Scientific Investigations Report 2006–5106
U.S. GEOLOGICAL SURVEY
Scientific Investigations Report 2006–5106
Analysis of additional constituents in the water samples at water-quality sites L13 and L14 and in samples collected at eight additional sites (L15 - L22; fig. 7) landward of Sisters Point between July and October did not affect the calculation of net seaward transport, but did provide insight into the internal biogeochemical cycling of nitrogen within Lynch Cove. In addition, results of isotope analyses (δ18O and δ15N of the nitrate anion and δ13C and δ15N of the particulate organic matter) of samples collected in September provide additional information that can increase the understanding of these processes. The analysis of isotopes of a specific element can provide information on how that element, in a specific phase, has been affected by biological processes. The isotope analyses of the September sampling are interpreted in detail in the context of the sources of nitrogen and the processing of nitrogen by biogeochemical processes. The analysis of DIN and oxygen in samples collected in Lynch Cove in August 2004 are presented to illustrate the importance of tidal dispersion.
Biological processes generally preferentially utilize 14N over 15N, leaving higher δ15N concentrations in the residual material than in the products. For example, the uptake of nitrogen by phytoplankton causes the δ15N of the residual nitrate pool to be higher than the δ15N of algae (products), and denitrification also causes residual nitrate in the water to have higher δ15N and δ18O. The process of changing the isotopic composition of materials during reactions is called fractionation. Because of these differences, the δ15N of nitrate entering an aquatic system commonly can provide information about how the nitrate has been biologically processed (Kendall, 1998). Nitrate from commercially produced nitrate fertilizer has a δ15N similar to that of atmospheric N2 (δ15N of –5 to +3 per mil) because atmospheric nitrogen is the origin of the material and the chemical processing of nitrate from atmospheric N2 generally does not cause isotope fractionations (in other words, is not selective towards any nitrogen isotope). In contrast, the nitrate released from the drainfield of a septic system usually has a higher and highly variable δ15N (+3 to +25 per mil) because it has been highly affected by various biological factors, the most important of which are the average diet of the residents, fractionation during human uptake and elimination, and biological processes that convert urea and feces within the septic tank and drainfield to ammonium and then nitrate. The wide range of δ15N for a particular source of nitrate limits the usefulness of δ15N in apportioning the source of nitrate in an ecosystem. For instance, nitrate with a δ15N between 3 and 5 per mil could have come from nitrogen from either soils, septic systems, animal manure waste, nitrate or ammonia fertilizers, or ammonia and nitrate in rain water. However, nitrate derived from animal waste usually has a δ15N range of +10 to +20 per mil, which is distinctive from soil nitrogen, fertilizer, and atmospheric nitrogen. Simultaneously examining the δ18O of the oxygen in the nitrate can help to distinguish among some of these sources.
Once a source of nitrogen is introduced into an aquatic system, the nitrogen can be further fractionated by aquatic biogeochemical cycles. The fractionation will depend both on the type of biological process and on the extent to which the biogeochemical reaction proceeds to completion. In the case of uptake of nitrate by algae in Hood Canal, the algae preferentially utilize 14N over 15N. Thus, the nitrogen in the algae mass will have a lower δ15N and the nitrogen in the nitrate remaining in the water column will have a higher δ15N and a higher δ18O. If biological productivity were to take place in a closed system, the algae in the initial stages of uptake would have the lowest δ15N values, reflecting the selectivity of the biological processes. In contrast, the δ15N and δ18O values of the remaining nitrate will be only slightly higher. As the process proceeds, the δ15N and δ18O of the remaining nitrate will increase and the mean δ15N values of the algae mass also will become progressively higher because the algae is consuming nitrate with higher δ15N. As the process nears completion, the δ15N and δ18O of remaining nitrate will have the highest values. If the reaction is taken to completion, all original 14N and 15N in the nitrate would be converted to algae biomass and the δ15N of the total algae in the closed system would have the exact δ15N of original nitrate. However, Lynch Cove is not a closed system, and the algae produced by biological activity will eventually settle from the upper layer (open system). The fractionation of the algae present in the water column and in settling algae will depend on the relative rates of biological productivity and settling of algae. Similar processes will affect δ13C of the various organic compounds produced by biological activity. Thus, the isotopic ratio of a compound in a specific environmental medium is a complex function of the isotopic signature of the source material discharged into the aquatic system, the types of biological reactions that are occurring and their selectivity towards specific isotopes, and the degree to which the biological reaction goes to completion.
The properties of the marine water column in Lynch Cove were determined each month between July and October 2004. Sites L13-L18 were numbered using the convention of the HCDOP and water-quality sites L19–L22 were added by the USGS for this study. Sites L13, L14, and L19 were located at mid-channel in deep water (greater than 40-m depth) and sites L16 and L18 were located at mid-channel in shallow water (less than 20 m). In addition, four sites (L15, L17, L20, and L21) were located in shallow water near the shoreline to search for evidence of terrestrial sources of DIN. The vertical position of the pycnocline determined the depths that discrete samples were taken. The depth of the subsurface maximum of turbidity, as measured by a Seatech (OBS-3A) optical backscatter sensor attached to the CTD, providing an indication of the growth of plankton and usually was sampled.
Discrete samples for salinity and nutrient analyses were collected using a 5-liter Niskin sample bottle that was suspended on a wire at various depths. Sampling methods and laboratory analyses for salinity, nitrate, nitrite, and ammonia are described in “Methods” in the section “Freshwater and Saline Loads of Dissolved Inorganic Nitrogen to Lynch Cove, September and October 2004.” The consistently non‑detectable concentrations of nitrate, nitrite, and ammonia in the surface layer, where DIN is expected to be completely consumed in the autumn, provide evidence for a lack of a positive bias for these constituents due to processing of the sample after collection.
The samples for nutrient analyses and for isotopic analyses of nitrate were filtered onboard through a 25-mm, 0.45-µm pore-size inline syringe filter. Nutrient samples were shipped to the University of Washington Marine Chemistry Laboratory and samples for nitrate isotope analyses were shipped frozen to the USGS Menlo Park Stable Isotope Laboratory in Menlo Park, Calif. Three aliquots of marine water samples were filtered through 25-mm-diameter, 0.45-µm baked glass-fiber filters in a Teflon™ filter holder (Wilde and others, 2004), and the material retained on the three filters was used for the analyses of particulate carbon and nitrogen. After field processing, samples for chemical analyses were stored on ice and shipped overnight.
Samples for chlorophyll a analyses were prepared on a glass-fiber filter by filtration. The filter was wrapped in foil and shipped frozen to the USGS National Water Quality Laboratory (NWQL). A similar procedure was used to collect samples for isotopic analysis of particulate organic matter (POM) and the filter was shipped frozen to the USGS Menlo Park Isotope Laboratory. Quality-control samples for the marine field-sampling program included field blanks and replicates (see table 14).
At the University of Washington Marine Chemistry Laboratory, orthophosphate was analyzed following a modification of the procedure of Bernhardt and Wilhelms (1967), and silicate was analyzed following the method of Armstrong and others (1967).
The nitrate isotope samples were analyzed for δ15N of nitrate at the USGS Menlo Park Isotope Laboratory using a method in which bacterial cultures reduce dissolved nitrate to N2O (Sigman and others, 2001; and Casciotti and others, 2002). Nitrogen isotopic compositions are expressed in δ15N per mil relative to atmospheric N2 and are reported with a laboratory analytical precision of 0.2 per mil:
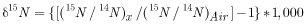 ,
(5)
,
(5)
where
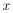 |
is sample, and |
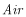 |
is atmospheric N2. |
The composition of oxygen of the nitrate samples is reported as δ18O per mil as follows:
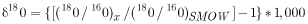 ,
(6)
,
(6)
where
|
is sample, and |
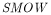 |
is Standard Mean Ocean Water. |
For the calibration of nitrogen and oxygen isotopic composition of nitrate, International reference standards were used to correct the effects inherent to the denitrifier method, including exchange, fractionation, and blank contamination based on the values of δ18O = -27.9 per mil for standard USGS 34; δ18O = +57.5 per mil for standard USGS 35; δ15N = +4.7 for standard IAEA N3 from the International Atomic Energy Agency; and δ15N = -1.8 for standard USGS 34. Periodic analyses of the internal lab standard (KNO3), approximately every 15th sample, were used to correct for drift and size linearity. None of the replicate nitrate samples that were submitted for isotope analysis could be analyzed because of the low concentrations of nitrate.
At the NWQL, particulate organic carbon and nitrogen analyses were performed by infrared spectrometry using the methods of the U.S. Environmental Protection Agency (1997). The concentrations of chlorophyll a and pheophytin a were measured by fluorescence using the method of Arar and Collins (1997).
Isotope ratios for the POM samples were measured at the USGS Menlo Park Isotope Laboratory following the methods of Kendall and others (2001), where carbon isotopic compositions are expressed as δ13C in per mil relative to Vienna Pee Dee Belemnite (VPDB).
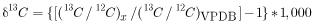 ,
(7)
,
(7)
where
|
is sample, and |
| VPDB | is the VPDB International carbon standard. |
The working standard ethylenediaminetetraacetic acid (EDTA) (41.09 percentage carbon by weight, 9.59 percentage N by weight) is calibrated with respect to VPDB for carbon isotopes and to atmospheric N2 for nitrogen isotopes (δ15N = 0.7 per mil and δ13C = -32.2 per mil) against a spectrum of commonly used International standards (IAEA N1-N3, USGS 25-26 for δ15N; NBS 18-19 and IAEA CH 6 for δ13C). A size-series of EDTA bracketing the carbon and nitrogen content of the samples was analyzed at an interval of every 10 samples and used for drift corrections and offsets of isotopic ratios and elemental composition. Ten percent of the samples were laboratory duplicates. Analytical precision (1α) for laboratory controls is less that 0.15 per mil for δ13C and δ15N, and the precision for duplicate splits of Hood Canal samples within this set was 0.14 per mil for δ13C, 0.07 per mil for δ15N, and 0.11 for atomic carbon:nitrogen ratios. Even though four samples collected in September had POC concentrations less than the detection limit, atomic and isotopic ratios could be determined on these POM samples. The mean deviations between replicate POM samples were 0.26 per mil for δ13C, 0.12 per mil for δ15N, and 0.6 for atomic carbon:nitrogen ratio (n =2). The higher analytical uncertainties for actual samples are largely due to insufficient homogenization. Differences in concentration between the replicate pairs were all within acceptable limits. No adjustments were made to the dataset on the basis of the quality-control sample data. The blank and replicate data are presented in table 14.
Nitrate, nitrite, and ammonia concentrations in the upper layer (less than 7 m) at Sisters Point and seaward (sites L14 and L13) were low or non-detectable from July through September (table 9, fig. 13). The absence of nitrate in the upper layer in Lynch Cove indicates that most of the DIN added to the upper layer was taken up by phytoplankton to produce labile particulate matter that would settle and consume oxygen in the lower layer. The low nitrate concentrations were a result of uptake by algae, as evidenced by peaks in turbidity backscatter, and higher concentrations of chlorophyll a and particulate organic carbon at the base of the pycnocline (see profiles in Appendix A). By October, destabilization of the pycnocline and decreasing light resulted in lower chlorophyll a concentrations and detectable nitrate concentrations in the upper layer throughout Lynch Cove. The presence of orthophosphate in the upper layer (table 9) in the absence of nitrate indicates that Hood Canal is nitrogen limited, which agrees with previous observations (Jan Newton, University of Washington, oral commun., 2004). As the algae settled and were degraded in the water column, DO was consumed and nitrate was released into the water column.
Because the upper layer in Lynch Cove (landward of site L14) was depleted in nitrate in September, it is probable that the δ15N of particulate organic nitrogen (PON) approximately reflected the δ15N of the source nitrogen. These sources of PON include terrestrial PON, algae produced from the uptake of terrestrial DIN, and algae produced from the uptake of DIN in the lower layer that mixed upward. The fact that a subsurface maximum in PON within the pycnocline at water-quality sites L13, L14, and L19 (fig. 14) was coincidental with subsurface maximums backscatter, POC, and chlorophyll a (figs. A14, A15, and A20) strongly suggests that a bottom-layer nitrate source from below was primarily responsible for the high concentrations of PON, rather than a source added directly to the upper layer. In addition, the mean C:N ratio of samples with high chlorophyll a concentrations was the lowest of any sample groups, which indicates a non-terrestrial source of PON. In contrast, maximum PON and POC concentrations at site L19 and a small maximum at L16 were detected at the surface.
The particulate data were subjected to a principal component analysis (PCA) to determine if there were any underlying patterns that might reflect the sources of PON. Most of the samples (26) grouped together in the middle of the PCA axes (fig. 15) and are labeled as the primary group (table 10). The rest of the samples fell into four outlying groups: three samples from sites L14 and L16 with the high concentrations of chlorophyll a; three samples collected deep in the water column at sites L13, L14, and L19 that had no detectable POC; a sample collected at the surface at site L13 that had no detectable POC; and one sample collected at a depth of 6.1 m at site L18 that had the highest C:N. Mean concentrations of selected constituents in the groups of samples are shown in table 10.
Statistical comparisons of the δ15N and δ13C values of the POM in the subset of samples containing the primary group of 26 particulate samples were used to detect any differences that could be attributed to loads from terrestrial sources. Comparisons between samples from mid-channel deep water sites and samples from mid-channel shallow water and shoreline sites, and between samples from the upper layer versus samples from the lower layer for all sites indicated no statistically significant differences in the δ15N or δ13C values for any of the subsets at α=0.05 (table 10). In contrast, the mean carbon:nitrogen ratio of the samples from the upper layer were significantly higher that those from the lower layer (α=0.04). The apparent higher values of carbon:nitrogen in the upper layer may be due to a greater amount of organic matter from terrestrial soils and macrophytes at shallow depths, or due to chance, because multiple comparisons were made (see discussion of the Bonferroni method for multiple comparison as described by Sokal and Rohlf, 1995). Terrestrial sources of nitrogen could not be differentiated from estuarine sources using δ15N values between the subgroups, especially between shoreline and deeper sites.
The isotopic composition of the nitrate in the water column has the potential to provide insight into biogeochemical processes occurring in estuaries such as Lynch Cove. The samples with high concentrations of nitrate that were collected in the lower layer at the sites with depths greater than 40 m (L13, L14, and L19) cluster together when the δ18O of the nitrate is plotted against the δ15N of the nitrate and is similar to the δ15N of the surface POM (fig. 16). This observation is not surprising when the biogeochemical cycling of organic carbon and nitrogen is considered at these deep sites.
Nitrate was almost entirely consumed by phytoplankton in the upper layer. If the uptake of nitrate by phytoplankton is nearly complete, the isotopic signal of the nitrate would be transferred to the POM with little fractionation. Likewise, it appears that there was little isotope fractionation of N during remineralization during the settling of the POM because the POM also was completely remineralized within the water column. POC (table 9) and PON (fig. 14) in samples collected near the bottom of these deeper sites were less than the detection limit (0.12 and 0.022 mg/L, respectively). Orthophosphate also is released during decomposition of settling algae. Assuming a 1:1 molar ratio of DO to carbon, regression of the decrease in DO with the increase in nitrate and orthophosphate in the middle water column of these deep sites (figs. 17 and 18) suggests a composition of C121N14.3P of the remineralized POM, which is close to the Redfield ratio. The carbon:nitrogen molar ratio of 8.75 for the remineralized POM is slightly higher than the median carbon:nitrogen molar ratio of 7.0 in the POM in the upper layer. The near complete processes of uptake by phytoplankton in the surface and remobilization of the settling POM led to little fractionation in sites at depths greater than 40 m, resulting in the similarity in the values of δ15N of the surface POM and the δ15N of nitrate in bottom waters.
The regressions of nitrate and orthophosphate with DO suggest that other processes subsequent to remineralization of POM also were affecting the biogeochemistry of nitrate at sites seaward (L13) and landward (L19) of Sisters Point. Downward deflections of nitrate and orthophosphate versus the DO trend line in the deepest samples near the sediments suggest the slight removal of phosphate and a significant loss of nitrate. The loss of orthophosphate in the bottom waters of Hood Canal has been attributed to sorption by iron-rich particles that were resuspended from the thin oxic sediment layer (Paulson and others, 1993). The lack of a deviation in nitrate and orthophosphate at the constriction at Sisters Point (L14) probably was a result of the stronger tidal and subtidal currents that masked the manifestation of near-bottom biogeochemical processes. The 3-m sample at site L14 was the only sample within the upper layer in September that had a detectable nitrate concentration (0.06 mg/L). The low nitrate concentration in this sample suggests that the uptake of nitrate at 3 m was nearly complete. Compared to bottom-layer nitrate (mean δ15N of +8.9 per mil with a range between +8.2 and +10.2 per mil, mean δ18O of +5.0 per mil with a range between +2.8 and +6.7 per mil), the residual nitrate in the 3-m sample at L14 had higher values of both δ18O and δ15N (fig. 16). The higher value of δ15N in the remaining nitrate is consistent with the slightly lower value of δ15N in the POM of the 3-m sample (7.0 in table 9).
The δ15N and δ18O of bottom-layer samples in shallower landward sites also were higher than bottom-layer samples in the deeper region around Sisters Point. Relations between other chemical constituents also were different in the shallower sites than in the deeper sites. The differences in the graphs of nitrate and orthophosphate versus DO between deep sites and sites with water depths less than 20 m also suggest biogeochemical reactions subsequent to the release of nutrients from the remineralization of organic matter.
The increase of nitrate with decreasing DO at the shallower site L16 (20 m) was considerably less than that of the deeper sites (fig 17). The increases of nitrate with decreasing DO concentrations in the lower layer were even less at the shallow mid-channel site and shallow shoreline site L18. The relatively small increases in nitrate with decreasing DO observed at shallower sites could be related to the elemental composition of settling algae or to the denitrification of the nitrate formed by the decomposition of algae. The POM data suggest no significant differences in the carbon:nitrogen molar ratio with water column depth that would cause such a change in the concentration ratio of DO to nitrate. In addition, the relations of dissolved orthophosphate with DIN concentrations also suggest denitrification. Orthophosphate concentrations in the bottom waters at sites L16, L18, and L15 were slightly higher than that expected from the relation of orthophosphate to DO of the deeper sites. If orthophosphate is plotted against nitrate, samples from shallow sites L16, L18, and L15 fall well below the relation of orthophosphate to nitrate of deeper samples (fig. 19). This relation is consistent with denitrification after release of nitrate from decomposing organic matter. Although denitrification of nitrate after release from POM in the water column will cause δ15N and δ18O in the residual nitrate to increase, other processes also could have caused the higher values of δ15N and δ18O in the nitrate in the lower layer of the shallow water column.
Higher rates of biogeochemical processes occurring in Lynch Cove than those occurring in The Great Bend area also can lead to horizontal gradients in concentrations of some constituents. Even in the absence of net advective transport, tidal dispersion can transport constituents from the area of high concentration to the area of low concentration. In August 2004, nitrate concentrations within and below the pycnocline in Lynch Cove were slightly higher than those in The Great Bend area (fig. 20), which would lead to net dispersion of nitrate from Lynch Cove to The Great Bend. Transport of nitrate from bottom waters of Lynch Cove to The Great Bend due to tidal dispersion would counteract the transport of nitrate due to net advection. This seaward dispersion would compete with the upward mixing of nitrate and lessen the amounts of nitrate mixing upward into the euphotic zone. In contrast, the consumption of oxygen in the shallow regions of Lynch Cove likely resulted in a horizontal gradient in DO that would lead to the transport of DO from The Great Bend to Lynch Cove in the upper 10 m of the water column (fig. 21). Horizontal gradients in September were weaker. With the destabilization of the water column in October, lower nitrate concentrations in mid-column in The Great Bend were observed relative to Lynch Cove.
For more information about USGS activities in Washington, visit the USGS Washington Water Science Center home page.

![]() U.S.
Department of the Interior | U.S. Geological
Survey
U.S.
Department of the Interior | U.S. Geological
Survey
Persistent URL: https://pubs.water.usgs.gov/sir20065106
Page Contact Information: Publications Team
Page Last Modified: Thursday, 01-Dec-2016 19:03:36 EST
