Open-File Report 01-415
Ostracode assemblages were studied from the Pleistocene Biloxi Formation in Jackson County, Mississippi. These assemblages reflect outer bay, inlet, and inner shelf paleoenvironments (with near-normal salinities) of the last interglacial highstand of sea level.
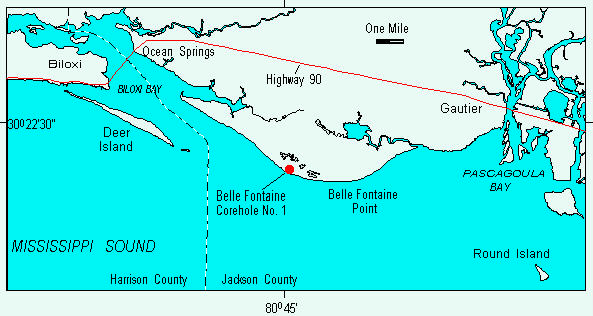
A stratigraphic test hole drilled in Jackson County, Mississippi recovered fossiliferous marine and marginal-marine sediments of the Biloxi Formation. The Biloxi Formation is an important late Pleistocene unit that represents a period of warm climate and high global sea level during the last interglacial period (Otvos, 1975, 1981, 1991). This paper describes the ostracodes from the Biloxi sediments from the Belle Fontaine No. 1 corehole and provides a brief paleoenvironmental interpretation based on the modern ecology of the ostracode species.
There are few previous studies of ostracodes from Pleistocene deposits of the Gulf of Mexico Coastal Plain. Cronin (1986) studied late Pleistocene and Holocene ostracodes from a series of coreholes from Corpus Christi Bay, Texas, and illustrated many of the species that also occur in the Belle Fontaine core. Otvos (1976) listed ostracodes from the Biloxi Formation that were identified by Paul Krutak. In contast to studies of fossil faunas, there are many studies of modern ostracode faunas from bays, lagoons, and continental-shelf environments along the Gulf Coast. Garbett and Maddocks (1979), Cronin (1979, 1986), and Kontrovitz (1976) provide useful reviews of previous ostracode studies.
The Belle Fontaine Beach No. 1 corehole was drilled in 1990 in the Gautier South 7-1/2' quadrangle at a surface elevation of 6 feet (Figure 1). Figure 2 shows the lithology of the Biloxi Formation and the location of the ostracode samples. Ten samples were studied from the interval between depths of 29.8 and 47.3 ft. Sediment was washed through a 63-micron sieve and ostracodes were picked from the greater than 150 micron size fraction. Species wee identified following the taxonomy of Garbett and Maddocks (1979) and Cronin (1979, 1986). Table 1 shows the number of specimens of each ostracode taxon in the ten samples. A total of 1,535 individual specimens and more than 42 species were found in the Biloxi Formation (species of Hulingsina were not separated).
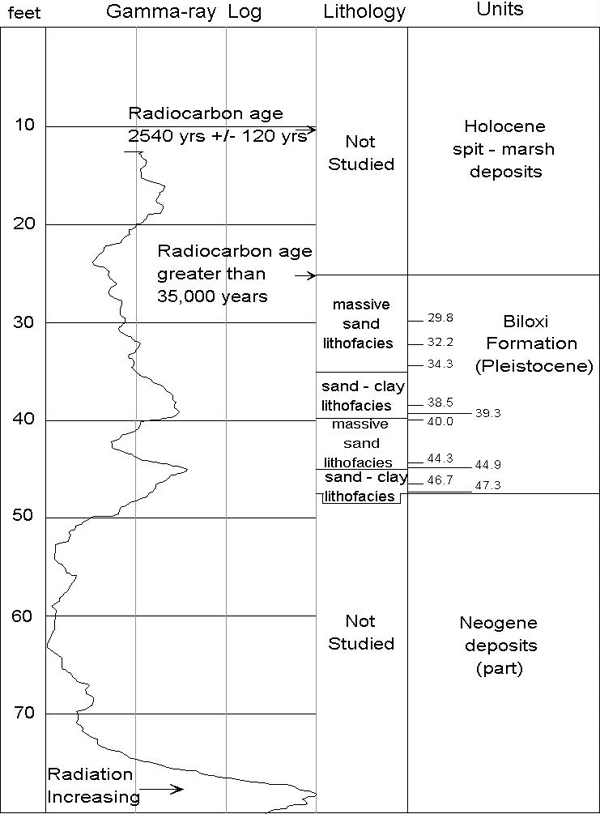
| Taxon / Feet in Core | 29.8 | 32.2 | 34.3 | 38.5 | 39.3 | 40.0 | 44.3 | 44.9 | 46.7 | 47.3 |
|---|---|---|---|---|---|---|---|---|---|---|
| *Actinocythereis cf. bahamaensis (Brady, 1868) | 1 | 1 | . | . | . | . | 1 | 1 | . | . |
| Actinocythereis subquadrata Puri, 1953 | 3 | 4 | 1 | 7 | 6 | . | 3 | 1 | 4 | 2 |
| Aurila laevicula Edwards, 1944 | 1 | 1 | . | . | 3 | . | 2 | . | . | 1 |
| *Campylocythere laeva Edwards, 1944 | . | . | . | . | 1 | . | 1 | . | . | . |
| *Climacoidea (Protoconcha) gigantica (Edwards, 1944) | 2 | 1 | . | . | 1 | . | 3 | . | 1 | . |
| Climacoidea (Protoconcha) multipunctata (Edwards, 1944) | . | . | . | . | 1 | . | 2 | . | 1 | . |
| **Climacoidea (Protoconcha) nelsonensis (Grossman, 1967) | 8 | 12 | 8 | 5 | . | 4 | 1 | 1 | 1 | 5 |
| Climacoidea (Protoconcha) tuberculata (Puri, 1960) | 7 | 5 | 5 | 15 | . | 5 | 2 | 7 | . | |
| *Cytherella sp. | . | . | . | . | . | . | . | . | . | 2 |
| *Cytherelloidea sp. | . | . | 1 | 1 | . | . | . | . | . | 1 |
| Cytheretta litorea Garbett and Maddocks, 1979 | 6 | 5 | 3 | 5 | 9 | 1 | 4 | 2 | 4 | 2 |
| Cytheretta louisianensis Kontrovitz, 1976 | . | 4 | . | . | 2 | . | 3 | 4 | 3 | 1 |
| **Cytheromorpha paracastanea (Swain, 1955) | 4 | . | 2 | 1 | . | . | . | 1 | . | 1 |
| **Cytherura cybaea Garbett and Maddocks, 1979 | 11 | 5 | 7 | . | 2 | . | . | . | 2 | 2 |
| Cytherura forulata Edward, 1944 | 11 | 19 | 11 | 4 | 10 | 2 | 2 | 6 | 8 | 15 |
| **Cytherura nucis Garbett and Maddocks, 1979 | . | . | . | . | . | . | . | 3 | . | . |
| *Cytherura cf. C.pseudostriata Hulings, 1966 | 1 | 3 | 2 | . | . | 1 | 2 | . | 1 | . |
| *Cytherura sablensis Benson and Coleman, 1963 | 1 | 2 | 1 | 2 | . | 1 | 2 | . | 1 | . |
| **Cytherura sandbergi Morales, 1966 | 9 | 11 | 5 | . | 9 | . | 3 | 1 | 2 | 1 |
| Cytherura valentinei Garbett and Maddocks, 1979 | 3 | 7 | 1 | 5 | . | . | 2 | 18 | 3 | 8 |
| Hulingsina spp. | 46 | 50 | 4 | 19 | 90 | 6 | 17 | 9 | 29 | 32 |
| **Loxoconcha moralesi Kontrovitz, 1976 | 3 | 6 | 2 | 35 | 10 | . | 20 | 39 | 10 | 13 |
| *Loxoconcha reticularis Edwards, 1944 | . | 2 | . | . | . | . | 1 | . | . | . |
| Malzella floridana (Benson and Coleman, 1963) | 3 | 1 | 3 | 2 | 11 | 2 | 3 | . | 1 | 3 |
| **Megacythere repexa Garbett and Maddocks, 1979 | 1 | 6 | 4 | 1 | 6 | . | . | . | 1 | . |
| *Neonesidea sp. | . | . | . | . | 5 | . | 1 | . | . | . |
| *Paracytheridea altila Edwards, 1944 | 10 | 9 | 8 | 2 | 21 | 1 | 4 | 2 | 1 | 9 |
| *Paracytheridea hazeli Cronin, 1990 | 1 | . | . | 1 | 10 | . | . | . | . | 1 |
| *Paracytheridea rugosa Edwards, 1944 | . | . | . | . | . | . | . | . | 1 | . |
| **Paracytheroma stephensoni (Puri, 1954) | 6 | 12 | 3 | . | . | . | 2 | 2 | 7 | 6 |
| ** Paracytheroma texana Garbett and Maddocks, 1979 | 1 | . | . | . | . | . | . | 1 | . | . |
| Paradoxostoma delicata Puri, 1954 | 1 | 1 | . | 3 | . | . | . | . | 4 | 2 |
| Pellucistoma magniventra Edwards, 1954 | . | 14 | 8 | . | . | . | . | 2 | 4 | 2 |
| Peratocytheridea bradyi (Stephenson, 1938) | 26 | 15 | 19 | . | 58 | 7 | 8 | 4 | 12 | 12 |
| **Peratocytheridea setipunctata (Brady, 1869) | 12 | 20 | 2 | 2 | 14 | 4 | 6 | . | 7 | 5 |
| **Perissocytheridea spp. | . | 1 | . | . | . | . | . | 1 | . | 1 |
| Pontocythere spp. | 2 | 1 | 1 | 2 | . | . | . | 1 | 2 | 1 |
| *Pterygocythereis sp. | . | . | . | 1 | . | . | . | . | . | 1 |
| *Pumilocytheridea sandbergi Bold | 2 | . | 2 | 4 | . | 1 | . | . | . | 4 |
| *Puriana carolinensis Hazel, 1983 | 14 | 5 | . | 1 | 7 | 4 | 3 | . | 3 | 5 |
| *Puriana convoluta Teeter, 1975 | 1 | 1 | . | . | 4 | 1 | 2 | . | 1 | . |
| *Puriana floridana Puri, 1960 | 7 | 9 | 7 | 5 | 14 | . | 3 | 11 | 9 | 4 |
| Other | . | 2 | . | . | . | . | . | 4 | 1 | . |
| Total | 204 | 235 | 148 | 108 | 309 | 35 | 106 | 117 | 131 | 142 |
*primarily lives on continental shelf
**primarily lives in bays
The most common ostracode genera are Hulingsina, Cytherura, Climacoidea (Proteoconcha), Loxoconcha, and Peratocytheridea. Loxoconcha moralesi and Peratocytheridea bradyi were the two most common species, averaging 11 percent or more of each assemblage. This is a late Pleistocene and Holocene fauna that lacks characteristic Neogene and early Pleistocene species.
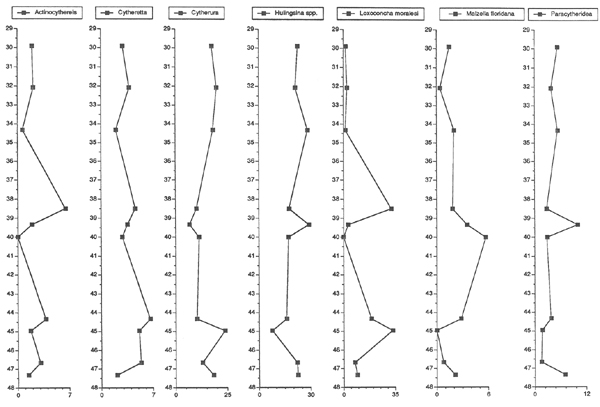
Plots of the percentages of common taxa in the Billoxi Formation (Figures 3 and 4) show that relatively minor changes in the assemblage composition occurred throughout the Biloxi Formation at this locality. A few notable exceptions are the high percentages of Peratocytheridea, Puriana, Malzella, and Paracytheridea, near 39.5 to 40 ft, followed by peak percentages of L. moralesi and Actinocythereis at 38.5 ft. L. moralesi also is common at 45 ft. Paracytheroma stephensoni and Pellucistoma magniventra also have relatively high percentages in the upper and lowermost parts of the core but are absent in the middle part.

The following papers on the distribution and ecology of modern ostracodes from the Gulf of Mexico region were used to interpret the paleoenvironmental significance of the Biloxi fauna: Benson and Coleman (1963), King and Kornicker (1970), Keyser (1975, 1977), Kontrovitz (1976), Garbett and Maddocks (1979), and references in these papers. The overall Biloxi ostracode fauna is a mixture of species that today inhabit benthic environments of the inner continental shelf, bays behind barrier islands, or both. For example, seventeen species from the Biloxi Formation do not occur or occur sporadically in bays along the Texas coast (Garbett and Maddocks, 1979) (see Table 1). Conversely, eleven species from the Biloxi generally do not occur in sediments on the continental shelf of the Gulf of Mexico (Table 1). Therefore, the Biloxi assemblages are considered to be death assemblages representing various degrees of post-mortum spatial and temporal mixing during sedimentation and burial. It is likely that much of the Biloxi at this locality represents deposition near a tidal inlet of a barrier island complex where many of these species live, or where mixing of shelf and bay species might occur.
The paleosalinity of the Biloxi was most likely polyhaline (20-30 parts per thousand) to occasionally euhaline (30-35 ppt). Keyser (1975, 1977) showed the salinity tolerance of many Biloxi species in his detailed study of ostracodes from brackish water environments in the Everglades of Florida. Some notable examples are: Paracytheroma stephensoni (more than 24 ppt), Cytheromorpha paracastanea (greater than 24 ppt), Actinocythereis subquadrata (greater than 22 ppt), Aurila laevicula (=A. amygdala of Keyser) (more than 17 ppt), and Cytherura forulata (greater than 14 ppt). All these species except C. paracastanea occur in euhaline salinities on the continental shelf.
The following features of the Biloxi Formation ostracode fauna indicate that late Pleistocene paleoenvironments, although near the coast and sometimes of reduced salinity, were probably never completely isolated from the open ocean. First, there are very few specimens of Perissocytheridea. Species of this genus predominate in upper and mid-bay environments of Texas bays where salinity is below 2 ppt. Second, species of Puriana, Paracytheridea, and Cytheretta, which comprise significant proportions of Biloxi assemblages, live primarily on the continental shelf in euhaline conditions; they are rarely found in modern sediments behind barrier islands. Third, Loxoconcha matagordensis, another predominant inner and mid-bay species, does not occur in the Biloxi fauna. The most restricted paleoenvironments seem to have occurred at 38.5 ft and 44.3 ft, where Loxoconcha moralesi reached 32 to33 percent of the assemblages. However, this species, which reaches its greatest proportions in modern mid-bay environments (Garbett and Maddocks, 1979), also occurs in shelf sediments (Kontrovitz, 1976), and its occurrence in the Biloxi with typical shelf taxa (Puriana, Paracytheridea) at these horizons might signify mixed death assemblages. Thus, it is doubtful that paleosalinities during the deposition of the Biloxi in this region of Mississippi Bay were below 20 ppt and most often were 25 to 30 ppt.
Substrate was another factor controlling the distribution of ostracode species, and changes through time between sandy, silty, and muddy substrates may also explain minor changes in the abundance of various species. For example, Climaciodea (Protoconcha) nelsonensis and Peratocytheridea setipunctata live in sandy substrates of Chincoteague Bay, Virginia but do not live in nearby muddy salt marsh environments (Cronin, unpublished data). The abundance of these two taxa at 40 ft may reflect a more sandy substrate as well as reduced salinity. Paracytheroma stephensoni prefers silt substrates (Keyser, 1977), and its abundance, along with Pellucistoma magniventra, in the upper and lower parts of the core may be related substrate.
The modern environment in which ostracode assemblages most like those of the Biloxi live today is outer Corpus Christi Bay and the adjacent continental shelf outside the opening of the bay. Corpus Christi Bay is a relatively open, narly euhaline environment, where there is a mixture of species having different ecologies occurring in the bottom sediments. Even taxa such as Pterygocythereis, Cytherella, and Cytheretta, which do not live behind barrier islands, occasionally occur in bottom sediments in Corpus Christi Bay, probably reflecting their transport from the inner shelf (Garbett and Maddocks, 1979).
Benson, R.H., and Coleman, G.L., 2nd, 1963, Recent marine ostracodes from the eastern Gulf of Mexico: Kansas University Paleontological Contributions 31, Arthropoda, article 2, 52 p.
Cronin, T.M., 1979, Late Pleistocene marginal marine ostracodes from the southeastern Atlantic Coastal Plain and their paleoenvironmental implications: Geographique Physique et Quaternaire, v. 33, no. 2, p. 121-173.
_____, 1986, Ostracodes from late Quaternary deposits, South Texas Coastal Complex, in Schiedler, J., ed., Stratigraphic studies of a late Quaternary barrier-type coastal complex, Mustang Island-Corpus Christi Bay area, south Texas coast: U.S. Geological Survey Professional Paper 13228-C, p. 33-64.
Garbett, E.C., and Maddocks, R.F., 1979, Zoogeography of Holocene cytheracean ostracodes in the bays of Texas: Journal of Paleontology, v. 53, no. 4, p. 841-919.
Keyser, D., 1975, Ostracodes of the Mangroves of South Florida, their ecology and biology: Bulletin of American Paleontology, v. 65, no. 282, p. 489-499.
_____, 1977, Ecology and zoogeography of Recent brackish-water Ostracoda (Crustacea) from south-west Florida, in Sixth International Ostracode Symposium, Saalfelden, p. 207-222.
King, C.E., and Kornicker, L.S., 1970, Ostracoda in Texas bays and lagoons - An ecological study: Smithsonian Contributions to Zoology 24, 92 p.
Kontrovitz, M., 1976, Ostracoda from the Louisiana Continental Shelf: Tulane Studies in Geology and Paleontology, v.12, no. 2, p. 49-100.
Otvos, E.G., 1975, Late Pleistocene transgressive unit (Biloxi Formation), northern Gulf Coast: Bulletin American Association of Petroleum Geologists, v. 59, p. 148-154.
_____, 1976, Post-Miocene geological development of the Mississippi-Alabama coastal zone: Journal of Mississippi Academy of Sciences, v. XXI, p. 101-114.
_____, 1981, Barrier island formation through nearshore aggradation - stratigraphic and field evidence: Marine Geology, v. 43, p. 195-243.
_____, 1991, Houston Ridge, SW Louisiana - end link in the late Pleistocene Ingleside barrier chain? Prairie Formation newly defined: Southeastern Geology, v. 31, p. 235-249.

![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: http://pubsdata.usgs.gov/pubs/of/2001/of01-415/chap6txt.htm
Page Contact Information: USGS Publications Team
Page Last Modified: Wednesday, 07-Dec-2016 18:56:37 EST
