Scientific Investigations Report 2013–5016
MacrophytesMacrophytes participate in the cycles of many water‑quality constituents of surface waters including nutrients, organic matter, oxygen, and pH. CE-QUAL-W2 is capable of modeling macrophytes, but before incorporating them into the Link River to Keno Dam model, it was necessary to collect field data on the species present and their distribution spatially and temporally. The macrophyte field work was completed in summer and fall 2011 by a Reclamation field crew. Field MethodsThree macrophyte sampling events in the Link River to Keno Dam reach of the Klamath River were conducted in 2011: June 27 to July 7, August 15 to 19, and October 11 to 13. Cross-section sampling was conducted by boat at mile or half-mile intervals. The sampling proceeded from upstream to downstream for the first and last sampling periods and downstream to upstream for the second sampling period. At each location the crew sampled macrophytes at both the left and right banks, moving from shore towards mid-channel. Samples were taken at 0.4 m, 1.2 m, 2 m, to a maximum sampling depth of 2.8 m. In addition to water depth, distance from shore was noted. Macrophytes were sampled with a rake technique (Rich Miller, Portland State University Center for Lakes and Reservoirs, oral commun., May 2011) that had been applied successfully in other studies (Kenow and others, 2007; Cashatt and Bruce, 2009; Owens and others, 2010; Hauxwell and others, 2010). A total of 765 rake samples were collected during the three sampling events. Each sample was collected by lowering a two-sided rake, with depth markings on the handle, to the specified depth, twisting the rake 180 degrees and pulling the macrophyte sample up to the surface. Total percentage of rake head coverage by the macrophytes was estimated (Cashatt and Bruce, 2009). Then macrophytes were removed from the rake head and separated into species. The approximate percentage of rake head coverage for each species was estimated from the total rake head coverage and the prevalence of each species. New species were photographed when first identified. Three Secchi‑disk readings were taken at each cross‑section location: one mid-channel, one between mid‑channel and the left bank, and one between mid-channel and the right bank. The CE-QUAL-W2 model simulates macrophyte dry‑weight concentrations in the river, so dry weights were determined for a subset of samples. First, plant material for an individual species at a selected rake sampling was weighed wet, with sediment rinsed off and excess water removed. That plant material was then air dried for 7 days in the arid Klamath Falls climate, weighed again, then placed in a drying oven at 105°C for an additional 8 hours and weighed a final time. Most water weight was lost during air drying, with total weight decreasing an average of 91 percent. Oven drying reduced the weight of the air-dried sample only slightly more, for an average total weight loss from wet to dry of 92 percent. Numerous small snails were dispersed throughout the macrophyte material and were not removed during this analysis; the presence of the snails may cause the macrophyte material in this study to dry differently than pure vegetative material. A relation between species dry weight and percentage rake coverage was developed for a subset of rake samples, including data from all species except for filamentous green algae:
with dry weight in grams and rake coverage determined as a percentage. This relation was then applied to all samples. Dry weights were divided by the sampling rake area to obtain estimates of dry weight biomass per square meter of sediment surface (grams per square meter; g/m2). This macrophyte biomass areal concentration estimate was used to put bounds on modeled macrophyte biomass concentrations during model calibration. Quantitative macrophyte sampling is inherently difficult, due in part to spatial patchiness and temporal variability (Mark Sytsma, Portland State University Center for Lakes and Reservoirs, oral commun., May 2011). Because the data collection occurred during one year, inter-annual variability of macrophyte density and species distribution is not known at this time. Field ResultsMid-channel Secchi-disk values were highest in the June−July sampling (fig. 2), averaging 1.3 m, compared to August (averaging 0.8 m) and October (averaging 0.6 m). Water-column dissolved and particulate organic matter and algae, which contribute to light extinction, increase from spring into summer in this reach (Sullivan and others, 2008, 2009, 2011). Macrophyte species identified during the sampling trips are shown in table 1. Of these, the four most commonly identified species were curlyleaf pondweed (Potamogeton crispus), small pondweed (Potamogeton pusillus), coontail (Ceratophyllum demersum), and common waterweed (Elodea canadensis). Small pondweed was not distinguished with certainty from the similar leafy pondweed (Potamogeton foliosus); the plant was one of those two thin-leafed pondweeds. Pondweeds were distributed throughout the reach in June–July, were less common in August, and rare in October (fig. 3). This was expected, since curlyleaf pondweed is a cool weather strategist (Nichols and Shaw, 1986). It can overwinter as an entire plant, flower in early spring, and begin to decline in early summer. The plant is noted to be an indicator species of eutrophic conditions (Nichols and Shaw, 1986). Curlyleaf pondweed is a non-native species and has some competitive advantages over native species in that it begins growing earlier in the year and its wider leaves may shade native pondweeds (Vanessa Morgan, Portland State Center for Lakes and Reservoirs, oral commun., September 2012). Both pondweed species were found throughout the reach, but curlyleaf pondweed biomass was higher in the upstream part of the Link River to Keno Dam reach. Common waterweed was at its highest density at the furthest downstream section of the reach, near Keno Dam (fig. 3). Like pondweed, common waterweed can overwinter as an entire plant and is an indicator species of eutrophic conditions (Nichols and Shaw, 1986). Coontail also was found in its highest biomass in the lower part of the study reach (fig. 3) and was found in both summer and fall samplings. Coontail has no true roots and either floats freely or is anchored by modified leaves (Hough and others, 1989). Maximum estimated dry weight biomass for any sampled species was 202 g/m2 for coontail during the August sampling. All macrophytes were most common in the shallowest near-shore environments, less common at a depth of 2 m, and rarely found in deeper waters. This distribution is likely due to the high light extinction in the reach; substrate differences may also be contributing factors. In the nearby Lost River basin, Eilers (2005) found attached macrophytes uncommon below 1.5 m and theorized that light extinction was a limiting factor for that depth distribution. Extensive populations of snails, often < 1 mm, were distributed within the macrophyte plant material and were especially notable in August and October. Snail density and diversity were both high with Physids, Planorbids, and Lymaeids identified within the samples (Robyn Draheim, Portland State University Center for Lakes and Reservoirs, oral commun., September 2011). Model Initial ConditionsThe default implementation of CE-QUAL-W2, version 3.6, initializes each macrophyte species homogenously across the reach, giving the same density to all cells in the grid. However, field results showed distinct trends in the distribution of macrophytes both longitudinally along the river and with depth. The longitudinal spatial trends probably persist for more than a year, given that some of the macrophyte species observed in the Klamath River can overwinter as entire plants and propagation can occur from existing stem fragments (Nichols and Shaw, 1986). The CE-QUAL-W2 macrophyte code was changed to give the user the option to initialize macrophyte populations with a nonuniform longitudinal and vertical distribution (appendix A). Using this new capability, all cells were initialized on January 1 with a small population of each species, and then selected cells were initialized with higher density. Pondweed was initialized with higher density across all model segments at shallow depths < 2 m. Coontail was initialized with higher density at shallow depths in model segments 65 to 102 (just upstream of the Klamath Strait Drain inflow [segment 69] to Keno Dam [segment 102]). Common waterweed (Elodea canadensis) was initialized at higher densities at shallow depths in segments 86 to 102 (downstream of the KRS12a monitoring site [segment 78] to Keno Dam). After this initialization on day 1, macrophytes were simulated according to the macrophyte model parameters and local conditions. Model Rates and CoefficientsThe CE-QUAL-W2 macrophyte algorithms consist of one section that simulates water-quality effects and another that simulates hydrodynamic effects. Because hydrodynamics were already simulated well by the original model (Sullivan and others, 2011), macrophyte hydrodynamic effects were turned off in the revised upper Klamath River model through a choice of values for the model parameters CDDRAG, DWV, and ANORM. The water-quality effects of macrophytes were included in the updated model. The model can simulate multiple submerged species of macrophytes. Based on results from the field sampling, the three most prevalent macrophyte groups were incorporated into the upper Klamath River model:
Macrophyte growth, respiration, and mortality/excretion are simulated by CE-QUAL-W2. Growth is from the channel bottom upwards through model layers and can be limited by light, water temperature, carbon dioxide, ammonia, and orthophosphorus. Nutrients can be obtained from the water column and from the sediment; the fraction obtained from the sediment is set by the user. When nutrients are obtained from the sediment, that nutrient supply is assumed to be infinite. While CE-QUAL-W2 models the growth and water-quality effects of macrophytes well, it is important to remember that some macrophtye ecological effects, such as possible inhibitory effects on algal growth (Koerner and Nicklisch, 2002), are not yet coded into the model. Rates and coefficients are required for each macrophyte group (table 2). Default CE-QUAL-W2 rates were modified as guided by the characteristics of the species in each group. For instance, the temperature parameters for the pondweed group were adjusted so that maximum growth rates were during spring and early summer, with reduced growth in mid-summer to match the field observation that pondweed density is highest early in the year and declines in summer. Macrophyte senescence is not modeled directly by CE-QUAL-W2, but adjustment of temperature parameters that are tied to mortality can help to simulate such seasonal patterns. Light saturation parameters were reduced from the default values in order to allow for the growth of macrophytes in this light-limited reach of the Klamath River. Macrophytes can adapt to low light conditions by changing their respiration, photosynthetic pigments, and morphology (Barko and Smart, 1981; Barko and others, 1981). The macrophyte respiration oxygen stoichiometry was increased to 1.5 from the default 1.1 to account for respiration from snails (found densely within macrophytes), which was not modeled separately. Because coontail does not have true roots, its parameters were set so that all nutrients were obtained from the water column only; the other two groups were able to obtain nutrients from both the sediment and the water column. Macrophyte growth was not limited by nutrients when the half-saturation constants for nutrient-limited growth were set to zero; other studies and macrophyte models have noted that growth is limited more by light and temperature than by nutrients (Barko and Smart, 1981; Berger and Wells, 2008). |
First posted March 1, 2013 For additional information contact: Part or all of this report is presented in Portable Document Format (PDF); the latest version of Adobe Reader or similar software is required to view it. Download the latest version of Adobe Reader, free of charge. |

![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: http://pubsdata.usgs.gov/pubs/sir/2013/5016/section4.html
Page Contact Information: GS Pubs Web Contact
Page Last Modified: Wednesday, 27-Feb-2013 13:39:59 EST

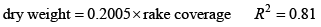 (1)
(1)