

Rearing method has limited effect on post-release movement of reintroduced age-0 Lake Sturgeon
Links
- More information: Publisher Index Page (via DOI)
- Document: XML
- Open Access Version: USGS Accepted Manuscript
- Download citation as: RIS | Dublin Core
INTRODUCTION
Many populations of Lake Sturgeon (Acipenser fulvescens) no longer exist throughout their historical native range (Ferguson and Duckworth 1997, Pollock et al. 2015). Within the Laurentian Great Lakes, Lake Sturgeon declines were caused by overharvest, pollution, and habitat loss (Auer 1996, Hay-Chmielewski and Whelan 1997, Peterson et al. 2007, Bruch et al. 2016). Lake Sturgeon are a long-lived species, late to mature (15–18 yr), and are intermittent spawners where males might spawn every 1-3 years and females every 4-9 years (Peterson et al. 2007, Smith and Barker 2005). These life history traits coupled with ongoing water and habitat quality issues continue to hinder Lake Sturgeon recovery (Harkness and Dymond 1961, Auer 1996, Shane et al. 2021). As a result, Lake Sturgeon are currently listed as a species of concern, threatened, or endangered in all states and provinces throughout the Great Lakes region (Bruch et al. 2016).
Within Ohio’s portion of Lake Erie, Lake Sturgeon remain extirpated from tributaries that historically had naturally reproducing populations (Goodyear 1982, Hay-Chmielewski and Whelan 1997, Boase 2008, Haxton et al. 2014). A large self-sustaining Lake Sturgeon population exists in the St. Clair-Detroit River System between Lake Huron and Lake Erie (Chiotti et al. 2023), which is about 40 km from the Maumee River. Current sampling efforts in the region have found no evidence that straying from the St. Clair-Detroit River System has led to the recolonization of the Maumee River (Kessel et al. 2018), despite environmental management actions that have improved the water quality (Richards et al. 2008) and habitat leading to the recovery of many fishes in the Maumee River (e.g., Catostomus spp., Moxostoma spp., Lepomis spp., Leuciscidae spp.; Mapes et al. 2015, Shane et al. 2021). Collier (2018) found there was sufficient suitable habitat in the Maumee River that would support Lake Sturgeon spawning and the recruitment of younger age classes. The presence of sufficient habitat but absence of Lake Sturgeon prompted a reintroduction effort in 2018 between state and federal agencies along with academic and private partners to reestablish a naturally reproducing Lake Sturgeon population in the Maumee River.
The long-term goal of the Maumee River reintroduction plan is to establish a self-sustaining population of Lake Sturgeon with a target of 1,500 naturally spawning adults (Collier et al. 2022). To achieve this goal, the plan is to stock 3,000 age-0 sturgeon each year into the Maumee River for at least 20 years (e.g., Schram et al. 1999). An experimental approach was adopted in the plan where sturgeon were raised either in a traditional hatchery at the Genoa National Fish Hatchery (Wisconsin, USA) or at the Toledo Zoo (Ohio, USA) in a streamside facility that used Maumee River water. Streamside techniques are used to expose early life stages to the water of rivers targeted for reintroduction in hopes of facilitating imprinting, increasing survival, and developing the movement and habitat selection patterns of natural cohorts (Holtgren and Auer 2004, Holtgren et al. 2007, Crossman et al. 2011, Mann et al. 2011). However, streamside rearing approaches are not always feasible for each river where managers might want to reintroduce Lake Sturgeon. A traditional hatchery approach allows for large numbers of fish to be raised and ultimately stocked without having to set up hatcheries in multiple locations (Trushenski et al. 2010).
Lake Sturgeon can remain in natal rivers as residents (Auer 1999, DeHann et al. 2006), move between rivers and connected lakes (Hay-Chmielewski and Whelan 1997, Boase et al. 2011, 2014), or move great distances away from stocking sites (Schram et al. 1999, Schram 2007, Welsh et al. 2019). We currently do not know the movement patterns for age-0 Lake Sturgeon reintroduced to the Maumee River, or if rearing strategy influences movement or habitat areas used by the sturgeon. Our objectives were to 1) determine whether reintroduced age-0 Lake Sturgeon stayed in the Maumee River or moved out to the western basin of Lake Erie and what habitat area was used in the river or lake, 2) determine whether rearing strategy affected sturgeon movement and area use, and 3) identify options that might facilitate the evaluation and success of Lake Sturgeon establishing a naturally reproducing population in the Maumee River. We tagged stocked age-0 Lake Sturgeon with acoustic transmitters in three years: 2018, 2019, and 2021. Our scope of inference was limited to approximately one-year post-release based on the estimated battery life of the tags. We predicted that streamside-reared Lake Sturgeon would stay in the Maumee River longer than sturgeon reared at Genoa National Fish Hatchery because of familiarity with river water chemistry (Crossman et al. 2011). We also predicted streamside sturgeon would exhibit shorter movements away from the Maumee River than traditionally reared sturgeon, and that movements by streamside-reared sturgeon might reflect the behavior of natural cohorts if they were to be reestablished in the river (e.g., Mann et al. 2011). We assumed if the sturgeon spent more time in the Maumee River, this could facilitate imprinting and system learning to increase the probability of returning to the river to spawn as adults (Holtgren et al. 2007, Crossman et al. 2011, Crossman et al. 2013).
METHODS
Study area
Our study area was focused on an approximately 22-km segment of the Maumee River, stretching from the Audubon Islands to the river’s confluence with Maumee Bay and extending into the western basin of Lake Erie and ending east of the islands region (i.e., Bass islands, Kelleys and Pelee islands; Fig. 1). Located downstream of the Grand Rapids and Providence dams, this section of the Maumee River supports a range of aquatic environments, including shallow bedrock, gravel substrates, periodic rapids, and a shipping channel (Shane et al. 2021, Collier et al. 2022). The adjacent western basin of Lake Erie is defined by its shallow depth (average depth ~7 m) and presence of numerous shoals and reef structures. We used acoustic receivers managed by the Great Lakes Acoustic Telemetry Observation System (GLATOS; https://glatos.org) to track post release movements and habitat area use of juvenile Lake Sturgeon within the Maumee River and western basin of Lake Erie (Fig. 1; Krueger et al. 2018).
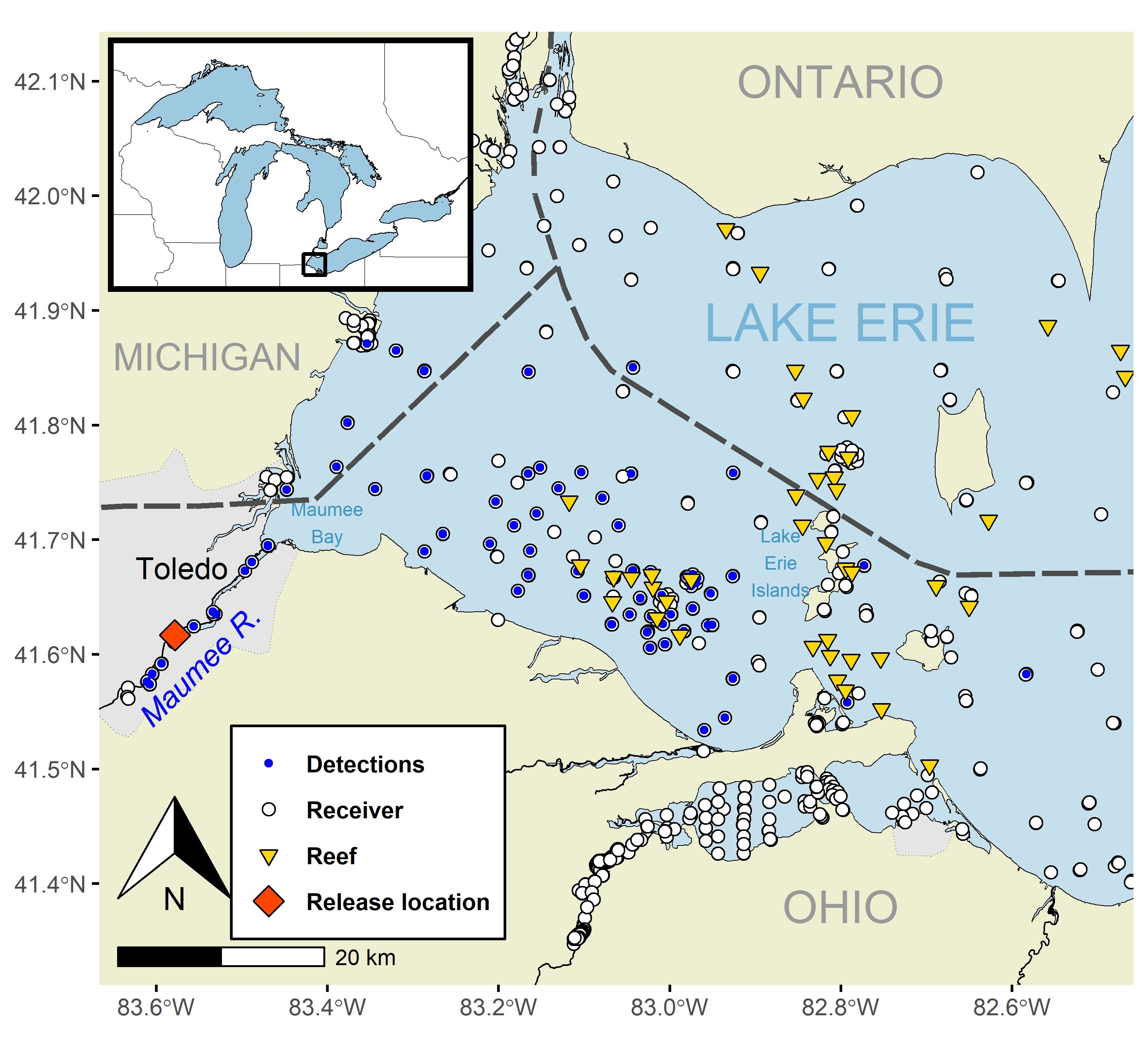
Study area with acoustic receiver locations (dots), reef locations in the western basin of Lake Erie (triangles), and the stocking site on the Maumee River (red diamond). Receivers where Lake Sturgeon were detected are filled with a blue dot. The black dashed lines represent the jurisdictional boundaries in Lake Erie.
Lake Sturgeon Rearing and Tagging
Detailed methods regarding age-0 Lake Sturgeon rearing and acoustic tagging are described in McKenna et al. (2025). To summarize, we selected forty individuals in 2018, 2019, and 2021 to receive V7 acoustic transmitters (69 kHz, 19.5mm long, 1.5g in air, 0.7g in water; Innovasea, Halifax, NS; see Table 1). The transmitters were programmed to transmit for 60 days and pause for 30 days to conserve battery life using low power (137 dB), with a random delay of 190–290 seconds (Table 1). This programming maximized how long we could infer movement and areas used (about one year) given the small tag size necessary for use with age-0 sturgeon. A longer delay would have likely reduced our detection probability substantially while the tags were active. However, the pause introduced some limitations. Specifically, we could not know if sturgeon moved between the river or lake, or more broadly, what areas were used during the pause.
Table 1.
Sample size, size metrics (mean ± 1 SD), data collection period and activation dates for the v7 acoustic tags, and the estimated battery life for the first three years of a Lake Sturgeon (Acipenser fulvescens) reintroduction effort in the Maumee River (Ohio, USA).Surgeries to implant acoustic transmitters were completed by veterinary staff at the Toledo Zoo’s stocking trailer. Lake Sturgeon were held for one to two weeks to ensure adequate recovery time before being released at a public boat ramp adjacent to the Toledo Zoo at about four months of age (Toledo, OH; 41.6144, -83.5811; Fig. 1). All but three tagged fish were detected on at least one receiver in the Maumee River on the day of release; the remaining three were detected the following day.
Telemetry Data Processing
Within Lake Erie, large-scale grids of receivers are maintained year-round (Kraus et al. 2018), while smaller arrays are deployed seasonally to address project-specific objectives (e.g., Bade et al. 2019). Functionally, this means that arrays of acoustic receivers can change with the addition of new projects or as locations of existing receivers shift throughout the project period as seen in Fig. 1 of McKenna et al. (2025). Due to receiver arrays changing between the years, we analyzed our data individually by year to account for changes in receiver effort. Detection data are uploaded to the GLATOS interface where information related to specific telemetry projects can be exported for analysis. Data collection periods for each release year are reported in Table 1. The last data export from GLATOS for data used in this study was 3 April 2023.
We processed detection data in RStudio version 4.2.0 (R Core Team 2022) using the ‘glatos’ package (Holbrook et al. 2017). We removed potential false detections using the false_detection() function following Pincock (2012). This resulted in the removal of 1,265 detections (1.73%), leaving a total of 71,676 detections for analysis of Lake Sturgeon released in 2018, 2019, and 2021.
Movement and Habitat Area
We assessed whether reintroduced sturgeon stayed in the Maumee River or moved out to Lake Erie by plotting the number of unique Lake Sturgeon detected each day within a release year. We also compared the number of days detected in the river to days detected within the lake for each rearing strategy within a release year. We condensed the raw detection data using the detection_events() function using one-day as the time separation value with the ‘glatos’ package. This function simplified the detection data, filtering 71,676 detections to 490 unique detection events. We then used the residence_index() function to calculate the residency index for each fish within each GLATOS array using the Kessel method (Kessel et al. 2016), which created a list of unique days (i.e., river and lake days) where detections occurred on the array using one day as our time interval within the life of the acoustic tags.
For each release year, we used an acoustic receiver deployed near the mouth of the Maumee River to infer entry and exit from the river (MAU-015: 41.69467, -83.46895). We determined that this receiver provided a reasonable indicator of fish exiting the river: of 74 fish detected in the lake from all release years, 56 were detected on this receiver before exiting the river, while 18 were not detected. A total of 46 fish were never detected in the lake; however, 35 were detected on the river mouth receiver, while the remaining 11 fish were not detected on this receiver. A majority of the fish that were not detected in the lake were from the 2018 release year (n = 28). Therefore, some downstream movements towards Lake Erie were undetected possibly due to the programmed pause in tag transmission, low detection probability, changes in receiver arrays or locations, low power of the V7 transmitters, or a combination of these factors.
To calculate the total number of days spent in the river or lake, we calculated the number of days between the first and last detection in the river and the last detection in the river to the last day detected in the lake, respectively. This calculation of days includes time during the pause of 30 days. For example, if a fish was detected in the river before and after the pause, those days were counted towards the ‘river days.’ If the detection after the pause occurred in the lake, the time from the last river detection to the first lake detection counted toward ‘lake days.’ While we cannot know if tagged fish moved between the river and lake during the 30-d pauses, Morton et al. (2025) suggested 18% of stocked age-0 Lake Sturgeon returned to the Genesee River (NY, USA) once they moved into Lake Ontario. Although 18% of individuals is not trivial, the results from Morton et al. (2025) indicated that the majority of age-0 Lake Sturgeon that leave a stocked river resided in and used lake habitats. We found no evidence of sturgeon moving between the lake and river when the tags were active but acknowledge this could have occurred for an unknown, but presumably small, number of sturgeon during the pause cycles.
We analyzed the number of days spent in the river versus the lake by rearing strategy for each release year using estimation statistics, specifically the ‘DABESTR’ package in R (Ho et al. 2019). This process has advantages over null-hypothesis significance testing in that the evaluation of statistical differences is not dichotomous (i.e., significant p-value or not); a confidence interval provides a specific measure of precision, whereas standard p-values often conflate an effect with a specific mean or sample size; error estimation with bootstrapping is robust; and effect sizes encourage quantitative reasoning over qualitative reasoning (e.g., see Ho et al. 2019). We illustrated the mean combined with bootstrapped 95% confidence intervals to evaluate differences within and among years and rearing strategies (Cumming 2012).
We calculated post-release habitat area (km2) used by Lake Sturgeon within each release year by rearing strategy using minimum convex polygons (MCPs) in ArcGIS Pro. We created MCPs from the short-term centers of activity (COAs) using the minimum bounding geometry tool in ArcGIS Pro. We calculated the COAs using the COA() function from the ‘V-Track’ package in R (Campbell et al. 2012), setting the time step equal to 60-minute time separations to minimize detections with the smaller nominal delay of the transmitters. We created COAs by estimating the mean location of receivers, which were weighted by the number of detections recorded at each receiver (Simpfendorfer et al. 2002). After importing the COAs into ArcGIS Pro, we clipped the MCPs to adjust the area of polygons to the boundary of the Maumee River and the western basin of Lake Erie using the clip geoprocessing tool, which adjusted the MCP measurements that were created over land instead of water. To transform the shape area measurement calculated from the MCPs, we used the calculate geometry attributes tool to calculate the MCP area in km2. To determine differences between rearing strategy and release years we examined 95% confidence intervals derived with the ‘DABESTR’ R-package, plotting the means and 95% confidence intervals. Differences were determined to occur if the 95% confidence intervals did not overlap. We additionally calculated differences in MCPs by rearing strategy to understand the extent of spatial overlap across all release years using the functions st_difference, st_intersection, and st_area from the ‘sf’ package (Pebesma 2018).
We acknowledge two important limitations of MCPs related to our study. First, MCPs are sensitive to outliers (Burgman and Fox 2003), which could make habitat area appear larger than it might actually be. Second, the MCPs could not distinguish between high-quality and low-quality habitats within the area used in our study. Alternative methods such as kernel utilization distributions (KUDs) exist to determine home range, habitat use, and habitat suitability (e.g., Vokoun 2003, Keating and Cherry 2009). However, we considered tradeoffs associated with using MCPs versus procedures like KUDs. Specifically, using KUDs would have required us to use receivers that were approximately in the same locations for the same duration during the three study years. Doing so would have substantially reduced the number of detections and important ecological information related to our study objectives because of the addition of receivers in 2019 (see McKenna et al. 2025). We wanted to maximize the number of detections available for our analyses, so we chose MCPs because of changes in receiver locations and density between release years, the small number of fish tagged each year (i.e. 40), and the programming pause to extend battery life.
We used distance traveled to understand whether the streamside-reared fish exhibited greater fidelity for the Maumee River than those raised at the Genoa National Fish Hatchery. We assessed the minimum distance traveled in kilometers using interpolated distance, which was the shortest estimated distance through water by detection location. To estimate interpolated paths, we used the interpolate_paths() function within the ‘glatos’ package in R. We calculated daily fish movements by summing distances along interpolated paths between detections within a 12-hr period. If a fish was only detected once within a time period on a single receiver, its movement was recorded as 0 km. For any gaps spanning multiple days, movement was estimated by dividing the distance between the last and next detection location and the timesteps between them similar to Harris et al. (2021). We then connected the minimum distance points to make paths using the st_cast() function and then calculated the minimum distances using the st_length() function from the ‘sf’ package. To visualize distance and movement patterns, we plotted the interpolated paths for each Lake Sturgeon in ArcGIS Pro. We assessed differences between rearing strategies and years through examination of 95% confidence intervals using the ‘DABESTR’ R-package. We determined statistical differences were supported by non-overlapping 95% confidence intervals.
RESULTS
All 120 tagged age-0 Lake Sturgeon were initially detected near the release location. Of the 120 sturgeon, 94 (78%) were detected in the Maumee River or Lake Erie one week after stocking. We did not detect 26 (22%) sturgeon one week after stocking with 10 (38%) being from 2018, 8 (31%) being from 2019, and 8 (31%) being from 2021. Although the fate of the remaining 26 fish beyond one week is uncertain, 25 were detected on multiple receivers within the Maumee River and were therefore included in our interpolated distance moved and MCP analyses. Only one fish from 2021 was excluded from these analyses due to being detected on a single receiver for a single day.
We found that the number of Lake Sturgeon in the Maumee River relative to Lake Erie decreased over time (Fig. 2A-C). The 2018 release year had the highest number of sturgeon that remained in the Maumee River. Of the 94 sturgeon detected beyond one week, we found 74 (79%) moved out of the river into the western basin of Lake Erie and 20 (21%) remained in the Maumee River after release. Of the 74 that moved out of the river, 30 (41%) were reared in the streamside facility and 44 (59%) were reared in a traditional hatchery. For the 20 sturgeon that stayed in the river, 13 (65%) were reared in the streamside facility and 7 (35%) were reared in a traditional hatchery. However, 18 (90%) of the 20 sturgeon detected in the river were from the 2018 release year, during which receiver coverage in the nearshore areas of Lake Erie was limited.
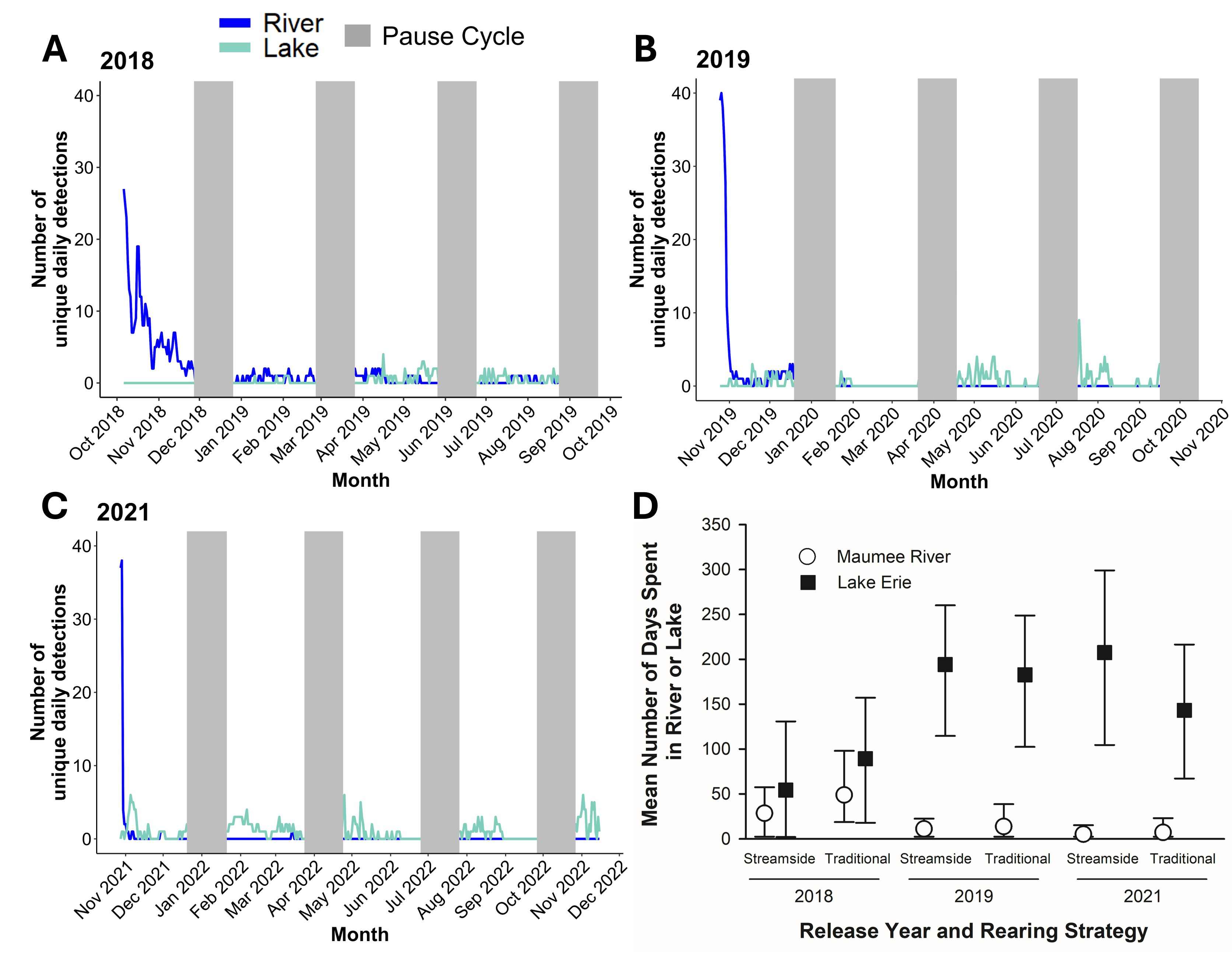
Number of unique daily detections of age-0 Lake Sturgeon for each release year that stayed in the Maumee River or left for the western basin of Lake Erie (A-C) and mean number of days (± 95% confidence intervals) Lake Sturgeon spent in the Maumee River (dots) versus Lake Erie (filled squares) for each release year and rearing strategy (D).
We found that the Lake Sturgeon spent more time on average in Lake Erie (54–207 d) than the Maumee River (3–47 d) after release (Fig. 2D). We found no effect of rearing strategy on the time spent within the river or lake among the three release years. Worth noting, the traditionally reared group spent on average 20 (74%) more days in the river compared to the streamside reared group in 2018, but there was overlap in the 95% confidence intervals indicating no support for this difference. In 2019 and 2021, sturgeon spent three to five days on average in the river regardless of rearing method.
Using MCPs, we did not find any differences in total habitat area used by Lake Sturgeon among the three release years (Fig. 3). Individual sturgeon from both groups used on average 19–81 km2 of habitat area, mostly in the western basin of Lake Erie. We found that sturgeon from both rearing strategies overlapped in 68% of the total habitat area used. Twenty-two percent of the habitat area used was unique to streamside reared sturgeon and 11% of the area used was unique to the traditionally reared sturgeon. However, two to three individuals represented the non-overlapping areas of use. Most of the habitat area used was in U.S. waters in the southern half of Lake Erie from the mouth of the Maumee River towards the islands area in the western basin.
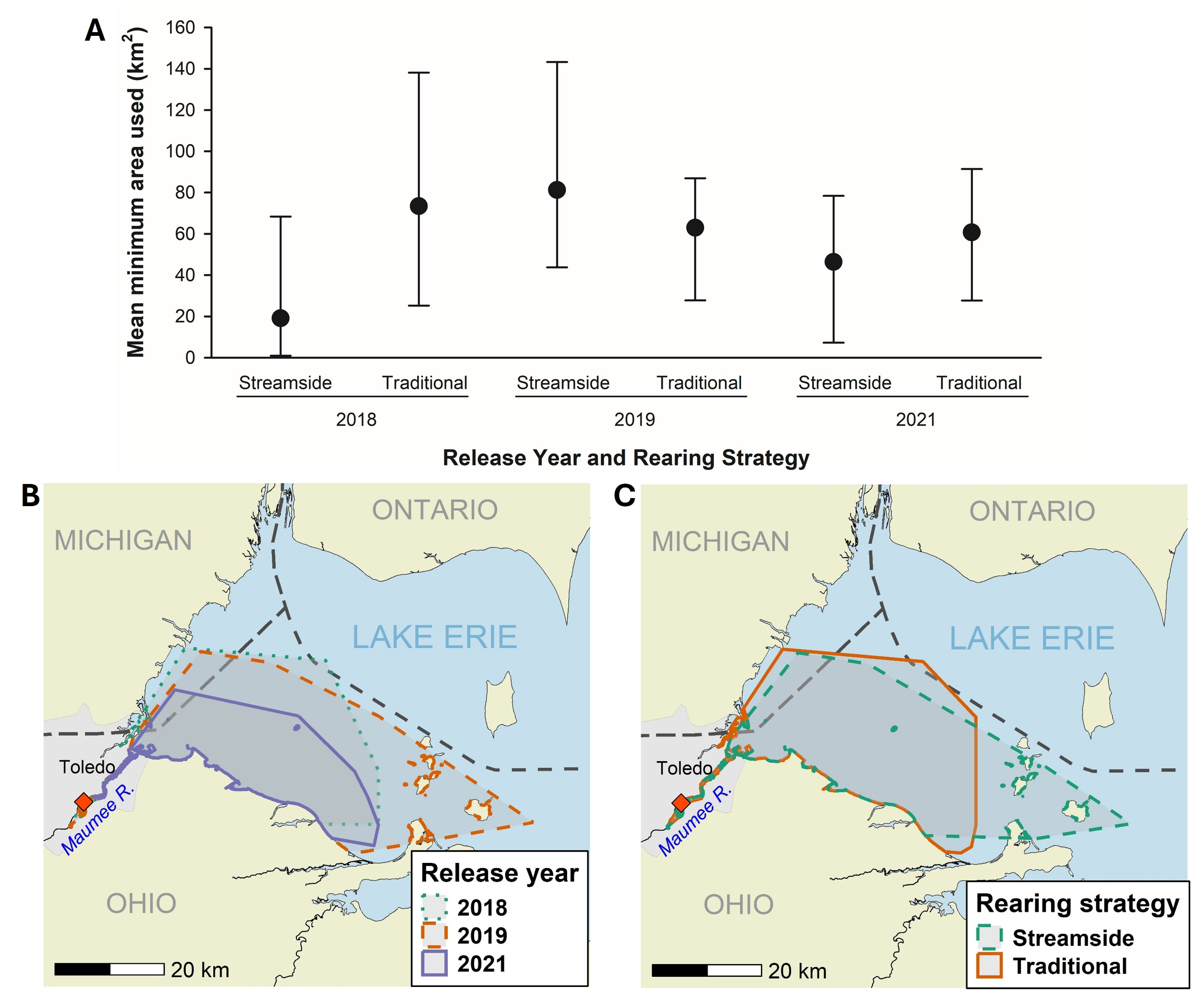
(A) Estimated mean minimum area used (km2; ± 95% confidence intervals) by reintroduced age-0 Lake Sturgeon for each release year and rearing strategy, (B) habitat area used for each release year combining rearing strategy, and (C) habitat area used for each rearing strategy combining all release years. Release location is indicated by a red diamond and the black dashed lines represent the jurisdictional boundaries in Lake Erie.
We estimated Lake Sturgeon moved, on average, 26–51 km after leaving the release site using interpolated distances. We did not find any differences in distances traveled between rearing strategy within any release year (Fig. 4). Sturgeon raised streamside in 2018 traveled shorter distances from the release site compared to other years, but distance traveled was comparable to the traditionally raised sturgeon and other release years because of overlapping confidence intervals. We also note that there were fewer receivers in 2018 compared to other release years.
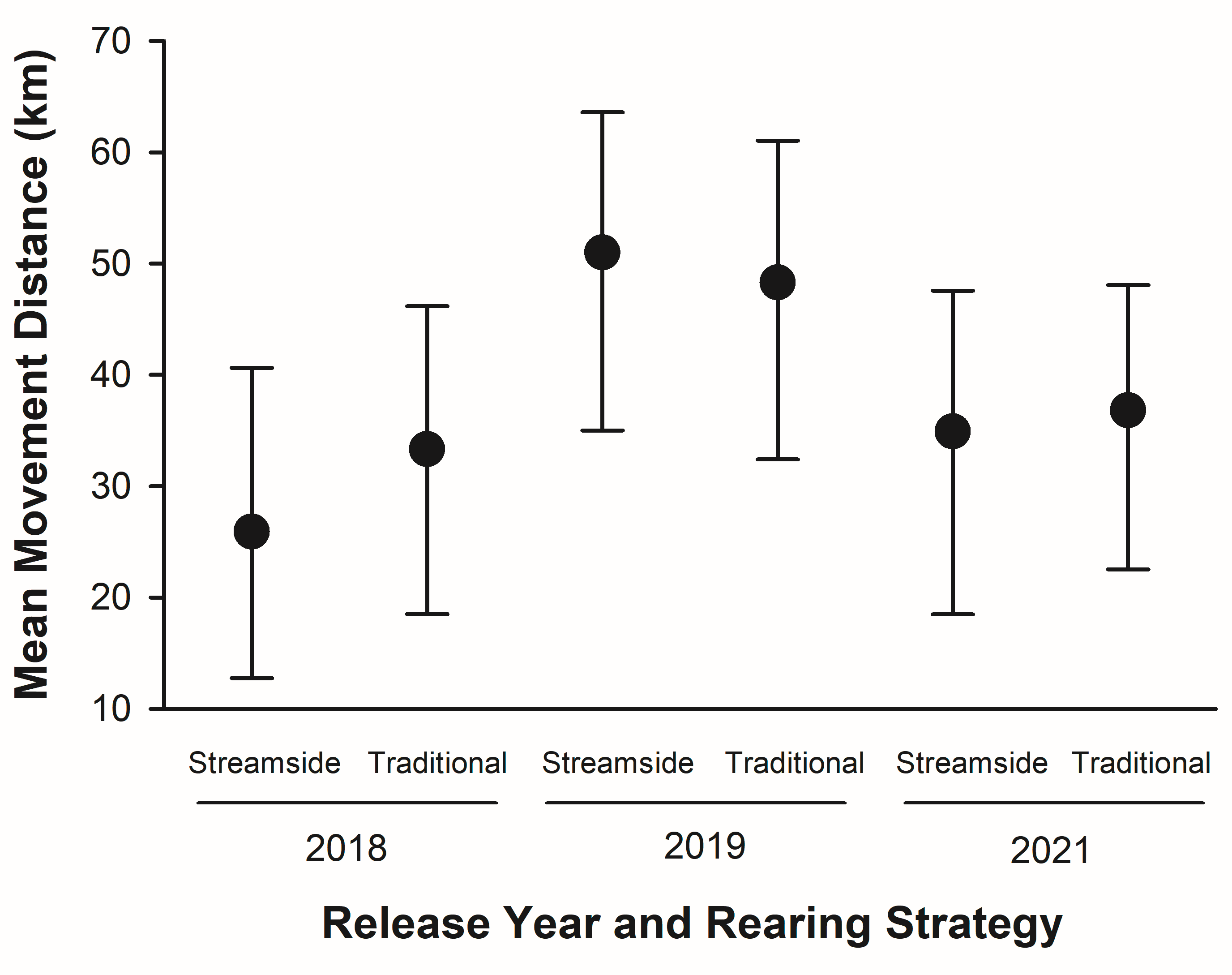
Estimated mean interpolated movement distance (km; ± 95% confidence intervals) by age-0 Lake Sturgeon reintroduced to the Maumee River for release years and rearing strategy.
Based on the interpolated paths for the 74 Lake Sturgeon that left the river for Lake Erie, movement paths in the lake were along the south shore of the western basin towards the islands area (Fig. 5). In 2018, 12 Lake Sturgeon (streamside “S” = 3, traditional “T” = 9) moved beyond the river with 58% (S = 2, T = 5) moving along the south shore of the western basin, 25% (S = 1, T = 2) moving toward the center of the western basin, and 17% (T = 2) moving along the shoreline of Michigan in a northeast direction. In 2019, 31 Lake Sturgeon moved beyond the Maumee River with 61% (S = 12, T = 7) moving along the south shore of the western basin of Lake Erie, 29% (S = 2, T = 7) moved towards the middle of the western basin, and 10% (S = 2, T = 1) moved northeast along the western shoreline. In 2021, 31 Lake Sturgeon moved beyond the Maumee River with 61% (S = 8, T = 11) having moved along the southern shore of the western basin while 29% (S = 2, T = 7) moved towards the middle of the basin and 10% (T = 3) traveled northeast along the Michigan shoreline.
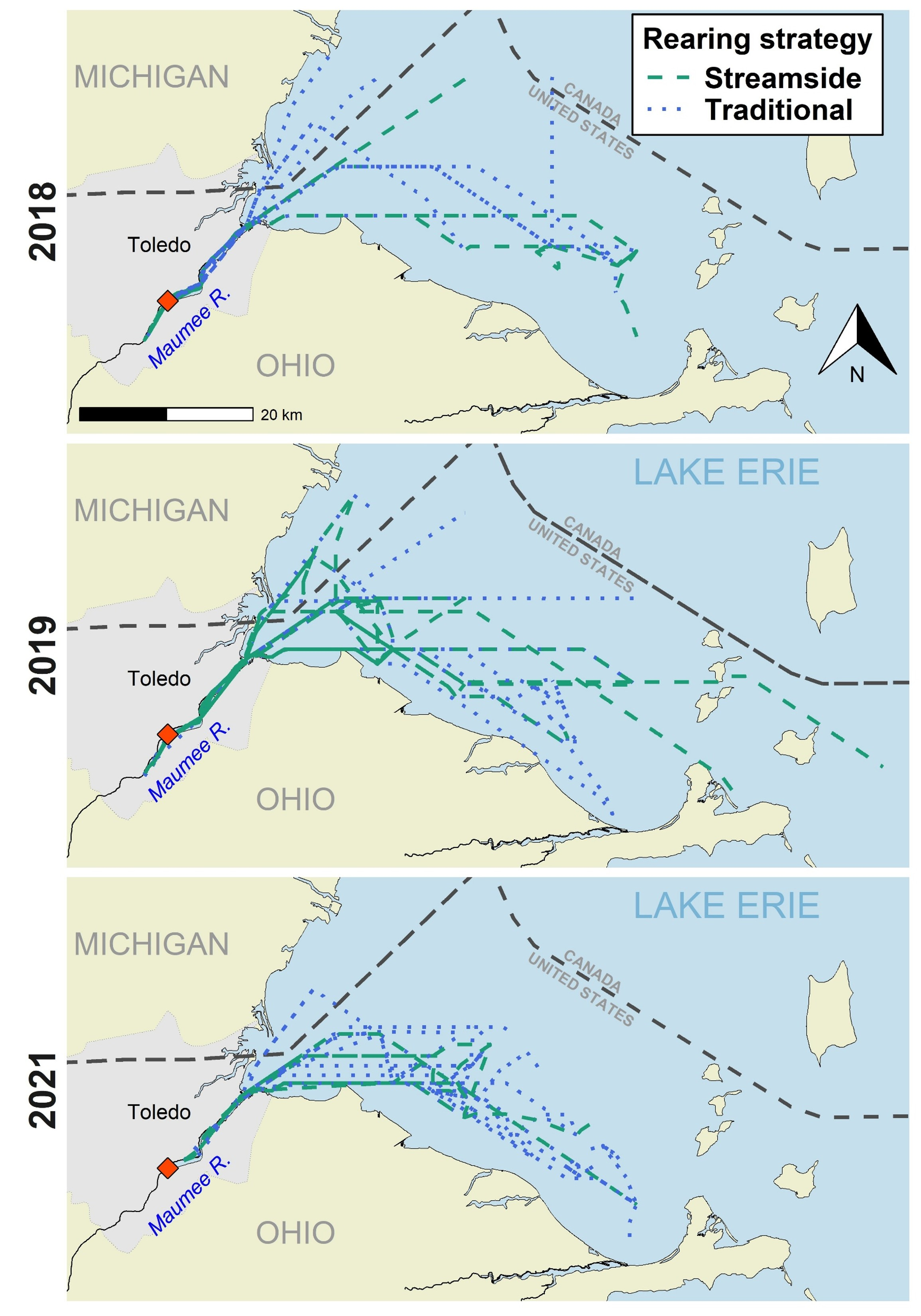
Interpolated paths of age-0 Lake Sturgeon in the Maumee River and the western basin of Lake Erie separated by a streamside raised group and traditional hatchery raised group. Release location is indicated by a red diamond and the black dashed lines represent the jurisdictional boundaries in Lake Erie. Samples sizes for each year are as follows: 2018: S = 20, T = 20; 2019: S = 20, T = 20; 2021: S = 11, T = 28.
DISCUSSION
Age-0 Lake Sturgeon exhibited similar movement behavior during the first three years of the reintroduction program regardless of rearing strategy. Most sturgeon moved downstream from the release site to Lake Erie, and a minority stayed in the Maumee River and were almost exclusively from the 2018 release year. Our evidence suggests that any imprinting that might have occurred during streamside rearing did not have a strong effect on river residency or movement behavior (also see Mann et al. 2011). However, reintroduced sturgeon might not reflect the imprinting process that occurs during early life until they reach maturity (e.g., Horrall 1981, Bett and Hinch 2016). There exists a need to investigate how rearing strategy influences the behavior of mature Lake Sturgeon that were part of the Maumee River reintroduction program—specifically, how rearing strategy influences spawning migrations, staging behavior, and spawning location selection. Such investigations could determine if successful imprinting occurred and facilitated natural spawning in the river and provide useful guidance for similar reintroduction efforts elsewhere.
The downstream movement to Lake Erie does not indicate streamside or traditionally reared fish will not return to spawn in the Maumee River. Downstream movement from rivers to lakes is a common behavior of reintroduced age-0 Lake Sturgeon. Half of the age-0 Lake Sturgeon stocked in the Genesee River (New York, USA) stayed in the river, while the other half left the river and moved into Lake Ontario (Morton et al. 2025). Further, some Lake Sturgeon reintroduced into the St. Louis River, a Lake Superior tributary, exhibited movement away from the stocking location into the lake (Schram et al. 1999, Schram 2007, Welsh et al. 2019). Movement distances by stocked individuals were up to 145 km for juvenile sturgeon and 300 km away as they became adults (Schram et al. 1999, Schram 2007). Despite some movement out of the St. Louis River and for long distances away from the river, many of the spawners in St. Louis River were of stocked origin (e.g., Welsh et al. 2019). To identify if spawners return to the Maumee River, it will be critically important to monitor the reintroduced Lake Sturgeon through maturity.
It is unclear whether movement out of the Maumee River and a greater residency in Lake Erie affected the imprinting phase of reintroduced Lake Sturgeon. The free-embryo and larval stages are critical for imprinting in fishes including Lake Sturgeon, but later life stages might also exhibit periods of imprinting (Dittman et al. 2015, Cathcart 2021, Kimmel et al. 2023). Although all sturgeon were stocked at approximately four months of age in the Maumee River, streamside-reared sturgeon were introduced to Maumee River water immediately upon fertilization, ensuring their exposure to the river environment within the critical window for imprinting as described by Kimmel et al. (2023). Straying rates are currently unknown for the Maumee River reintroduction program and will not be known until stocked fish begin spawning as adults. If evidence emerges that straying is an issue and the need arises to increase the likelihood of imprinting and subsequently promote adults returning to the Maumee River to spawn, strategies are available. Kimmel et al. (2023) suggest that hatchery-reared fish can respond behaviorally to chemical cues 50 days after a 7-day exposure history to target system odors. Stocking numbers could be increased at a streamside facility to offset the potential straying of traditionally raised sturgeon. Additionally, Maumee River water could be transported to a traditional hatchery and used in a recirculating culture system to raise embryos and larvae to expose them to the unique chemical cues during the window of imprinting while also implementing precautions to avoid pathogen introduction (e.g., Oidtmann et al. 2018).
The timing (i.e., fall during October) and release location might have also influenced the exit of sturgeon from the Maumee River due to water temperature and discharge. Downstream movements by multiple age classes of Lake Sturgeon are typically observed more frequently than upstream movements during water cooling periods (Caroffino et al. 2009, Morton et al. 2025) and periods of increased discharge (Tucker et al. 2022), possibly in search of warmer waters and greater food availability (Benson et al. 2005). Typically, regional rainfall is high during the fall in northwestern Ohio and Maumee River discharge starts to climb during this time (Culbertson et al. 2016). Elevated river discharge and water cooling surrounding the timing of stocking could have triggered the downstream movement we observed. Adding stocking sites upstream might aid a longer exposure to Maumee River water during periods of increased discharge and water cooling potentially facilitating imprinting and system learning.
Proximity to Lake Erie might have also influenced the exit of Lake Sturgeon from the Maumee River. Stocking occurred approximately 16 river kilometers from the mouth of the river due to the adjacent high-quality habitat (Collier et al. 2022). However, habitat below our stocking site was characterized as moderate to poor and made up of mostly silt and clay (Collier et al. 2022). Habitat preferences of Lake Sturgeon and juvenile sturgeon are variable across systems, with some studies suggesting sand and gravel substrates are preferred (Benson et al. 2005, Boase et al. 2014a, Hintz et al. 2015, Hintz et al. 2016), while others suggest silt and clay substrates may be important when sand and gravel are limited (e.g., Knights et al. 2002, Trested et al. 2011, Hrenchuk et al. 2017). Together these studies indicate the Maumee River might have more available suitable habitat for age-0 Lake Sturgeon than previously assumed. Adding release sites further upstream would increase exposure to the diversity of habitats along the cline of the Maumee River, potentially increasing river residence during a time when changing river conditions in the fall months could be triggering downstream movement. However, longer residence in the Maumee River is not without disadvantages, such as increased spatial and temporal overlap with predatory fishes (e.g., Flathead Catfish Pylodictis olivaris; Flowers et al. 2011), but additional data are needed to determine if predation is an issue affecting the survival of reintroduced Lake Sturgeon.
After leaving the Maumee River, the majority of tagged Lake Sturgeon were detected along the south shore of Lake Erie in U.S. waters of the western basin. While we cannot identify specific habitats used in this area, a couple of hypotheses merit further investigation. Within the detected area are reef structures. We do not know if these reefs provide suitable habitat for reintroduced Lake Sturgeon, but the reefs are important spawning grounds and provide beneficial habitat for many fishes of Lake Erie (Chiasson et al. 1997, Barth et al. 2009, Boase et al. 2014b, Manny et al. 2015, Roseman et al. 1996). Our tagged fish could have also simply followed the Maumee River water plume, which encompasses these reef structures (Niu and Xia 2021). Fish survival and growth during the early life stages can benefit from river plumes due to reductions in vulnerability to visual predators and higher food availability within the plumes (Reichert et al. 2010, Carreon-Martinez et al. 2014, Carreon-Martinez et al. 2015, Wishingrad et al. 2015). While it would be difficult to test whether reintroduced sturgeon used the rocky reefs or the Maumee River water plume given the spatiotemporal overlap of these two habitats, such an investigation could yield insights into why the majority of tagged fish spent time in this area of the western basin.
Some of the differences between the release years in detections, movements, and area use were likely due to changes in receiver arrays and coverage. In 2019, 25 additional receivers were added to the southern shore of the western basin of Lake Erie. This additional receiver coverage accounted for 20% of the Lake Sturgeon detections in 2019, which provided important data regarding movement and time spent in the river or lake for the 2019 and 2021 release years. The 25 receivers were not available in 2018, which might in part account for the sturgeon that were not detected in the lake. Current GLATOS arrays were also implemented to study large-bodied fishes typically implanted with larger, more powerful acoustic transmitters that have a detection radius of 800 to 1200 meters (Kraus et al. 2018). The transmitters used in our study have a detection radius of 300 to 400 m. The relatively sparse detection rates observed for Lake Sturgeon are not unique to our study, as other studies that focused on small-bodied fishes using smaller acoustic transmitters have also noted fewer detections (e.g., Cisco, Coregonus artedi) from the existing lake-wide array in Lake Erie (Kraus et al. 2024). Our ability to successfully detect fish relied on the sampling efforts occurring within the GLATOS network. We acknowledge that the ebb and flow of the number and location of receivers might have influenced our results, but we accounted for this by not generalizing conclusions among release years, only within release years. However, the grid design of the primary GLATOS array should reduce bias compared to other array designs (Kraus et al. 2018). As the reintroduction program continues, increased receiver coverage will aid our understanding of Lake Sturgeon recovery within Lake Erie. Additional receivers would increase detections and our resolution on movement behaviors, straying rates, habitat selection, and survival (e.g., see McKenna et al. 2025). All reintroduced Maumee River sturgeon have passive integrated transponder (PIT) tags for identification later in life. Opportunistically tagging these sturgeon as they become adults with long-lived acoustic tags combined with greater receiver coverage would help determine whether mature adults return to the Maumee River to spawn and rearing strategy facilitated imprinting of the returning spawners.
Our results should be interpreted considering the limitations of our study. First, our acoustic tags were programmed to maximize battery life and reduce the delay in transmission rate. While tag programming was sufficient to address our objectives, the 30-d pause restricted our ability to infer movement and habitat area use during this time, but also capture the effects of seasonality (e.g., Colborne et al. 2019, Izzo et al. 2022). It is possible that Lake Sturgeon moved between the lake or the river during the transmission pause. Knowing the detection history during this time could have also changed our residency estimates. Tags programmed without a long pause would likely reduce the uncertainty (confidence intervals) surrounding residency estimates. Second, we used the first and last detection rather than when the tag died to determine residency. Doing so could have introduced some measurement error. However, we used last detection rather than extrapolating residency to the estimated tag death date because this would have biased residency towards the habitat (lake vs. river) of last detection. Third, we cannot know the exact fate of all sturgeon that were not detected beyond the first week after release. Mortality, predation, movement upstream beyond our receiver locations in the river, or a lack of detection are all possible and are the focus of ongoing research. Ultimately, future work will need to consider the tradeoffs between battery life, transmission rate, and receiver coverage when using small transmitters. Predation transmitters would also add value to potentially understanding the unknown fates of some of the reintroduced Lake Sturgeon.
Conclusions
The goal of the Maumee River reintroduction program is to reach 1,500 naturally reproducing adult Lake Sturgeon in the river. Not until the late 2020s to early 2030s will we know if Lake Sturgeon have begun returning to the Maumee River to spawn (e.g., Welsh et al. 2019). Finding evidence of a self-sustaining Lake Sturgeon population will take longer than the current study. As the effort continues, it will be important to monitor the river for spawning adults, or if reintroduced sturgeon have strayed to colonize other systems. Passive-integrated transponder tags were implanted in all reintroduced Maumee River Lake Sturgeon. Efforts to sample these sturgeon as they age will be essential for evaluating project success and adaptive management. We discussed context for options that could be blended with ongoing operations to facilitate project success. The options include: 1) add stocking locations further upriver, 2) consider raising eggs and larvae on source water during the imprinting window or increase streamside rearing numbers if straying becomes an issue, and 3) increase GLATOS receiver coverage in the Maumee River and Lake Erie. These options might also serve other reintroduction efforts occurring throughout Lake Erie and other systems within the Lake Sturgeon’s native range.
Acknowledgements
We dedicate this work to the memory of Dr. Christopher Vandergoot, whose insights, mentorship, and dedication were instrumental in the conception of this research and development of this manuscript. His contributions to the field of fisheries science, aquatic biology, and his unwavering support for his colleagues are deeply missed. His absence is profoundly felt in our community. This work was funded by USGS Grant and Cooperative Agreement G20AC00447 (USFWS/USGS Science Support Program). This work was partially funded by the Great Lakes Fishery Commission (Grant ID #2013_BIN_44024) by way of Great Lakes Restoration Initiative appropriations (Grant ID #GL-00E23010). This paper is contribution 160 of the Great Lakes Acoustic Telemetry Observation System (GLATOS). The authors would like to thank Doug Aloisi, Orey Eckes, staff from the Genoa National Fish Hatchery, staff from the USFWS Alpena FWCO – Detroit River Substation: James Boase, Jennifer Johnson, Aaron Mettler, Paige Wigren, Jason Fischer, and the Toledo Zoo and Aquarium and staff: Megan Wozniak, Kaitlynn Zakcoczennyj, Jaiden Brandebury, Mariah Miller, Gus Harmon, Day’Veonna Smith, Hunter Sauerwine, Chris Otto, Felicity Galavan, Di-shon McCollough, Jane Driggs, Honesty Miller, Natalie Carlisle, Katie Herzog, and Kent Bekker. We also appreciate the veterinary staff at the Toledo Zoo: Dr. Genevieve Dumonceaux, Dr. Dennis Michels, Megan Pinder, and Brittany Exharos. The findings and conclusions of this article are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
References
Bade, A. P., T. R. Binder, M. D. Faust, C. S. Vandergoot, T. J. Hartman, R. T. Kraus, C. C. Krueger, and S. A. Ludsin. 2019. Sex-based differences in spawning behavior account for male-biased harvest in Lake Erie walleye (Sander vitreus). Canadian Journal of Fisheries and Aquatic Sciences 76:2003-2012.
Kessel, S. T., D. W. Hondorp, C. M. Holbrook, J. C. Boase, J. A. Chiotti, M. V. Thomas, T. C. Wills, E. F. Roseman, R. Drouin, and C. C. Krueger. 2018. Divergent migration within lake sturgeon (Acipenser fulvescens) populations: Multiple distinct patterns exist across an unrestricted migration corridor. Journal of Animal Ecology 87:259-273.
Krueger, C. C., C. M. Holbrook, T. R. Binder, C. S. Vandergoot, T. A. Hayden, D. W. Hondorp, N. Nate, K. Paige, S. C. Riley, A. T. Fisk, and S. J. Cooke. 2018. Acoustic telemetry observation systems: challenges encountered and overcome in the Laurentian Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 75:1755-1763.
Additional Information
Data availability: Data used in this study are openly available at https://doi.org/10.5066/P13YXIQY
Competing interests: The authors declare there are no competing interests.
Authors
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
McKenna, J.R., Chiotti, J.A., Vandergoot, C.S., Kraus, R., Faust, M.D., Slagle, Z.J., Weimer, E.J., Cross, M.D., and Hintz, W.D., 2026, Rearing method has limited effect on post-release movement of reintroduced age-0 Lake Sturgeon: Canadian Journal of Fisheries and Aquatic Sciences, v. 83, 13 p., https://doi.org/10.1139/cjfas-2025-0328.
Study Area
| Publication type | Article |
|---|---|
| Publication Subtype | Journal Article |
| Title | Rearing method has limited effect on post-release movement of reintroduced age-0 Lake Sturgeon |
| Series title | Canadian Journal of Fisheries and Aquatic Sciences |
| DOI | 10.1139/cjfas-2025-0328 |
| Volume | 83 |
| Publication Date | May 22, 2026 |
| Year Published | 2026 |
| Language | English |
| Publisher | Canadian Science Publishing |
| Contributing office(s) | Great Lakes Science Center |
| Description | 13 p. |
| Country | United States |
| State | Ohio |
| Other Geospatial | Lake Erie, Maumee River |