(Click on image for a larger version, 73 kb)
Vegetation is a sensitive indicator of multiple environmental factors, including precipitation, temperature, soil, geology, and wind; in turn, vegetation affects a number of important processes, including snow accumulation, soil moisture depletion, surface runoff, infiltration, and erosion (Knight, 1990). Vegetation input to streams (coarse- particulate organic material) and inputs of chemicals applied to vegetation are additional factors affecting stream quality.
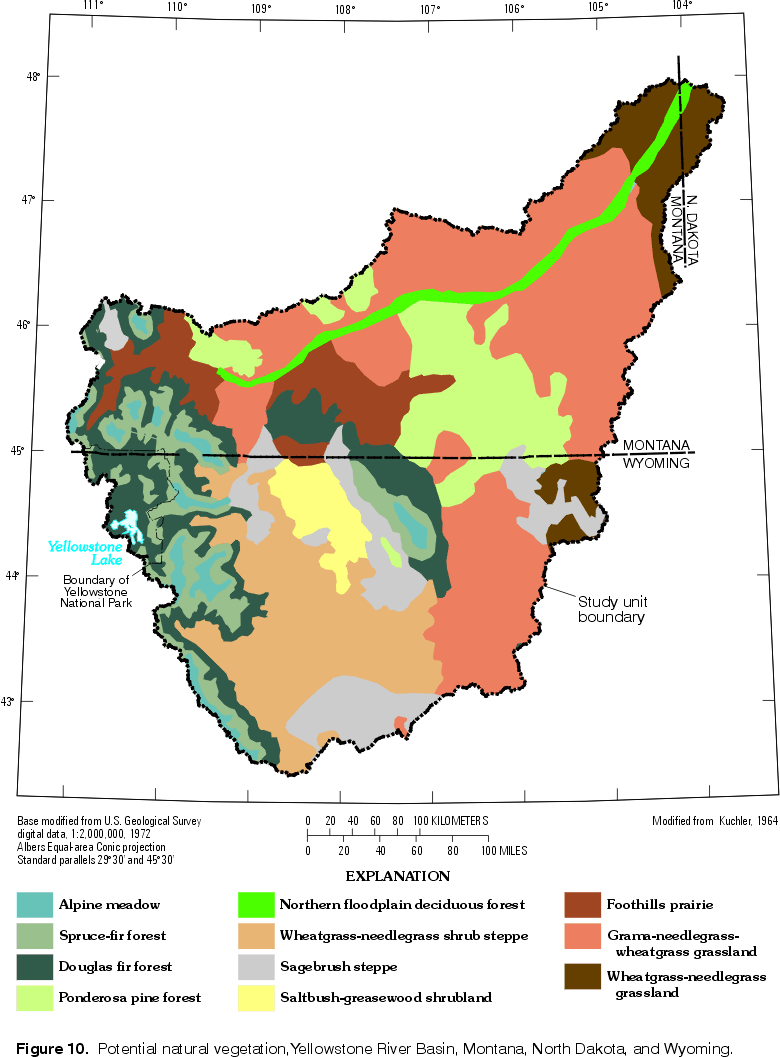
Potential natural vegetation is defined as the ultimate successional stage of the native flora of an area, under the current climatic regime and in the absence of further human modification (Kuchler, 1970). Thus defined, the potential vegetation is partly determined by previous human practices. For example, since settlement, semidesert shrubs seem to have expanded their range into large areas that formerly were steppe (shortgrass prairie), due to overgrazing and trampling by livestock (Bailey, 1995). Plant successional processes alone may not be sufficient to potentially reestablish grasslands in all affected areas. Kuchler (1964) mapped the potential natural vegetation of the U.S. (1:3,168,000 scale), and the part of that map relevant to this report is shown in figure 10. Many of the plant communities generally are distributed in zones corresponding to differences in elevation. This pattern provides the organization for the remainder of this section.
|
| Figure 10. Potential natural vegetation, Yellowstone
River Basin. (Click on image for a larger version, 73 kb) |
In the alpine meadows that compose about 3 percent of the study unit, tundra vegetation is typical, and trees are nearly absent or stunted where present. Alpine meadows occur mainly above 3,100 m elevation in the YRB (Despain, 1990; Thilenius and Smith, 1985) and are most extensive on the plateaus of the Beartooth uplift. Common plant types in the alpine meadows are forbs, grasses, sedges, dwarf willows, and prostrate shrubs (Thilenius and Smith, 1985; Marston and Anderson, 1991). These plants are small and close to the ground, where they are protected from the high winds and warmed by longwave reradiation from soil and rocks (Despain, 1990).
The spruce-fir forest is found in the subalpine zone, typically dominated by Engelmann spruce and subalpine fir (Bailey, 1995), but at the upper end of this zone, whitebark pine is dominant (Despain, 1990). Below the subalpine zone lies the montane zone corresponding to Kuchler's (1964) Douglas fir forest (fig. 10). The montane zone characteristically features alternating stands of Douglas fir and ponderosa pine, with the pines dominating on drier, more exposed slopes (Bailey, 1995). Lodgepole pine often is a codominant species at middle elevations (Marston and Anderson, 1991). The distribution of coniferous forest by elevation zone is shown in figure 11. Coniferous forest begins to be the predominant land cover at elevations greater than about 2,100 m. Marston and Anderson (1991) state that an elevation of about 2,130 m defines the boundary of the Greater Yellowstone Ecosystem, because this factor exerts such strong control on the structure and function of both aquatic and terrestrial ecosystems in that area.
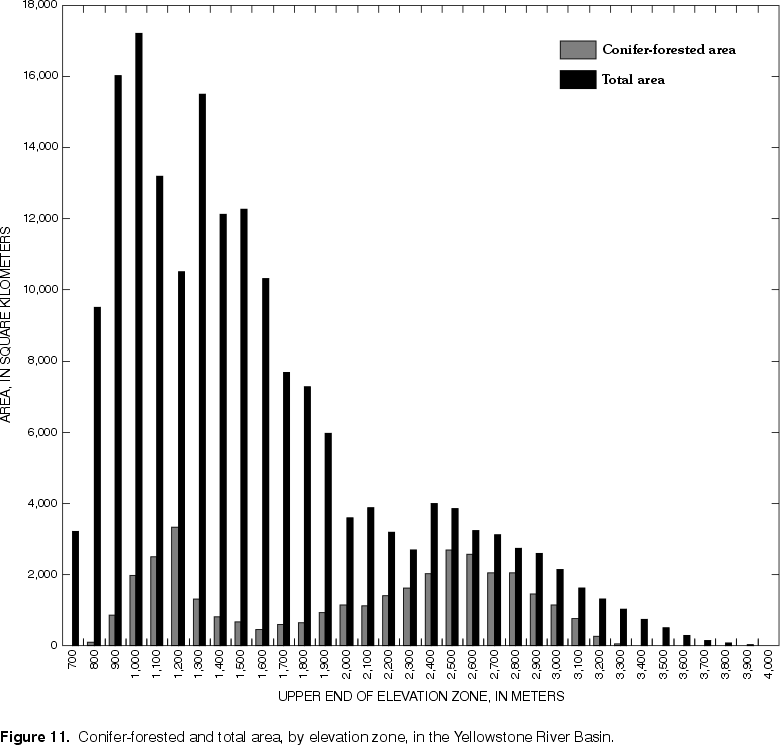 |
| Figure 11. Conifer-forested and total area, by elevation
zone, Yellowstone River Basin. (Click on image for a larger version, 41 kb) |
In addition to elevation, moisture gradients and disturbance history influence forest types. For example, in subalpine forests of Greater Yellowstone, subalpine fir is more common in moist locations, whereas lodgepole pine and Douglas fir dominate on drier slopes (Despain, 1990). Mountain vegetation zones extend to lower altitudes on wetter east- and north-facing slopes and steppe vegetation extends farther upward on south- and west-facing ones (Bailey, 1995; Marston and Anderson, 1991).
A major disturbance such as fire restarts the long successional process by which vegetation progresses through a series of plant communities toward the climax community (Despain, 1990). Under a natural fire regime, the wildfire return interval in most of the mountain forests in western parts of the YRB is between 200 and 400 years (Despain, 1990; Romme, 1980). Such intervals do not necessarily allow the successional process to culminate. In addition to vegetation disturbance, forest fires produce increased stream discharge, erosion, and sediment transport from severely burned watersheds (Tiedemann and others, 1979). Also, loss of riparian vegetation and woody debris jams increases the risk of serious washouts of organic matter, nutrients, and sediment stored in channels if high-flow events shortly follow the burning of much of a watershed (McIntyre and Minshall, 1996). The large fires of 1988 in the Greater Yellowstone Area burned more than 3,300 km2 of mostly forested land in the YRB (Minshall and Brock, 1991; Greater Yellowstone Post-Fire Resource Assessment Committee, 1988). The fire-related changes were expected to cause substantial impacts to extensively burned watersheds, but large watersheds (greater than 500 km2) were only partially burned and principal streams were little affected (Minshall and others, 1989; Minshall and Brock, 1991, p. 126).
Soil and rock types also affect forest communities through nutrient and moisture availability. For example, in the Greater Yellowstone Area, soils formed from andesite have good water-holding capacity and mineral nutrient content, whereas soils formed from rhyolite are coarse textured, with poor water-holding capacity and only one-third the nutrient level of andesitic soils (Despain, 1990). As a result, distinctly different forest communities are found on these contrasting soils: lodgepole pine is persistently dominant in areas underlain by rhyolite flows, whereas spruce-fir communities are usually dominant in old-growth forests on soils formed from andesite (Marston and Anderson, 1991).
In the lower-elevation, open woodlands of the Great Plains Province, ponderosa pine is the dominant species. Such forest lands cover about 6 percent of the study unit, and correspond to the peak in conifer-
forested area at about 1,200 m elevation (fig. 11). Grasses, with some sagebrush, typically compose the ground cover and understory in the open woodlands (Bailey, 1995).
Coniferous forests actually cover about 19 percent of the study unit, whereas they potentially would cover about 28 percent under natural conditions (Kuchler, 1964). The major forest types and percentage of YRB forested land they occupy are lodgepole pine (34 percent), ponderosa pine (22 percent), spruce-fir (19 percent), Douglas fir (14 percent), broadleaf hardwoods (6 percent), and pinyon-juniper (4 percent) forests (U.S. Department of Agriculture, Forest Service, 1970).
Steppe is the most widespread vegetation class in the YRB, covering about 44 percent of the study unit, as estimated by comparing data from Bailey (1995) and U.S. Geological Survey (1986). Kuchler's map (1964; fig. 10) shows three categories corresponding to steppe: foothills prairie, grama-needlegrass-wheatgrass grassland, and wheatgrass-needlegrass grassland. The short, typically bunched grasses of the steppes include predominantly grama, needlegrass, and wheatgrass (Bailey, 1995; Kuchler, 1964; Marston and Anderson, 1991). Numerous species of wildflowers bloom during spring and summer, with sunflower among the most prominent (Bailey, 1995). Sagebrush is an important component of the vegetation mosaic in many grassland areas. Prickly-pear cactus apparently was abundant in the early 1800s, as it is today (U.S. Department of the Interior, 1965). Invasive exotic species, such as Russian thistle, are abundant in some locales. Except in moist hollows and valley floors, steppe grasses are usually sparse, with bare soil exposed between the bunches (Bailey, 1995).
Sagebrush steppe or semidesert shrub is the dominant vegetation type in about 21 percent of the study unit (compare Bailey, 1995, and U.S. Geological Survey, 1986). Kuchler's map (1964; fig. 10) indicates that these shrub lands (wheatgrass-needlegrass shrub steppe, sagebrush steppe, and saltbush-greasewood shrubland) would potentially cover about 25 percent of the study unit. Typically, this vegetation type is a mixture of sagebrush with short grasses, but greasewood is prominent in extensive alkaline areas of the Bighorn Basin (Bailey, 1995). Sagebrush is typically found on well drained sites where most of the precipitation occurs as snowfall, whereas grasslands tend to be found where summer rainfall predominates (Knight, 1990). Other common shrubs include rabbitbrush, bitterbrush, and serviceberry (Marston and Anderson, 1991).
In the YRB, typically narrow riparian zones lie between the aquatic and terrestrial ecosystems and include several fluvial surfaces: channel islands and bars, channel banks, floodplains, and lower terraces on which the vegetation is dominated by deeply rooted plants capable of extracting water from the alluvial aquifer (Goodwin and others, 1997). The riparian vegetation community along a stream controls or influences several important ecologic characteristics, such as availability of habitat, canopy closure above the channel, water temperature, photosynthetic productivity, input of allochthonous resources, benthic invertebrate community composition, stream bank stability, density of coarse woody debris elements, and step size in step-pool channels (Hupp and Simon, 1986; Mihuc and others, 1996; Sweeney, 1992). Riparian vegetation may additionally benefit water quality by filtering sediments and utilizing nutrients (Correll and others, 1992; Knight, 1994). Riparian vegetation communities are highly productive and support an abundance of wildlife (Marston and Anderson, 1991), and domestic livestock also congregate there (Knight, 1994). Concentrations of large ungulates may trample banks and contribute to increased erosion and embeddedness. In contrast, beaver often produce a beneficial effect on water quality through dam building (Knight, 1994). Dams arrest streambed degradation, sedimentation behind the dams allows for greater stream clarity downstream, and enhanced bank storage of water results in higher late-summer streamflow that is beneficial to fish and wildlife.
Riparian vegetation is highly variable in the YRB and may include marsh, meadow, shrublands, and forest communities (Knight, 1994). At higher elevations, sedges, grasses, and short willows dominate the community typically found along headwater streams, but alders and tall willows become more conspicuous along lower mountain streams (Knight, 1994). In the foothills and intermontane basins, the riparian communities generally are dominated by cottonwoods, with aspens, lodgepole pine, and spruces also common (Knight, 1994; Marston and Anderson, 1991). Along many streams of the eastern plains, grasses, rushes, and sedges are dominant plants in herbaceous riparian communities; and black greasewood, common chokecherry, coyote willow, silver buffaloberry, silver sagebrush, and western snowberry are dominant species in shrub riparian communities (Jones and Walford, 1995). The riparian zone along many other lowland streams is typically a woodland community with plains cottonwood being most abundant, and ash, boxelder, willows, and the exotic Russian olive all being common locally (Jones and Walford, 1995; Knight, 1994). The abundance of old-growth cottonwoods appears to be declining in some areas due to flood suppression (Knight, 1994). Cottonwood regeneration is related to flooding and associated creation of bare sand and gravel bars that this pioneer species colonizes. Northern floodplain deciduous forests (fig. 10) occupy about 1 percent of the YRB, mostly as riparian communities along several of the principal streams (not only the Yellowstone River).
Other prominent vegetation in the study unit includes native hay, alfalfa, and seasonal coverage by crops such as small grains, beans, sugar beets, and corn. Agricultural lands are discussed in more detail in a separate section of this report.
Return to Environmental Setting of the Yellowstone River Basin, WRIR 98-4269