USGS Circular 1316
Synthesis of U.S. Geological Survey Science for the Chesapeake Bay Ecosystem and Implications for Environmental Management | ||||
Chapter 12: Factors Affecting Coastal Wetland Loss and Restoration | ||||
|
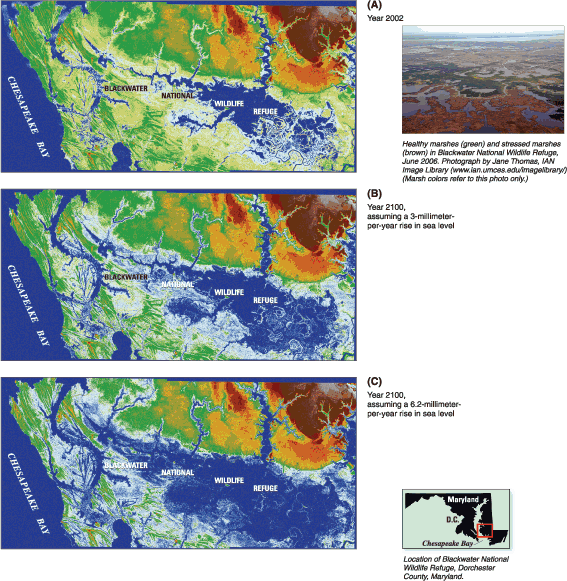
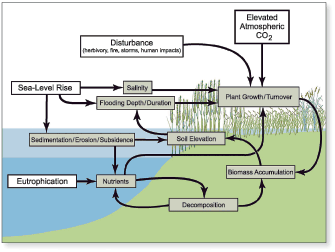
Tidal and nontidal wetlands in the Chesapeake Bay watershed provide vital hydrologic, water-quality, and ecological functions. Situated at the interface of land and water, these valuable habitats are vulnerable to alteration and loss by human activities including direct conversion to non-wetland habitat by dredge-and-fill activities from land development, and to the effects of excessive nutrients, altered hydrology and runoff, contaminants, prescribed fire management, and invasive species. Processes such as sea-level rise and climate change also impact wetlands. Although local, State, and Federal regulations provide for protection of wetland resources, the conversion and loss of wetland habitats continue in the Bay watershed. Given the critical values of wetlands, the Chesapeake 2000 Agreement has a goal to achieve a net gain in wetlands by restoring 25,000 acres of tidal and nontidal wetlands by 2010. The USGS has synthesized findings on three topics: (1) sea-level rise and wetland loss, (2) wetland restoration, and (3) factors affecting wetland diversity. Chesapeake Bay is a drowned-river-valley estuary where emergent tidal wetlands migrate landward (upslope) in response to sea-level rise through the accumulation of mineral sediments and plant matter. Wetlands convert to shallow, open-water habitat (such as ponds) through interior marsh breakup if they do not build vertically at a pace equal to sea-level rise, which is currently about 3 mm/yr (millimeters per year) in the Bay (Douglas, 2001). The majority of tidal marsh in Chesapeake Bay is in the lower part of Marylands Eastern Shore. Extensive areas of submerged upland marshes in the Blackwater River-Fishing Bay region of Dorchester County, Maryland have converted to open water over the past century, particularly those marshes at Blackwater National Wildlife Refuge (BNWR). The rate of sea-level rise is predicted to increase two- to four-fold during the next century (Church and others, 2001). To determine what impact this sea-level change would have on wetland resources and to improve land-use planning within the immediate vicinity of BNWR for the next century, USGS scientists developed a digital elevation model (DEM) of BNWR land surfaces from LIDAR (Light Detection And Ranging) data collected in March 2002 (fig. 12.1A) (Larsen and others, 2004). DEM simulations using current sea-level rise rates (approximately 3 mm/yr) reveal that high marsh will convert to low marsh and low marsh will continue to convert to open water for the next century, assuming 2002 surface elevations remain unchanged (fig. 12.1B). Marsh loss rates will be higher, and the area impacted larger, for predicted future rates of sea-level rise (about 6 mm/yr) (fig. 12.1C). Measurements of marsh vertical accretion, marsh-surface elevation change, and shallow soil subsidence made by USGS scientists over 5 consecutive years reveal that marsh-surface elevations are not static but are actually decreasing at most sampling stations at BNWR (G. Guntenspergen, U.S. Geological Survey, written commun., 2007). The declining marsh surface elevations at BNWR indicate that the DEM projections likely underestimate the extent of future marsh loss. The BNWR marsh system is characterized by low mineral sediment supply. Although major storms, such as Hurricane Isabel in 2003, deposit mineral sediments on the marsh every few decades, the increase in marsh elevation is often minimal. This soil organic matter accumulation comprised mostly of plant roots plays an important role in vertical soil development. Several factors affect the ability of the marshes at BNWR to build vertically through soil matter accumulation and therefore likely influence the rate of ongoing interior marsh breakup. These factors include grazing of vegetation by muskrat and nutria, altered flooding and salinity patterns, annual prescribed burning of vegetation, overabundance of nutrients, subsidence, and changes in the rate of sea-level rise (fig. 12.2). For example, intense grazing of marsh vegetation by nutria, an exotic species introduced to the United States from South America, severely reduced plant production at BNWR. Following the removal of more than 9,000 nutria from the region between 2002 and 2004, there has been strong recovery of marsh vegetation (M. Haramis, U.S. Geological Survey, written commun., 2007). These findings imply that the combination of sea-level rise and factors affecting sediment accumulation rates will govern the rate of wetland loss along the estuary. Thus, resource managers will have to fully understand the combination of factors affecting marsh loss at a particular site for successful wetland restoration. Sediments dredged from Chesapeake Bay navigation channels are being used to restore degraded wetland habitats within the Bay. During the past decade, several wetland restoration projects using dredged sediments have been undertaken, including Poplar Island, Anacostia River, and Barren Island. USGS investigations at Poplar Island brackish marshes in the central Bay (Erwin and others, 2003) and Kenilworth and Kingman tidal freshwater marshes in the Anacostia River, Washington, D.C. (Hammerschlag and others, 2006), revealed some of the important processes controlling and limiting habitat quality in these reconstructed wetlands.
At Kingman and Kenilworth marshes, studies showed there is a large, functionally diverse seed bank of wetland species in the Anacostia River available to colonize dredged sediment deposits (Neff and Baldwin, 2005). The combination of natural colonization and vegetative planting efforts facilitated rapid development of vegetated marsh habitats wherever restored soil elevations were suitable (Hammerschlag and others, 2006). However, marsh establishment at several sites was affected by grazing from an overabundance of resident Canada geese. USGS scientists also found geese herbivory to be an important factor in the decline of wild rice along the tidal Patuxent River (Haramis and Kearns, 2007). Removal of geese by hunting, and efforts to protect and re-establish rice by fencing and planting, led to successful restoration of this marsh type. The results imply that tidal wetland creation from dredged sediments is an effective method for restoring wetland habitats when the proper intertidal soil elevations are established and maintained and herbivory is managed. The results also imply the presence of an existing seedbank may enhance the success of wetland restoration. USGS studied the correlation between wetland restoration and changes in bird populations at Poplar Island, which eroded to less than 5 acres in 1996, and is undergoing wetland restoration that will include 550 or more acres of constructed tidal wetlands, creeks, ponds, and mudflats. USGS found that the restored habitat is attracting desired common terns, least terns, snowy egrets, cattle egrets, American black ducks, and osprey (Erwin and others, 2003). This site is the only nesting area for common terns in the Maryland part of the Bay and thus, is critical to species survival in Maryland. However, constructed upland habitats also attract undesirable bird species such as gulls and great horned owls and mammal predators (red foxes) that harass or prey upon the desired bird species. USGS studies showed the diversity of coastal and nontidal wetlands are affected by multiple factors. Grazing by exotic species such as resident Canada geese at Anacostia River marshes (see above) can prohibit plant development and change vegetation composition. Exotic colonizers, such as Phragmites, can out-compete native vegetation and cause a loss in diversity and in habitat value and function. USGS studies of native and invasive varieties of Phragmites reveal that the invasive variety can grow in saltwater concentrations at which the native varieties cannot survive. They also produce more shoots per gram of rhizome tissue and have a higher relative growth rate than the native varieties (Vasquez and others, 2005). These findings imply that the diversity of a tidal wetland will depend on controlling competition and predation from non-desired species, which also attempt to colonize restored and native wetlands. In forested wetlands, patterns of plant zonation and diversity are strongly influenced by physical conditions, such as flooding patterns related to variations in river flows and local geomorphology (such as hydrogeomorphology). Nontidal riparian and flood-plain wetland communities are typically highly diverse areas in the landscape, but the reasons for this are poorly understood. Although plant diversity and composition can be attributed in part to hydrologic conditions (such as seasonal flooding patterns), recent USGS investigations reveal that hydrologic conditions alone do not describe forested wetland plant patterns (Alexander-Augustine and Hupp, 2002). Plant diversity is strongly impacted by hydroperiod (the period during which wetlands are flooded), micro-scale changes in relief, and upstream-downstream position within the stream corridor. More importantly, however, the influence of hydrogeomorphology on species richness varies with spatial scale. Species richness was described by hydrogeomorphic variables (downstream position, river discharge, stream power, and topographic relief) at the plot scale (400 square miles). Tree diversity was best explained at the site scale (1 hectare), and hydrogeomorphic variables were best explained at the watershed scale (Alexander-Augustine and Hupp, 2002). Thus, a combination of spatial, hydrologic, and geomorphic conditions explains plant diversity patterns in forested wetlands. These findings imply that the diversity of a tidal and forested wetland will depend on controlling competition and predation from non-desired species, which also attempt to colonize a restored wetland. Resource managers need to understand these conditions when developing management plans for riverine wetlands. ReferencesAlexander-Augustine, L.E., and Hupp, C.R., 2002, Process and pattern: Hydrogeomorphology and biodiversity in forested wetlands on the Chesapeake Bay Coastal Plain, in Proceedings of the 87th Annual Meeting of the Ecological Society of America and the 14th Annual International Conference of the Society for Ecological Restoration, August 49, 2002, Tucson, Arizona. Church, J.A., Gregory, J.M., Huybrechts, P., Kuhn, M., Lambeck, K., Nhuan, M.T., Qin, D., and Woodworth, P.L., 2001, Changes in sea level, in Climate Change 2001: The scientific basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change, Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der Linden, P.J., Dai, X., Maskell, K., and Johnson, C.A., eds.: Cambridge, United Kingdom and New York, New York, USA, Cambridge University Press, p. 639693. Douglas, B.C., 2001, Sea level change in the era of the recording tide gauge, p. 3764, in Douglas, B.C., Kearney, M.S., and Leatherman, S.P., eds., Sea level rise: History and consequences: San Diego, California, Academic Press, 272 p. Erwin, R.M., Brinker, D., and Fruh, B., 2003, Poplar Island environmental restoration project: Build it and they (wildlife) will come, but are all welcome?, in Proceedings of the 13th Biennial Coastal Zone Conference, Baltimore, Maryland, July 1317, 2003, NOAA/CSC/20322-CD. CD-ROM. Hammerschlag, R.S., Baldwin, A.H., Krafft, C.C., Paul, M.M., Brittingham, K.D., Rusello, K., and Hatfield, J.S., 2006, Final ReportFive years of monitoring reconstructed freshwater tidal wetlands in the Urban Anacostia River, accessed June 6, 2007, at http://www.pwrc.usgs.gov/resshow/hammerschlag/anacostia.cfm. Haramis, G.M., and Kearns, G.D., 2007, Herbivory by resident Canada geese: The loss and restoration of wild rice along the tidal Patuxent River, Maryland: Journal of Wildlife Management, v. 71, p. 788794. Larsen, C., Clark, I., Guntenspergen, G.R., Cahoon, D.R., Caruso, V., Hupp, C., and Yanosky, T., 2004, The Blackwater NWR inundation model. Rising sea level on a low-lying coast: Land use planning for wetlands: U.S. Geological Survey Open File Report 041302, available online only at https://pubs.usgs.gov/of/2004/1302/ Neff, K., and Baldwin, A., 2005, Seed dispersal into wetlands: Techniques and results for a restored tidal freshwater marsh: Wetlands, v. 25, no. 2, p. 392404. Vasquez, E.A., Glenn, E.P., Brown, J.J., Guntenspergen, G.R., and Nelson, S.G., 2005, Salt tolerance underlies the cryptic invasion of North American salt marshes by an introduced haplotype of the common reed Phragmites australis (Poaceae): Marine Ecology Progress Series, v. 298, p. 18. | |||

![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: http://pubsdata.usgs.gov/pubs/circ/circ1316/html/circ1316chap12.html
Page Contact Information: GS Pubs Web Contact
Page Last Modified: Tuesday, 10-Jul-2018 11:22:01 EDT