Open-File Report 01-415
Benthic foraminifera are noted for their great intraspecific phenotypic variability, much of which can be related to variation among environmental properties such as temperature, salinity, dissolved oxygen, nutrient levels, and light intensity (e.g. Bradshaw, 1957, 1961; Chang and Kaesler, 1974; Wang and Lutze, 1986; Jorissen, 1988). Paralic environments are particularly subject to wide seasonal, tidal, and daily variability, and the resultant phenotypic variations among paralic foraminifera have caused many researchers to assign ecophenotypes of single species to several different species (Schnitker, 1974; Poag, 1978; Walton and Sloan, 1990). I published a study of modern benthic foraminifera of San Antonio Bay, Texas, in which I correlated the dominance of observed ecophenotypes (formae) of Ammonia parkinsoniana, Elphidium gunteri, and Elphidium galvestonense with scope="col"temperatures and salinity patterns measured in the bay (Poag, 1978). Ecophenotypes concentrated in areas of maximum salinity and temperature included Ammonia parkinsoniana forma tepida, Elphidium gunteri forma typicum, and Elphidium galvestonense forma typicum. Areas of minimum salinity and temperature were characterized by Ammonia parkinsoniana forma typica, Elphidium gunteri forma salsum, and Elphidium galvestonense forma mexicanum.
Because these species (or their close relatives) are known to have inhabited Gulf Coast paralic environments since the Oligocene (Poag, 1966, 1987), it is reasonable to assume that similar paleoecophenotypes were produced by paleoenvironmental variations. This pilot study was designed to determine the presence and nature of these paleoecophenotypes, and to evaluate their potential for interpreting stratigraphic successions of Quaternary bioecozones (stratigraphic zones defined by the ecologic conditions inferred from their fossils).
I obtained 12 samples from the upper 49 ft of the USGS-Belle Fontaine No. 1 (BelF No. 1) core, taken at Belle Fontaine Beach in southern Jackson County, Mississippi (fig. 1), in 1990. I split each sample into aliquots containing approximately 500 to 1000 specimens of Ammonia and Elphidium (combined), and extracted all specimens for quantitative analysis. Quantitative census data are presented in the Appendix.
I was able to identify most of the same paleoecophenotypes (formae) of Ammonia and Elphidium in the BelF No. 1 samples that I identified in SAB as ecophenotypes (fig. 2). Moreover, no new paleoecophenotypes (formae not identified in SAB) were observed in the Quaternary samples. Also, the maximum and minimum concentrations of paleoecophenotypes in the core were generally the same as those of analogous ecophenotypes in SAB. The most consistent and marked faunal relations are the dominance in all samples (except one) of E. gunteri forma salsum (pl. 1, figs. G, H) and its inverse abundance (in all but the same single sample) to that of A. parkinsoniana forma tepida (pl. 1, figs. A, B). The mexicanum forma of Elphidium galvestonense (pl. 1, figs. E, F) also occurred in all samples, but because its forma typicum was entirely missing (Appendix), the relative importance of the two E. galvestonense paleoecophenotypes could not be fully evaluated.
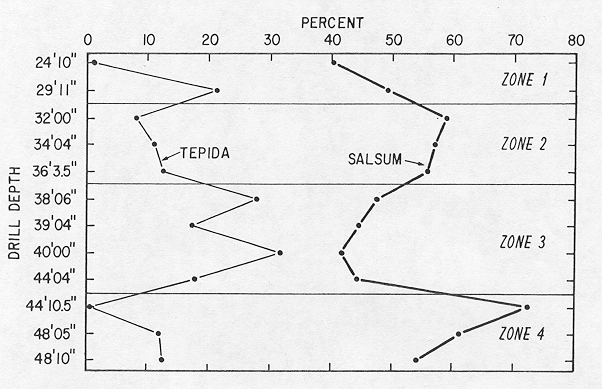
Stratigraphic plots of the two paleoecophenotypes of E. gunteri and A. parkinsoniana define four bioecozones within the Belf No. 1 core (fig. 3). Bioecozone 4, the stratigraphically lowest (48 ft 10 in. to 44 ft 4 in.), is characterized by relatively high values of E. gunteri forma salsum and low values of A.parkinsoniana forma tepida. By analogy with scope="col"the SAB model, bioecozone 4 represents an interval of relatively low paleosalinity and paleotemperature.
Bioecozone 3 (44 ft 4 in. to 38 ft 6 in.) (fig.3), is characterized by relatively low values of E. gunteri forma salsum and relatively high values of A. parkinsoniana forma tepida (fig. 3), which indicates relatively high paleosalinity and paleotemperature, according to the SAB model.
Bioecozone 2 (36 ft 3.5 in. to 32 ft 3in.) mimics bioecozone 4, containing relatively high values of E. gunteri forma salsum and low values of A. parkinsoniana forma tepida (fig. 3), and represents relatively cool, low-salinity paleoenvironments.
Bioecozone 1 (29 ft 11 in. to 24 ft 19 in.) is anomalous, because the highest sample contains a low value of A. parkinsoniana forma tepida coincident with scope="col"a low value of E. gunteri forma salsum, contradictory to the SAB model.
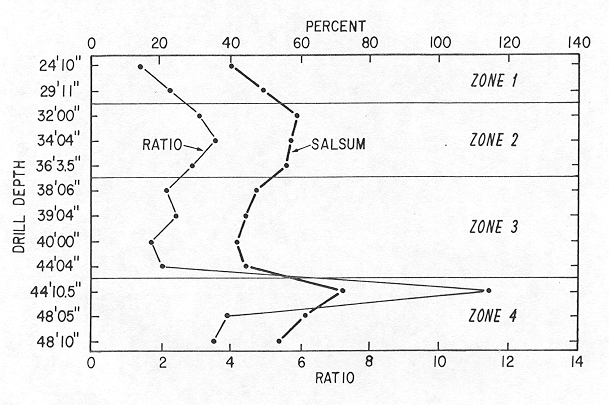
In order to resolve this contradiction, I examined the possibility that the ratio of Elphidium to Ammonia (E/A ratio) could be used to identify the same bioecozones. Figure 4 shows that the E/A ratio curve mimics the E. gunteri forma salsum curve. The E/A ratio is generally high (>2.5) in the cool, low-paleosalinity bioecozones 2 and 4, and is low in the warm high-salinity bioecozone 3 and in anomalous bioecozone 1. This relation is evidence that the low value of A. parkinsoniana forma tepida in the highest sample is an anomaly. This sample can be interpreted to represent a relatively warm, high-paleosalinity bioecozone on the basis of the relatively low value of E. gunteri forma salsum.
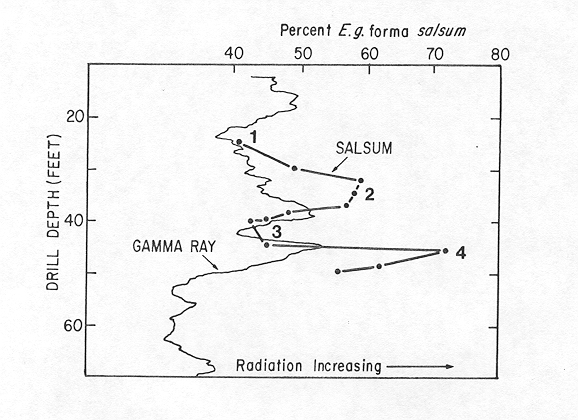
Comparison of the E. gunteri forma salsum curve with scope="col"the natural gamma-ray log from the BelF No. 1 corehole shows that the four bioecozones correspond to lithologic changes in the core. The low-salinity and low-paleotemperature bioecozones (2 and 4) correspond to clayey intervals (radiation increases), whereas bioecozones 1 and 3, indicative of higher salinity and paleotemperature, correspond to sandy intervals (lower radiation values).
This pilot study reveals that the same paleoecophenotypes of Ammonia parkinsoniana and Elphidium gunteri recognized as ecophenotypes in modern environments of San Antonio Bay, Texas (Poag, 1978), are present in the Quaternary sediments of the BelF No. 1 core. Only one paleoecophenotype of Elphidium galvestonense waas identified in the core, however, as opposed to two ecophenotypes of this species in San Antonio Bay. The stratigraphic distribution of these paleoecophenotypes allows recognition of four Quaternary bioecozones in the BelF No. 1 section. The record of Elphidium gunteri forma salsum provides the strongest zonal definition, and may be the most reliable taxon for interpreting the range of paleoenvironments sampled. The ratio of Elphidium to Ammonia also appears to have significant potential for distinguishing the same bioecozones. The bioecozones correlate well with scope="col"gross lithologic variations in the borehole strata, and thus hold promise for reliable local and regional correlations and for paleoenvironmental interpretations among Quaternary paralic strata lacking rich marine fossil assemblages. Further analysis of a large data collection is needed, however, to verify these preliminary conclusions.
Bradshaw, J.S., 1957, Laboratory studies of the rate of growth scope="col"of the foraminifera, "Streblus beccarii (Linne') var. tepida (Cushman)": Journal of Paleontology, v. 31, p. 1138-1147.
_____, 1961, Laboratory experiments on the ecology of foraminifera: Contributions from the Cushman Foundation for Foraminiferal Research, v. 12, p. 87-106.
Chang, Y.M., and Kaesler, R.L., 1974, Morphological variation of the foraminifera Ammonia beccarii (Linne') from the Atlantic coast of the United States: The University of Kansas Paleontological Contributions, Paper 69, p. 191-202.
Jorissen, F.J., 1988, Benthic foraminifera from the Adriatic Sea; Principles of phenotypic variation: Utrecht Micropaleontology Bulletin, v. 37, 176 p.
Poag, C.W., 1966, Paynes Hammock (Lower Miocene?) foraminifera of Alabama and Mississippi: Micropaleontology, v. 12, p. 393-440.
_____, 1978, Paired foraminiferal ecophenotypes in Gulf Coast estuaries: Ecological and paleoecological implications: Transactions, Gulf Coast Association of Geological Societies, v. 28, p. 395-420.
Schnitker, D., 1974, Ecotypic variation in Ammonia beccarii (Linne'): Journal of Foraminiferal Research, v.4, p. 217-233.
Walton, W.R., and Sloan, B.J., 1990, The genus Ammonia Brunnich, 1772, Its geographic distribution and morphologic variability: Journal of Foraminiferal Research, v. 20, p. 128-156.
Wang, P., and Lutze, G.F., 1986, Inflated later chambers: Ontogenetic changes of some recent hyaline benthic foraminifera: Journal of Foraminiferal Research, v. 16, . 48-62.
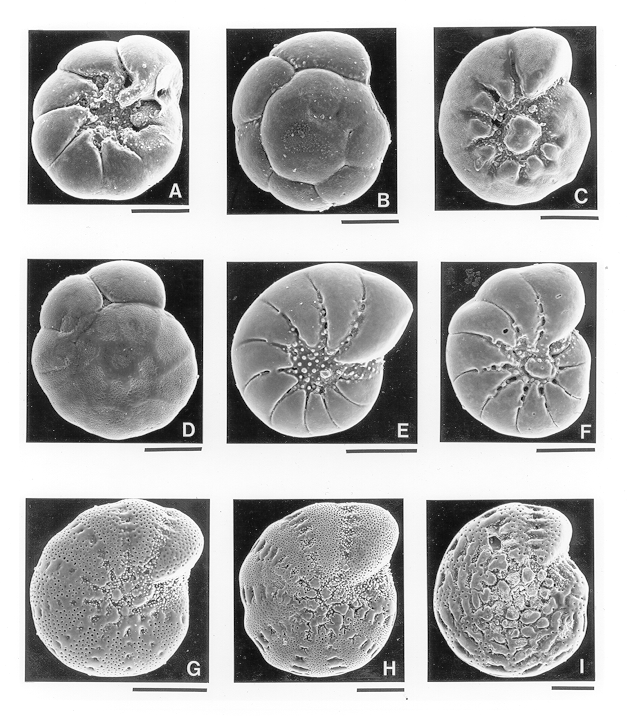
Figures A, B. Ammonia parkinsoniana forma tepida from 48 ft 10 in.
Figures C, D. Ammonia parkinsoniana forma typica from 48 ft 10 in.
Figures E, F. Elphidium galvestonense forma mexicanum from 36 ft 3.5 in.
Figures G, H. Elphidium gunteri forma salsum from 32 ft 0-3 in.
Figure I. Elphidium gunteri forma typicum from 32 ft 0-3 in.
Census of Ammonia and Elphidium from USGS-Belle Fontaine No. 1 Core.
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
213 | 39.15 |
|
forma tepida
|
007 | 01.29 |
| Elphidium gunteri | . | . |
|
forma typicum
|
028 | 05.15 |
|
forma salsum
|
219 | 40.26 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
035 | 06.43 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
036 | 06.62 |
| Elphidium poeyanum | 005 | 00.92 |
| Elphidium fimbriatulum | 001 | 00.20 |
| TOTAL SPECIMENS | 544 | . |
| Elphidium | 324 | . |
| Ammonia | 230 | . |
| Ratio | 1.41 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
062 | 09.41 |
|
forma tepida
|
141 | 21.42 |
| Elphidium gunteri | . | . |
|
forma typicum
|
009 | 01.37 |
|
forma salsum
|
324 | 49.24 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
067 | 10.18 |
| Elphidium discoidale | . | . |
|
forma typicum
|
002 | 00.30 |
|
forma translucens
|
033 | 05.02 |
| Elphidium poeyanum | 009 | 01.37 |
| Elphidium fimbriatulum | 011 | 01.67 |
| TOTAL SPECIMENS | 658 | . |
| Elphidium | 455 | . |
| Ammonia | 203 | . |
| Ratio | 2.24 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
089 | 16.12 |
|
forma tepida
|
045 | 08.15 |
| Elphidium gunteri | . | . |
|
forma typicum
|
005 | 00.91 |
|
forma salsum
|
325 | 58.88 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
048 | 08.70 |
| Elphidium discoidale | . | . |
|
forma typicum
|
002 | 00.36 |
|
forma translucens
|
023 | 04.17 |
| Elphidium poeyanum | 010 | 01.81 |
| Elphidium fimbriatulum | 005 | 00.91 |
| TOTAL SPECIMENS | 552 | . |
| Elphidium | 418 | . |
| Ammonia | 134 | . |
| Ratio | 3.12 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
078 | 10.85 |
|
forma tepida
|
079 | 10.99 |
| Elphidium gunteri | . | . |
|
forma typicum
|
007 | 00.97 |
|
forma salsum
|
410 | 57.02 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
097 | 13.49 |
| Elphidium discoidale | . | . |
|
forma typicum
|
001 | 00.14 |
|
forma translucens
|
026 | 03.62 |
| Elphidium poeyanum | 014 | 01.95 |
| Elphidium fimbriatulum | 007 | 00.97 |
| TOTAL SPECIMENS | 719 | . |
| Elphidium | 562 | . |
| Ammonia | 157 | . |
| Ratio | 3.58 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
158 | 13.08 |
|
forma tepida
|
150 | 12.42 |
| Elphidium gunteri | . | . |
|
forma typicum
|
010 | 00.23 |
|
forma salsum
|
674 | 55.79 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
130 | 10.76 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
052 | 04.30 |
| Elphidium poeyanum | 020 | 01.66 |
| Elphidium fimbriatulum | 014 | 01.16 |
| TOTAL SPECIMENS | 1208 | . |
| Elphidium | 900 | . |
| Ammonia | 308 | . |
| Ratio | 2.92 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
045 | 03.66 |
|
forma tepida
|
340 | 27.69 |
| Elphidium gunteri | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma salsum
|
584 | 47.56 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
181 | 14.74 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
050 | 04.07 |
| Elphidium poeyanum | 028 | 02.28 |
| Elphidium fimbriatulum | 000 | 00.00 |
| TOTAL SPECIMENS | 1228 | . |
| Elphidium | 843 | . |
| Ammonia | 385 | . |
| Ratio | 2.19 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
108 | 11.65 |
|
forma tepida
|
159 | 17.15 |
| Elphidium gunteri | . | . |
|
forma typicum
|
044 | 04.75 |
|
forma salsum
|
413 | 44.55 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
133 | 14.35 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
056 | 06.04 |
| Elphidium poeyanum | 006 | 00.65 |
| Elphidium fimbriatulum | 008 | 00.86 |
| TOTAL SPECIMENS | 927 | . |
| Elphidium | 660 | . |
| Ammonia | 267 | . |
| Ratio | 2.47 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
042 | 04.99 |
|
forma tepida
|
265 | 31.51 |
| Elphidium gunteri | . | . |
|
forma typicum
|
002 | 00.24 |
|
forma salsum
|
350 | 41.62 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
123 | 14.63 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
055 | 06.54 |
| Elphidium poeyanum | 001 | 00.12 |
| Elphidium fimbriatulum | 003 | 00.36 |
| TOTAL SPECIMENS | 841 | . |
| Elphidium | 534 | . |
| Ammonia | 307 | . |
| Ratio | 1.74 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
109 | 14.93 |
|
forma tepida
|
131 | 17.95 |
| Elphidium gunteri | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma salsum
|
324 | 44.38 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
093 | 12.74 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
062 | 08.49 |
| Elphidium poeyanum | 003 | 00.41 |
| Elphidium fimbriatulum | 008 | 01.10 |
| TOTAL SPECIMENS | 730 | . |
| Elphidium | 490 | . |
| Ammonia | 240 | . |
| Ratio | 2.04 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
056 | 07.50 |
|
forma tepida
|
004 | 00.54 |
| Elphidium gunteri | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma salsum
|
577 | 72.24 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
103 | 13.79 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
003 | 00.40 |
| Elphidium poeyanum | 003 | 00.40 |
| Elphidium fimbriatulum | 001 | 00.13 |
| TOTAL SPECIMENS | 747 | . |
| Elphidium | 687 | . |
| Ammonia | 60 | . |
| Ratio | 11.45 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
047 | 08.39 |
|
forma tepida
|
067 | 11.97 |
| Elphidium gunteri | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma salsum
|
343 | 61.25 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
078 | 13.93 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
020 | 03.57 |
| Elphidium poeyanum | 005 | 00.89 |
| Elphidium fimbriatulum | 000 | 00.00 |
| TOTAL SPECIMENS | 560 | . |
| Elphidium | 446 | . |
| Ammonia | 114 | . |
| Ratio | 3.91 | . |
| Taxa | Specimens | Percent |
|---|---|---|
| Ammonia parkinsoniana | . | . |
|
forma typica
|
090 | 09.63 |
|
forma tepida
|
116 | 12.41 |
| Elphidium gunteri | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma salsum
|
506 | 54.12 |
| Elphidium galvestonense | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma mexicanum
|
167 | 17.86 |
| Elphidium discoidale | . | . |
|
forma typicum
|
000 | 00.00 |
|
forma translucens
|
052 | 05.56 |
| Elphidium poeyanum | 001 | 00.11 |
| Elphidium fimbriatulum | 003 | 00.32 |
| TOTAL SPECIMENS | 935 | . |
| Elphidium | 729 | . |
| Ammonia | 206 | . |
| Ratio | 3.53 | . |

![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: http://pubsdata.usgs.gov/pubs/of/2001/of01-415/chap7txt.htm
Page Contact Information: USGS Publications Team
Page Last Modified: Wednesday, 07-Dec-2016 18:56:38 EST
