreal pollen (NAP) is a relatively small component of the assemblages. Little variation exists among pollen assemblages from these samples; Pinus pollen is most abundant from deeper sediments, and, although its abundance is variable, Quercus pollen shows no distinctive distribution pattern in this area. The pollen assemblages represent the shallow marine record of the southern mixed forest, which consists primarily of pine-dominated forests with abundant oak and a variety of hardwoods and shrubs and which covers most of Florida, southern Mississippi and Alabama, and southeastern Georgia.
Bint, A.N., 1988, Recent dinoflagellate cysts from Mermaid Sound, northwestern Australia: Memoir of the Association of Australasian Palynologists, v. 5, p. 329-341.
Bradford, M.R., and Wall, D.A., 1984, The distribution of recent organic-walled dinoflagellate cysts in the Persian Gulf, Gulf of Oman, and northwestern Arabian Sea: Palaeontographica Abt. B, v. 192, p. 16-84.
Christensen, N.L., 1988, Vegetation of the southeastern coastal plain: in Barbour, M.G., and Billings, W.D., eds., North American Terrestrial Vegetation: Cambridge, Cambridge University Press, p. 317-364.
Cross, A.T., Thompson, G.G., and Zaitzeff, J.B., 1966, Source and distribution of palynomorphs in bottom sediments, southern part of Gulf of California: Marine Geology, v. 4, p. 467-524.
Davey, R.J., 1971, Palynology and palaeo-environmental studies, with special reference to the continental shelf sediments of South Africa: in Farinacci, A. (ed.), Proceedings of the 2nd International Conference on Planktonic Microfossils, Roma, 1970: Rome, Edizioni Technoscienza, p. 331-347.
Davey, R.J., and Rogers, J., 1975, Palynomorph distribution in Recent offshore sediments along two traverses off South West Africa: Marine Geology, v. 18, p. 213-225.
Eleuterius, C.K., and Beaugez, S.L., 1979, Mississippi Sound: a hydrographic and climatic atlas: Blossman Printing, Ocean Springs, Mississippi, 135 p.
Groot, J.J., and Groot, C.R., 1966, Marine palynology: possibilities, limitations, problems: Marine Geology, v. 4., p. 387-395.
Heusser, L.E., 1983, Pollen distribution in the bottom sediments of the western North Atlantic Ocean: Marine Micropaleontology, v. 8, p. 77-88.
Heusser, L.E., and Balsam, W.L., 1977, Pollen distribution in the northeast Pacific Ocean: Quaternary Research, v. 7, p. 45-62.
Hooghiemstra, H., 1988, Palynological records from northwest African marine sediments: a general outline of the interpretation of the pollen signal: Philosophical Transactions of the Royal Society of London, Series B, v. 318, p. 431-449.
Hooghiemstra, H., Agwu, C.O.C., and Beug, H.-J., 1986, Pollen and spore distribution in recent marine sediments: a record of NW-African season wind patterns and vegetation belts: "Meteor" Forschungsergebnisse, Reihe C, v. 40, p. 87-135.
Küchler, A.W., 1985, Potential Natural Vegetation -- National Atlas of the United States of America: U.S. Geological Survey, 2nd edition, 1:7,500,000.
Litwin, R.J., and Andrle, V.A.S., 1992, Modern palynomorph and weather census data from the U.S. Atlantic Coast (Continental Margin Program samples and selected NOAA weather stations): U.S. Geological Survey Open-File Report 92-263, p. 1-31.
McMinn, Andrew, 1990, Recent dinoflagellate cyst distribution in eastern Australia: Review of Palaeobotany and Palynology, v. 65, p. 305-310.
Morzadec-Kerfourn, M.-T., 1983, Intérêt des kystes de dinoflagellés pour l'établissement de reconstitution paléogéographique: exemple du Golf de Gabès (Tunisie): Cahiers de Micropaléontologie, v. v, p. 15-22.
Morzadec-Kerfourn, M.-T., 1988, Paléoclimats et paléoenvironnements du Tardiglaciaire au Récent, en Méditerranée oriental, à l'est du delta du Nil; l'apport des microfossiles à membrane organique: Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, v. 12, no. 1, p. 267-275, fig. 1-3.
Mudie, P.J., 1982, Pollen distribution in recent marine sediments, eastern Canada: Canadian Journal of Earth Sciences, v. 19, p. 729-747.
Muller, Jan, 1959, Palynology of Recent Orinoco delta and shelf sediments: Reports of the Orinoco Shelf Expedition; Volume 5: Micropaleontology, v. 5, no. 1, P. 1-32, pl. 1.
National Climatic Center, 1983, Climate Normals for the U.S. (Base:1951-1980): Detroit, Gale Research Co., 712p. National Oceanographic and Atmospheric Administration, 1985, Climates of the States, vol. 1, Alabama-New Mexico: Detroit, Gale Research Co., 758 p.
Robinson, M.K., Bauer, R.A., and Schroeder, E.H., 1979, Atlas of North Atlantic - Indian Ocean monthly mean temperatures and mean salinities of the surface layer: Naval Oceanographic Office, Bay St. Louis. Rossignol, Martine, 1961, Analyse pollinique de sŽdiments marins quaternaires en Isra‘l: I: SŽdiments rŽcents: Pollen et Spores, v. 3, no. 2, p. 303-324.
Traverse, A., and Ginsburg, R.N., 1966, Palynology of the surface sediments of Great Bahama Bank, as related to water movement and sedimentation: Marine Geology, v. 4, p. 417-459.
Turon, J.-L., and Londeix, L., 1988, Les assemblages de kystes de dinoflagellés en Méditerranée occidentale (Mer d'Alboran). Mise en évidence de l'évolution des paléoenvironnements depuis le dernier maximum glaciaire: Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, v. 12, no. 1, p. 13-344, pl. 1-7.
Wall, D., Dale, B., Lohmann, G.P., and Smith, W.K., 1977, The environmental and climatic distribution of dinoflagellate cysts in modern marine sediments from regions in the North and South Atlantic Oceans and adjacent seas: Marine Micropaleontology, v. 2, p. 121-200.
Watts, W.A., 1969, A pollen diagram from Mud Lake, Marion County, north-central Florida: Geological Society of America Bulletin, v. 80, p. 631-642.
Watts, W.A., 1970, The full-glacial vegetation of northwestern Georgia: Ecology, v. 51, no. 1, p. 17-33.
Watts, W.A., 1975, A late Quaternary record of vegetation from Lake Annie, south-central Florida, Geology, v. 3, p. 344-346.
Watts, W.A., 1980, The late Quaternary vegetation history of the southeastern United States: Annual Review of Ecology and Systematics, v. 11, p. 387-409.
Watts, W.A., and Stuiver, M., 1980, Late Wisconsin climate of northern Florida and the origin of species-rich deciduous forest: Science, v. 210, p. 325-327.
Williams, D. B., 1971, The occurrence of dinoflagellates in marine sediment, in Funnel, D. B., and Riedel, eds., The Micropaleontology of Oceans: Cambridge, Cambridge University Press, p. 231-243.
Gonyaulacalean cysts and reworked cysts from the Gulf of Mexico and Mississippi Sound.
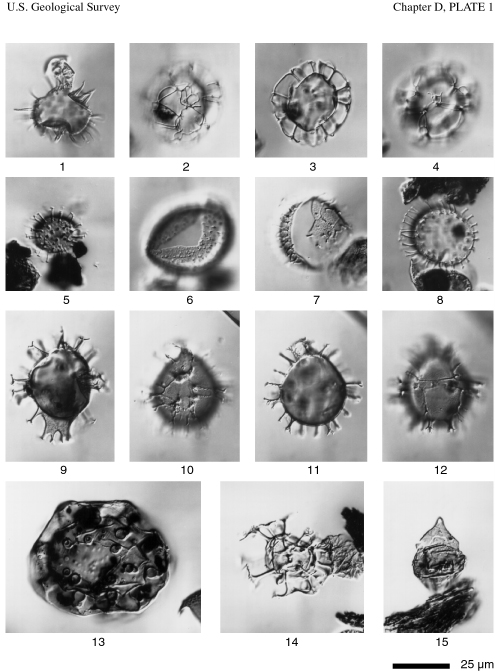
Figure 1. Lingulodinium machaerophorum (Deflandre & Cookson) Wall. Ventral view at midfocus, sample R4518A (4), GC-4. Edwards slide coordinates, 36.8x109.1.
Figures 2-4. Nematosphaeropsis rigida Wrenn. Ventral views of ventral surface, optical section, dorsal surface, sample R4518A (2), GC-4. Edwards slide coordinates, 34.3x108.0.
Figure 5. Operculodinium centrocarpum (Deflandre & Cookson) Wall sensu Wall, 1967. Slightly oblique ventral view of ventral surface, sample R4611 (1), GC-10A. Edwards slide coordinates, 27.3x90.2.
Figure 6. Operculodinium israelianum (Rossignol) Wall. Ventral view of dorsal surface, sample R4515A (1), GC-1. Edwards slide coordinates, 20.4x86.5.
Figure 7. Polysphaeridium zoharyi (Rossignol) Bujak et al. Apical view of apical region, sample R4519A (4), GC-5. Edwards slide coordinates, 33.7x76.3.
Figure 8. Polysphaeridium zoharyi (Rossignol) Bujak et al. Antapical view of antapex, sample R4515A (1), GC. Edwards slide coordinates, 36.4x88.6.
Figure 9. Spiniferites mirabilis (Rossignol) Sarjeant. Dorsal view at midfocus, sample R4518A (4), GC-4. Edwards slide coordinates, 20.7x72.5.
Figures 10-12. Spiniferites sp. Ventral view of ventral surface, optical section, dorsal surface, sample R4519A (4), GC-5. Edwards slide coordinates, 32.4x89.4.
Figure 13. Tuberculodinium vancampoae (Rossignol) Wall. Apical view of apex, sample R4518A (4), GC-4. Edwards slide coordinates, 31.0x80.2.
Figure 14. Areosphaeridium sp. Orientation uncertain, sample R4516A (3), GC-2. Edwards slide coordinates, 17.8x87.6. This specimen is reworked from Paleogene sediments.
Figure 15. Chatangiella sp. Dorsal view of dorsal surface, sample R4627 (2), GC-13G. Edwards slide coordinates, 21.0x79.5. This specimen is reworked from Cretaceous sediments.
Peridinialean cysts from the Gulf of Mexico and Mississippi Sound.

Figure 1. Multispinula quanta Bradford. Apical view at midfocus, sample R4611 (1), GC-10A. Edwards slide coordinates, 25.1x97.1. Note the typical reniform outline and smooth surface texture.
Figure 2,3. Multispinula cf. M. quanta Bradford. Antapical? view at midfocus, sample R4612 (1), GC-10B. Edwards slide coordinates, 18.2x106.1. The outline is reniform but the surface texture is scabrate.
Figure 4,5. Multispinula cf. M. quanta Bradford. Apical? view of upper and lower surfaces, sample R4619A (4), GC-5. Edwards slide coordinates, 37.2x80.2. The outline is circular and the surface texture is scabrate.
Figure 6,9. Multispinula cf. M. quanta Bradford. Dorsal view of dorsal and ventral surfaces, sample R4623 (2), GC-13C. Edwards slide coordinates, 35.0x83.3.
Figure 7. Brigantedinium simplex Wall ex Lentin & Williams. Antapical view of apical surface, sample R4613 (2), GC-11A. Edwards slide coordinates, 19.7x102.5.
Figure 8. Multispinula? sp. Apical view of apex, sample R4619A (4), GC-5. Edwards slide coordinates, 34.7x75.3.
Figure 10. cyst of Protoperidinium sp. Ventral view of dorsal surface, sample R4518A (4), GC-4. Edwards slide coordinates, 31.6x103.6.
Figure 11. cyst of Protoperidinium oblongum(Aurivillius) Balech. Dorsal view of dorsal surface, sample R4613(2), GC-11A. Edwards slide coordinates, 29.8x83.2.
Figure 12. Stelladinium stellatum (Wall & Dale) Reid. Orientation uncertain, sample R4613(2), GC-11A. Edwards slide coordinates, 28.1x83.8.

![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: http://pubsdata.usgs.gov/pubs/of/2001/of01-415/chap4txt.htm
Page Contact Information: USGS Publications Team
Page Last Modified: Wednesday, 07-Dec-2016 18:56:36 EST