

Assessing the feasibility of reintroducing San Francisco gartersnakes (Thamnophis sirtalis tetrataenia) to La Honda Creek Open Space Preserve, San Mateo County, California
Links
- Document: Report (5.7 MB pdf) , HTML , XML
- Download citation as: RIS | Dublin Core
Acknowledgments
This work would not have been possible without the Midpeninsula Regional Open Space District, which has facilitated USGS research on San Francisco gartersnakes on their open space preserves. We thank many biological technicians who contributed to USGS studies of San Francisco gartersnakes for the past 15 years. We thank D. Wood and H. Waddle for providing thoughtful feedback that improved this report. Amphibian survey data from La Honda Creek Open Space Preserve used in this manuscript may be requested from Ariel Starr, Midpeninsula Regional Open Space District.
Abstract
Reintroductions are used worldwide to increase the viability of species and restore native ecological communities. The success of reintroductions is usually judged by the establishment of self-sustaining populations, restoration of naturally occurring ecological communities, and the species resuming its ecological function. Recovery for the endangered San Francisco gartersnake (SFGS, Thamnophis sirtalis tetrataenia), a subspecies with a small range in San Mateo and Santa Cruz counties in California, will likely require reintroduction and establishment of new populations within its historical range. La Honda Creek Open Space Preserve (LHC), managed by the Midpeninsula Regional Open Space District (MROSD), is one potential site for the reintroduction of SFGS. The La Honda Creek Open Space Preserve is a preserve managed for wildlife, recreation, grazing, and agriculture located near extant populations of SFGS inhabiting other open space preserves managed by MROSD (Cloverdale Ranch Open Space Preserve [CR]; Russian Ridge Open Space Preserve [RR]). We compared the habitat and prey communities at LHC to nearby open space preserves that support extant SFGS populations. Based on pond surveys done annually since 2008, the occurrence of California red-legged frogs (Rana draytonii), Sierran chorus frogs (Pseudacris sierra), and Pacific newts (Taricha spp.) at LHC indicates a similar prey community at this preserve to those at CR and RR. Likewise, the landscape at LHC is a similar mosaic of wetlands, open grassland, shrub-dominated scrub, and coast redwood (Sequoia sempervirens) and Douglas fir (Pseudotsuga menziesii) forest that meets the habitat requirements for the life history of SFGS at CR and RR. One difference between LHC and preserves with SFGS populations is the lack of vegetative cover immediately adjacent to some wetlands at LHC, which could affect the ability of SFGS to disperse from wetlands and find terrestrial refuges. To evaluate alternative reintroduction strategies, we simulated population viability for a fixed number of SFGS released at LHC into one to six subpopulations (where each wetland represents a subpopulation) over a period from 5 to 20 years. Population simulations indicated that the highest average viability (in other words, the lowest probability of quasi-extinction) occurred when all SFGS were released into a single subpopulation and releases continued annually for 15 to 20 years. Our results indicate that LHC is a good candidate for reintroducing SFGS with suitable habitat, climate, and prey for this snake subspecies. Supporting SFGS populations at LHC could require habitat management to provide sufficient vegetative cover in the terrestrial environment near wetlands. Maintaining genetic diversity in the reintroduced population will also be paramount to ensure negative effects of inbreeding and homozygosity do not affect population viability.
Introduction
Translocation of plants and animals can be used to achieve biodiversity conservation goals. When an organism is translocated to a currently unoccupied location within its indigenous range, it is called a reintroduction. Reintroductions are increasingly used to improve the viability of imperiled species (Taylor and others, 2017). As the number of attempted reintroductions grows, there are notable success stories (for example, black-footed ferrets [Mustela nigripes]; Jachowski and others, 2011) and high-profile reintroductions that have failed (for example, the Brown Treecreeper [Climacteris picumnus] in Australia; Bennett and others, 2012). Many studies have attempted to compile data on these efforts and ascertain what separates the successes from the failures to learn from and improve future reintroductions.
Reintroductions could fail if the proposed site is not abiotically and biotically suitable for the target species. The climatic suitability of a site is highly correlated to the probability of success for conservation translocations (Bellis and others, 2020). Unlike translocations outside a species’ historical range, climatic suitability might be less of a concern in the near term for reintroductions to formerly occupied sites. For reptiles, common factors that explain failed reintroductions include (1) poor survival during the inactive season (for example, overwintering mortality in temperate climates; Roe and others, 2010), (2) high dispersal-related mortality after release (Germano and Bishop, 2009), (3) low habitat quality at the reintroduction site (Germano and Bishop, 2009), and (4) decreased survival during the establishment phase compared to wild conspecifics (Bertolero and others, 2018). Feasibility studies that investigate if the ecological community can support the target species are vital to ensure the reintroduction does not waste valuable resources and expose individuals to a high risk of mortality in the new environment.
The San Francisco gartersnake (SFGS, Thamnophis sirtalis tetrataenia) is a subspecies of the common gartersnake listed as endangered under the U.S. Endangered Species Act (ESA; U.S. Fish and Wildlife Service, 1973, as amended) and the California Endangered Species Act (California Code of Regulations, 1971). San Francisco gartersnakes persist in 13 known populations in San Mateo County and northwestern Santa Cruz County, California (U.S. Fish and Wildlife Service, 2020). Many of these populations are geographically isolated, have low abundance and genetic diversity, and likely have limited or no genetic or demographic connectivity to other SFGS populations (Wood and others, 2020). The recovery plan for SFGS (U.S. Fish and Wildlife Service, 1985) calls for conservation actions to protect six extant populations known at the time and create four new populations. The recovery plan also calls for maintaining a minimum of 200 adult SFGS in a 1:1 sex ratio at each of 10 populations for at least 15 consecutive years for the species to be considered recovered (U.S. Fish and Wildlife Service, 1985). Most extant SFGS populations for which abundance has been estimated fall below this minimum threshold of 200 adults and have low genetic diversity (Wood and others, 2020). Given the fragmentation of habitat and existence of dispersal barriers (for example, roads, urban areas) within the historical range of SFGS, it is unlikely that snakes will be able to recolonize unoccupied but suitable habitat without human intervention. Considering the threats facing SFGS, reintroducing this subspecies to unoccupied suitable habitat within its historical range could be necessary to ensure its viability in the 21st century.
Despite the urbanization and development of the San Francisco Bay area since the 19th Century, large swaths of open space are protected in rural parts of San Mateo County, including state parks and several preserves managed by the Midpeninsula Regional Open Space District (MROSD), land trusts, and other nongovernmental organizations. San Francisco gartersnake populations exist in at least two preserves managed by MROSD: (1) Cloverdale Ranch Open Space Preserve (CR) near the coast in southwestern San Mateo County and (2) Russian Ridge Open Space Preserve (RR) further inland in the Santa Cruz Mountains (Halstead and others, 2011; Kim and others, 2018; Wood and others, 2020). It is possible that suitable habitat exists within other open space preserves managed by MROSD that is not occupied by SFGS at the time of this publication. The need to create additional populations of SFGS to increase the viability of this subspecies is made clear in the species’ recovery plan (U.S. Fish and Wildlife Service, 1985) and more recent species status assessment (U.S. Fish and Wildlife Service, 2020). One promising site for the reintroduction of SFGS to establish a new population is the La Honda Creek Open Space Preserve (LHC) owned by MROSD and in rural San Mateo County near the communities of La Honda and Woodside (fig. 1). The LHC is bordered to the south and east by State Highway 84 and to the north by Skyline Blvd (State Highway 35). The size of LHC, the wetland and upland habitats present, the ecological communities contained within, and the proximity to existing SFGS populations all combine to make this preserve a promising site for the reintroduction of SFGS.
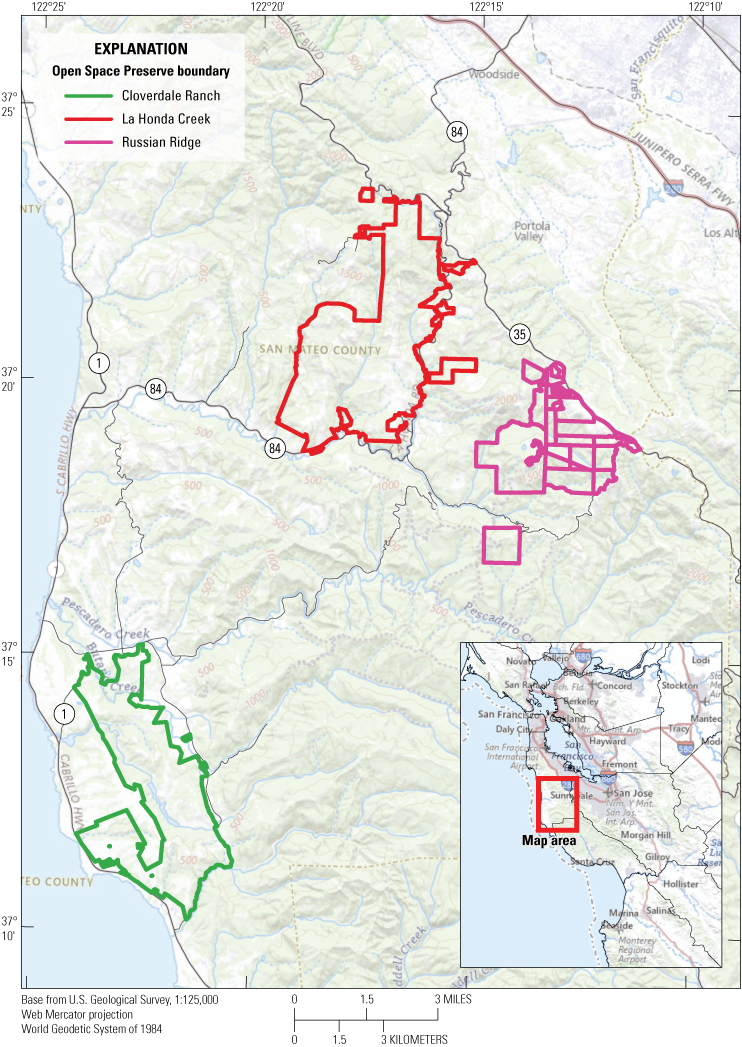
The location of Cloverdale Ranch Open Space Preserve (OSP), La Honda Creek Open Space Preserve, and Russian Ridge Open Space Preserve within San Mateo County, California. Russian Ridge OSP and Cloverdale Ranch OSP contain extant populations of San Francisco gartersnakes (Thamnophis sirtalis tetrataenia). Inset shows study area with the San Francisco Bay area.
We compiled data on the herpetofaunal community and habitat within LHC to evaluate if this preserve is abiotically and biotically suitable for the establishment of an SFGS population by comparing LHC to the CR and RR preserves that support extant SFGS populations. We then used a demographic model for SFGS to simulate the viability of reintroduced populations of SFGS at LHC under different scenarios to determine if introducing all snakes at one pond or dividing individuals among multiple ponds is more likely to result in a viable population. Finally, we discuss the data needed to monitor the status of reintroduced SFGS populations at LHC, evaluate if the reintroduction is successful, and take alternative actions if necessary.
Goals and Objectives
-
• Compile existing data on populations of SFGS prey species within LHC
-
• Compare prey communities and habitat at LHC to other preserves supporting SFGS populations
-
• Simulate reintroductions of SFGS to LHC to identify optimal strategies that result in the highest viability of reintroduced populations
Study Area
La Honda Creek Open Space Preserve is a 2,515 hectare (ha; 6,215 acre) preserve in San Mateo County, California, that encompasses historical ranches, farms, and timberlands (fig. 2). Elevations at LHC range from approximately 58 to 713 meters (m; 190 to 2,340 feet), with generally higher elevations in the northern half of the preserve and lower elevations in the southern half. La Honda Creek Open Space Preserve is managed to provide habitat for wildlife along with compatible ecological grazing and agricultural uses, hiking, equestrian, and bicycling trails for public recreation. Two Conservation Management Units also exist within LHC that are closed to the public. Most uplands at LHC are coniferous or evergreen hardwood forests with coast redwoods (Sequoia sempervirens) and Douglas fir (Pseudotsuga menziesii), followed by open grasslands and shrub-dominated scrub. Many wetlands are present in LHC, including more than 25 ponds and three main creeks with associated tributaries. Substantial parts of LHC were historically ranch land, and grazing is used as a management tool on four pastures within the preserve. Grazing is used to reduce fuel loads to mitigate fire risk and to manage grassland vegetation (Midpeninsula Regional Open Space District, 2012). One of the objectives of the LHC’s Master Plan is to “Protect and enhance populations of listed wildlife species” (Midpeninsula Regional Open Space District, 2012). One such listed species is the California red-legged frog (CRLF, Rana draytonii), which is listed as threatened under the U.S. Endangered Species Act (U.S. Fish and Wildlife Service, 1996) and breeds in several ponds at LHC (see “Prey Community at La Honda Creek Open Space Preserve”).
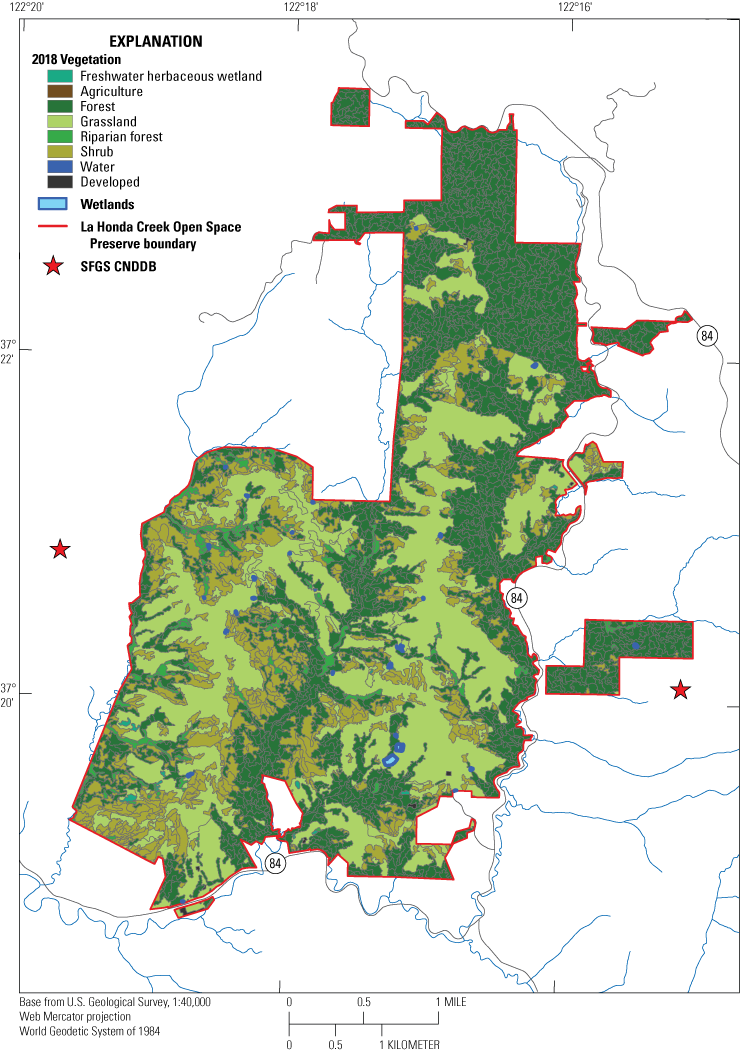
Land cover at La Honda Creek Open Space Preserve. Stars represent historical records of San Francisco gartersnakes (Thamnophis sirtalis tetrataenia) from the California Natural Diversity Database, originally observed by S.J. Barry in 1983 (Barry, 1994). Land cover data are from the 2018 San Mateo Countywide Fine Scale Vegetation Map (Golden Gate National Parks Conservancy and others, 2022). Abbreviations: CNDDB, California Natural Diversity Database; SFGS, San Francisco gartersnake.
San Francisco Gartersnake (Thamnophis sirtalis tetrataenia)
The San Francisco gartersnake is a subspecies of the common gartersnake (T. sirtalis) occurring on the San Francisco Peninsula and in the Santa Cruz Mountains. The extant distribution of SFGS is primarily in San Mateo County, with one population reported in northwestern Santa Cruz County (U.S. Fish and Wildlife Service, 2020). San Francisco gartersnakes are associated with wetlands surrounded by open grasslands and shrublands from the Pacific Coast to the Santa Cruz Mountains and low-lying habitats near the San Francisco Bay. San Francisco gartersnakes primarily forage in and around ponds, wetlands, and riparian areas for amphibians. Native anurans such as CRLF and Sierran chorus frogs (Pseudacris sierra) make up a large proportion of the diet of SFGS (Kim and others, 2021). Other prey for SFGS include Pacific newts (Taricha spp.), non-native American bullfrogs (Lithobates catesbeianus), and small fish (U.S. Fish and Wildlife Service, 2020). Sierran chorus frogs reach much smaller adult sizes than CRLF, but are widespread throughout California where they breed in a variety of wetland types and are often the most abundant amphibian at their breeding sites (Fisher and Shaffer, 1996; Halstead and others, 2023).
Methods
Suitability of the Ecological Community
We focused on presence and abundance of three prey taxa: (1) CRLF, (2) Sierran chorus frogs, and (3) Pacific newts (Taricha spp.). We analyzed egg mass counts and surveys for CRLF collected at LHC from 2008 to 2023 to determine the abundance of this prey species in ponds on site. We defined evidence of CRLF breeding based on the observation of CRLF egg masses, larvae, or recently metamorphosed juvenile frogs (that is, frogs that underwent metamorphosis in the year of the survey) at a wetland. We also searched through annual amphibian monitoring survey results at LHC for documentation of the occurrence of Sierran chorus frogs and newts at the preserve. Finally, although American bullfrogs are not established at LHC, their potential invasion was identified as a threat to wildlife communities in the preserve’s Master Plan (Midpeninsula Regional Open Space District, 2012). We compiled records on the occurrence of American bullfrogs (Global Biodiversity Information Facility, 2023a), Sierran chorus frogs (Global Biodiversity Information Facility, 2023b, c), and Pacific newts (Global Biodiversity Information Facility, 2023d) in the landscape surrounding LHC from the Global Biodiversity Information Facility (GBIF). The non-native American bullfrog is a potential threat to SFGS as a predator of young SFGS and a competitor for shared prey because bullfrog and SFGS diets overlap substantially where they co-occur (Kim and others, 2021).
We compared the herpetofaunal community at LHC to two nearby preserves that support SFGS populations: CR and RR. The Cloverdale Ranch Open Space Preserve is approximately 8.7 kilometers (km; 5.4 miles [mi]) southwest of LHC and encompasses 2,500 ha (6,200 acres) of herbaceous non-native grassland, coastal scrub, and evergreen and hardwood forest. The Russian Ridge Open Space Preserve is a 1,416 ha (3,500 acre) cattle ranch approximately 2.2 km (1.4 mi) to the southeast of LHC in the Santa Cruz Mountains, and similar to CR has a landscape of non-native grassland, scrub, and forest. We compiled occurrence data of CRLF at CR and RR from the California Natural Diversity Database (CNDDB). We compiled abundance estimates of American bullfrogs, CRLF, and Sierran chorus frogs at RR in 2014–15 from an earlier USGS study that used double-observer methods (Kim and others, 2018).
Simulating Reintroductions
We simulated reintroducing SFGS to LHC using a demographic population model developed and published by USGS biologists (Rose and others, 2023). Survival, growth, and fecundity of SFGS are size-dependent (Rose and others, 2022; Schoenig and others, 2023a). San Francisco gartersnakes grow rapidly when they are young, with snakes’ growth rate declining as individuals reach greater snout-vent lengths (SVL). Survival is highest for snakes of intermediate size (300–550 millimeters [mm] SVL) and decreases for larger adult females (550–700 mm SVL), with the survival rates of the largest (greater than 700 mm SVL) and smallest (less than 300 mm SVL) snakes highly uncertain based on available data (Rose and others, 2022). We incorporate that uncertainty in the model for our population simulations. The probability of reproduction and fecundity for female SFGS increase with size, with longer snakes more likely to be gravid and to give birth to larger litters of neonate snakes (Schoenig and others, 2023a). Details on the size-based demographic model for SFGS are available in Rose and others (2023), and a diagram representing the model is shown in figure 3. For consistency in our simulations, we assumed a neonate survival rate (from birth to 1 year old) of 0.30, which resulted in a slowly growing population in a stochastic environment in previous simulations for SFGS (Rose and others, 2023). We acknowledge that neonate survival could be higher or lower than this value in the wild, but fixing neonate survival to one value is sufficient for comparing relative outcomes among different reintroduction scenarios.
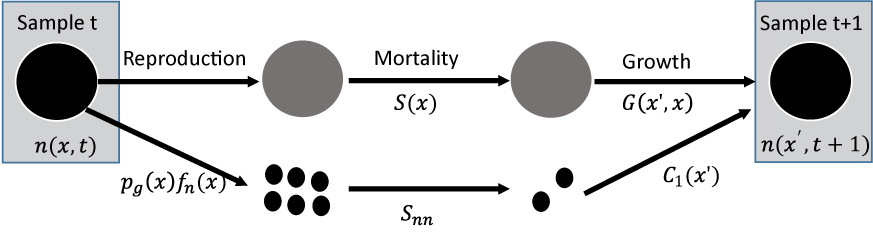
The San Francisco gartersnake (SFGS, Thamnophis sirtalis tetrataenia) population model used for simulating snake reintroductions. The model simulates the distribution of snake sizes (snout-vent length) in year t+1, n(xʹ,t+1), based on the distribution of snake sizes in year t, n(x,t). The function for SFGS survival is S(x), growth is G(xʹ,x), neonate survival is Snn, reproduction is the product of the probability a snake is gravid pg(x) and fecundity fn(x). The size of juvenile recruits when they join the sampled population at 1-year-old is C1(xʹ). Figure reproduced from Rose and others (2023).
Based on the results of Rose and others (2023), we simulated reintroduced populations of SFGS in which head-started, 1-year-old juvenile snakes are released to the wild. In head-starting, gravid adult female snakes are temporarily removed from donor populations and held in captivity until they give birth. The adult female snakes are then returned to the site from which they were collected. The neonate snakes are raised in captivity before being released to the wild at an age and size expected to increase their chances of survival (Nguyen and others, 2023). Head-starting provides reptiles the opportunity for increased growth and survival compared to neonates born in the wild (Tuberville and others, 2019). Previous population simulations indicated that releasing head-started juvenile snakes resulted in the greatest viability of reintroduced and source populations (Rose and others, 2023). Our population model and simulations only track female SFGS, but we assume that an equal number of female and male juvenile SFGS are released on average.
For the purposes of simulation, we considered each wetland to represent a potential site for a subpopulation of SFGS. We simulated six scenarios for reintroduction, with the number of distinct subpopulations at which snakes were released varying from one to six. We did not model connectivity and dispersal of snakes among subpopulations. Instead, we modeled each subpopulation in isolation. Treating each subpopulation as distinct likely produces conservative estimates of quasi-extinction risk because dispersal of snakes from a nearby subpopulation could “rescue” a declining subpopulation from extinction (Brown and Kodric-Brown, 1977). The total number of head-started female juvenile SFGS released each year was held constant at 36, to limit the number of adults required to produce juveniles each year to 10 adult females. This number of adult females required to produce enough neonates for rearing and release, assuming a 1:1 sex ratio among neonates, was estimated in an earlier study on SFGS reintroduction strategies and is intended to minimize effects on SFGS donor populations (Rose and others, 2023). The number of individuals was divided evenly among subpopulations. For example, if 36 total juvenile SFGS were released in each scenario, this would be 36 SFGS at 1 subpopulation, 18 SFGS per subpopulation for 2 subpopulations, or 12 SFGS per subpopulation for 3 subpopulations (table 1). For the scenario with 5 subpopulations, 4 subpopulations had 7 SFGS released each year and the 5th subpopulation had 8 SFGS released each year, a total of 36 SFGS released annually. The scenarios were run under four durations of reintroduction efforts. An equal number of SFGS were reintroduced annually for 5, 10, 15, or 20 years. The size of juvenile SFGS released was drawn from a normal distribution based on the mean (338.9 mm) and standard deviation (46.4 mm) of expected SVL for 1-year-old juvenile SFGS from a von Bertalanffy growth function fitted to capture-recapture data (Rose and others, 2023). It is possible that head-started SFGS reared in captivity could reach larger sizes at 1 year old, but without data on growth in captivity, we made the conservative assumption that captive-reared snakes would be a similar size to those in wild populations.
Table 1.
Results from reintroduction scenarios for San Francisco gartersnakes (Thamnophis sirtalis tetrataenia) at La Honda Creek Open Space Preserve.[Subpopn, subpopulation; Mean pqext, mean probability of quasi-extinction during the 30-year simulation for subpopulations in that scenario; Mean N30, mean abundance of SFGS after 30 years for all simulations; Mean tqext, mean time to quasi-extinction for simulations that go quasi-extinct in that scenario]
For each scenario, we ran 1,000 simulations with different parameter values for SFGS growth, survival, and fecundity to reflect parametric uncertainty (in other words, uncertainty about the true value of growth, survival, and reproductive rates) and environmental stochasticity (in other words, random annual variation in survival and growth rates). Demographic stochasticity, or randomness in the birth and death of individuals, is an important factor in the probability of extinction for small populations (Caughley, 1994; Melbourne and Hastings, 2008). To account for demographic stochasticity, we simulated reproduction and survival for each individual SFGS using random draws from Poisson and Bernoulli distributions, respectively. We ran each simulation for 30 years and calculated the number of SFGS surviving in the final year. We defined a quasi-extinction threshold of five individuals. If the population of SFGS was less than or equal to five individuals at any point during the simulation, it was considered to have gone extinct and the simulation stopped. We recorded the year in which the subpopulation went extinct for each simulation and calculated the average time to quasi-extinction for each scenario in which the subpopulation went quasi-extinct (in other words, simulations in which the subpopulation did not go quasi-extinct after 30 years did not factor into the average time to quasi-extinction calculation). Finally, we calculated the quasi-extinction probability using the proportion of the 1,000 simulations in which each subpopulation went extinct. For scenarios with greater than one subpopulation, we calculated the quasi-extinction probability for each individual subpopulation.
Demographic population models are, by design, a simplification of the real dynamics of populations. Predictions from demographic population models can be inaccurate because of several factors that exist in nature and are not captured by the model. Rather than focus on the absolute quasi-extinction probability predicted from any single simulation, we use the results of the population simulations to compare the viability and abundance of populations among different scenarios (McCarthy and others, 2003).
Results
Records of SFGS from 1983 reported by Barry (1994) were within 1.5 km (0.9 mi) of the eastern and western borders of LHC (fig. 2). These nearby occurrences, combined with the existing population of SFGS at nearby RR 3.7 km (2.3 mi) to the southeast, and the number of wetlands present at LHC, indicate it is likely that SFGS historically occurred within the present-day boundary of LHC. If SFGS previously occupied LHC, the cause of the species’ putative extirpation is unknown. One possible explanation for the extirpation of SFGS is that incompatible land uses (for example, intensive grazing, agriculture, or forestry) negatively affected aquatic and terrestrial habitats and prey populations.
San Francisco gartersnake populations occur in cooler, fog-influenced coastal regions, and warmer inland regions of San Mateo County (U.S. Fish and Wildlife Service, 2020). The LHC is likely suitable climatically for SFGS given its proximity to the extant population at RR and the more coastal population at CR. Mean total annual precipitation at LHC from 1991 to 2020 averaged 800 mm, slightly lower than the 924 mm at RR, but higher than the average precipitation of 680 mm at CR (PRISM Climate Group, 2022). The climatic effect of the Pacific Ocean, approximately 7 km (4.4 mi) west of the western edge of LHC, moderates temperatures in this region, limiting extreme low temperatures in the winter and high temperatures in the summer.
Climate change could affect the suitability of LHC for SFGS in the future, and these effects will likely be similar to those at other sites occupied by SFGS. Increased variability in precipitation in the 21st Century could increase the risk of droughts, wildfires, and heavy rainfall events (Diffenbaugh and others, 2015; Swain and others, 2018; Gershunov and others, 2019; Swain, 2021). Long-term droughts could negatively affect amphibian populations (Moss and others, 2021) that are prey for SFGS. The potential effects of wildfires on SFGS are more complex. Although wildfires could cause direct mortality and disturb habitat occupied by SFGS, fires could prevent succession and maintain the open grassland and scrub habitats used by SFGS. A study of an SFGS population after a controlled burn indicated that the survival probability of SFGS in burned areas was likely unchanged after the fire (Halstead and others, 2019). The effects of heavy rainfall and flooding on SFGS populations are unknown. Heavy rainfall can stimulate breeding by amphibians (Jensen and others, 2003), producing abundant prey for semiaquatic snakes in wet years. Overall, the volatile interannual fluctuations in rainfall predicted by climate models represents increased environmental stochasticity, which, in general, is expected to increase extinction probability for populations (Caughley, 1994; Doak and others, 2005). Given the limited geographic range of SFGS, the risks posed by climate change are likely to affect most existing SFGS populations, and suitable but unoccupied habitats, in similar ways.
Overall, the habitat present at LHC is similar to areas occupied by SFGS populations in the Santa Cruz Mountains, with one notable exception regarding terrestrial habitat immediately adjacent to wetlands at LHC (see next paragraph). The LHC has more than 25 wetlands, including natural, improved, and constructed ponds. Some wetlands are permanent and hold water year-round, and some are seasonal, filling with water during winter and spring rain and drying during the summer and autumn. The wetlands at LHC range from 0.01 ha (0.03 acres) to 1.03 ha (2.55 acres) in surface area. The Midpeninsula Regional Open Space District has improved several wetlands in the past 15 years to provide better habitat for native amphibians, and has developed a plan to make wetlands suitable for SFGS should the subspecies be reintroduced (Midpeninsula Regional Open Space District, 2012). The LHC has several wetlands with emergent vegetation that are used for breeding by native amphibians (see “Prey Community at La Honda Creek Open Space Preserve”). These wetlands are adjacent to upland grassland and scrub (fig. 2) that could be used for dispersal or refuge during inactive periods.
Two key requirements for SFGS habitat beyond wetlands are (1) the presence of mammal burrows and other subterranean refuges and (2) emergent vegetation and shrubs along the edge of wetlands. Mammal burrows and subterranean refuges provide stable temperatures and moisture for snakes to overwinter during colder months, aestivate during summer heat, and avoid predators. The overwintering habitat of SFGS is not well-studied, but it is thought that snakes move into upland habitat during the autumn and take refuge underground in mammal burrows during late autumn and winter before returning to foraging habitat in the spring (U.S. Fish and Wildlife Service, 2020). The proximity of shrub-covered hillsides adjacent to wetlands could be important for allowing SFGS to migrate from foraging grounds used in the spring and summer to refuges used during the fall and winter months and for promoting movement between wetlands. On a shorter time scale, snakes need to retreat from foraging habitats to gestate their young, digest their prey, and shed their skin. During these activities, snakes are vulnerable to predators and take cover underground or under thick vegetation, emerging to bask near these refuges. One outstanding question regarding refuges for SFGS at LHC is whether sufficient mammal burrows and other underground structures are present near wetlands. The historical agricultural land use in LHC has resulted in decreased abundance of small mammal burrows in some areas (J. Andersen, Midpeninsula Regional Open Space District, written commun., 2023). A field visit by a USGS biologist (E. Schoenig) in November 2023 confirmed that small mammal burrows are present in the uplands near wetlands used by amphibians for breeding at LHC. The fact that many other snake species occur at LHC, including other gartersnakes (see below), indicates that sufficient burrows and other upland refuges are present at LHC for SFGS.
The second key requirement for SFGS terrestrial habitat is cover immediately adjacent to wetlands provided by emergent wetland vegetation (for example, cattails [Typha spp.], tules [Scheonoplectus acutus]), riparian trees (for example, willows [Salix spp.]), and scrub (for example, coyote bush [Baccharis pilularis], blackberry [Rubus spp.]) further upland. Several ponds at LHC are adjacent to coyote bush scrub that could provide cover and refuge for SFGS during their inactive periods, and willow and tule stands grow along some pond edges. Field surveys of wetlands at LHC by USGS in November 2023 indicated that at many water bodies, there was little vegetative cover near the water’s edge. Grazing by cattle in and around wetlands results in only short herbaceous cover that is avoided by SFGS, and direct grazing on emergent vegetation minimizes ideal foraging area. Both types of grazing were evident at some LHC wetlands along with heavy traffic by cattle that can collapse burrows and cause compacted ground in the immediate wetland vicinity. As a result, some aquatic features at LHC are surrounded by a ring of bare ground that creates a deterrence and potential risk for snakes moving between their foraging habitat and upland refugia. The habitat surrounding wetlands could be made more suitable for SFGS if the access of cattle to most parts of the water body were limited by exclosures that buffered the water’s edge and extended to the adjacent upland areas to provide protected connectivity between foraging and upland habitats. The value of buffers that limit land use affecting surrounding wetlands has been demonstrated for many amphibians and semiaquatic reptiles (Burke and Gibbons, 1995; Semlitsch and Bodie, 2003).
The community of snakes that occurs at LHC also is relevant for evaluating the suitability of the preserve for SFGS. All the snake species known to occur at CR, except for SFGS, have been present at LHC, along with one additional species (forest sharp-tailed snake, Contia longicauda) that also is present at RR (Amanda Sparkman, Westmont College, written commun., 2023). These include racer (Coluber constrictor), gophersnake (Pituophis catenifer), ring-necked snake (Diadophis punctatus), rubber boa (Charina bottae), and northern Pacific rattlesnake (Crotalus oreganus oreganus), and two other gartersnakes: Santa Cruz gartersnake (T. atratus atratus) and coast gartersnake (T. elegans terrestris). Given that SFGS co-occur with these other snake species in many of the remaining SFGS populations, it is likely that the SFGS could coexist with the snake community at LHC.
Prey Community at La Honda Creek Open Space Preserve
The MROSD has done extensive surveys for CRLF breeding at several wetlands within LHC since 2008, with the most complete data available from 2016 to 2023. Not all wetlands are monitored, and the counts that follow likely represent a subset of the total number of wetlands used for breeding by CRLF at LHC. From 2016 to 2023, the number of ponds at LHC with evidence of CRLF breeding ranged from a minimum of 5 wetlands in 2022 to a maximum of 11 wetlands in 2019. In total, CRLF breeding has been documented in 13 ponds on at least one occasion from 2016 to 2023 (fig. 4). Ponds Driscoll Ranch (DR)03, DR04, DR05, DR06, DR07, and DR08 had the most consistent evidence of CRLF breeding from 2016 to 2023 based on egg mass counts (table 2), larval counts (table 3), and counts of recently emerged metamorphs (table 4). Five of these six ponds (all but DR03) are within an approximately 1-km radius of one another in the center of LHC, to the east and west of Harrington Creek (fig. 4). The intervening habitat between these five central ponds is largely grassland (herbaceous cover) and scrub, with forest surrounding Harrington Creek and its tributaries. In contrast, pond DR03 is near the eastern edge of LHC, approximately 300 m west of State Highway 84. In addition, CRLF breeding is presumed to occur in DR19, DR20, and DR26 each year, but these wetlands are not regularly surveyed (table 2; J. Chung, Midpeninsula Regional Open Space District, written commun., 2023).
Table 2.
Summary of California red-legged frog egg mass counts at La Honda Creek Open Space Preserve from 2008 to 2023.[Unpublished data (not publicly available at the time of publication) were used with permission from the Midpeninsula Regional Open Space District on November 25, 2024. Abbreviations: DR, Driscoll Ranch; NS, site was not sampled for egg masses that year; —, no data; LHC, La Honda Creek Open Space Preserve]
Table 3.
Summary of California red-legged frog larvae counts at La Honda Creek Open Space Preserve from 2008 to 2023.[Unpublished data (not publicly available at the time of publication) were used with permission from the Midpeninsula Regional Open Space District on November 25, 2024. Abbreviations: DR, Driscoll Ranch; NS, site was not sampled for larvae that year; Y, site had larvae but no count is available; LHC, La Honda Creek Open Space Preserve]
Table 4.
Summary of California red-legged frog metamorph counts at La Honda Creek Open Space Preserve from 2008 to 2023.[Unpublished data (not publicly available at the time of publication) were used with permission from the Midpeninsula Regional Open Space District on November 25, 2024. Abbreviations: DR, Driscoll Ranch; NS, site was not sampled for metamorphs that year; Y, site had California red-legged frog metamorphs but no count is available; —, no data; LHC, La Honda Creek Open Space Preserve]
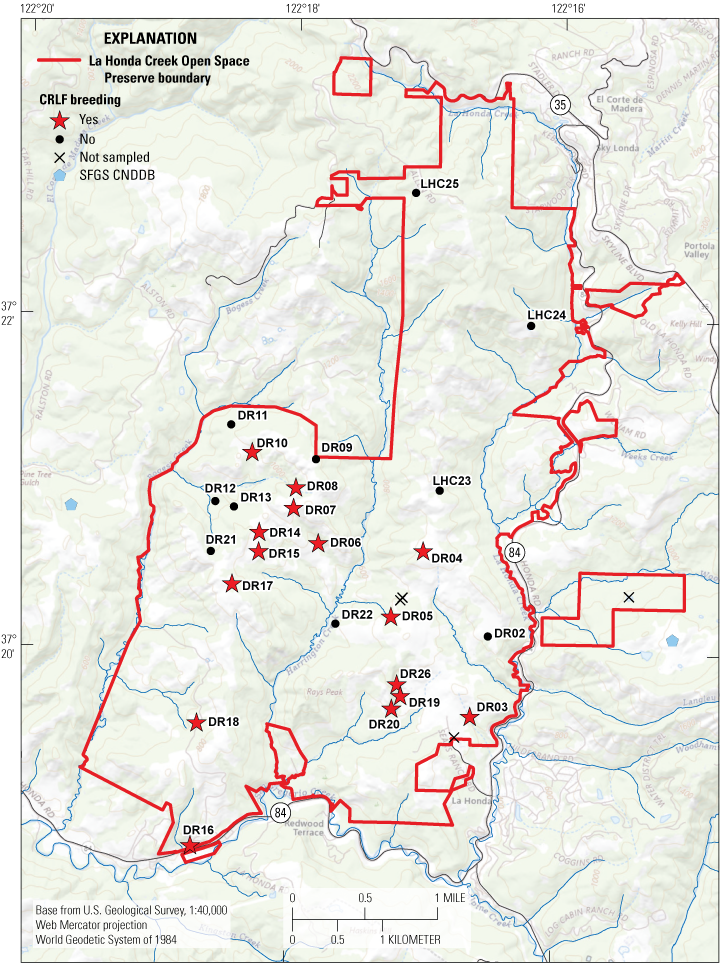
Sites with documented breeding by California red-legged frog (CRLF) at La Honda Creek Open Space Preserve from 2016 to 2023. Documented breeding was determined by the presence of egg masses, larvae, or recently metamorphosed juvenile CRLF at a pond during surveys by Midpeninsula Regional Open Space District in any year from 2016 to 2023. Unpublished data (not publicly available at the time of publication) were used with permission from the Midpeninsula Regional Open Space District on November 25, 2024. Abbreviations: CNDDB, California Natural Diversity Database; DR, Driscoll Ranch; LHC, La Honda Creek Open Space Preserve; SFGS, San Francisco gartersnake.
Although Sierran chorus frogs and Pacific newts were not the focus of amphibian breeding surveys at LHC, incidental observations of egg masses, larvae, and metamorphs of each were recorded during CRLF surveys. In addition, many observations of Sierran chorus frogs have been recorded from the southern half of LHC in the GBIF database (fig. 5; Global Biodiversity Information Facility, 2023b, c) and Pacific newts have been observed at wetlands within LHC during surveys by MROSD staff and throughout the Santa Cruz Mountains surrounding LHC based on GBIF data (fig. 5; Global Biodiversity Information Facility, 2023d).
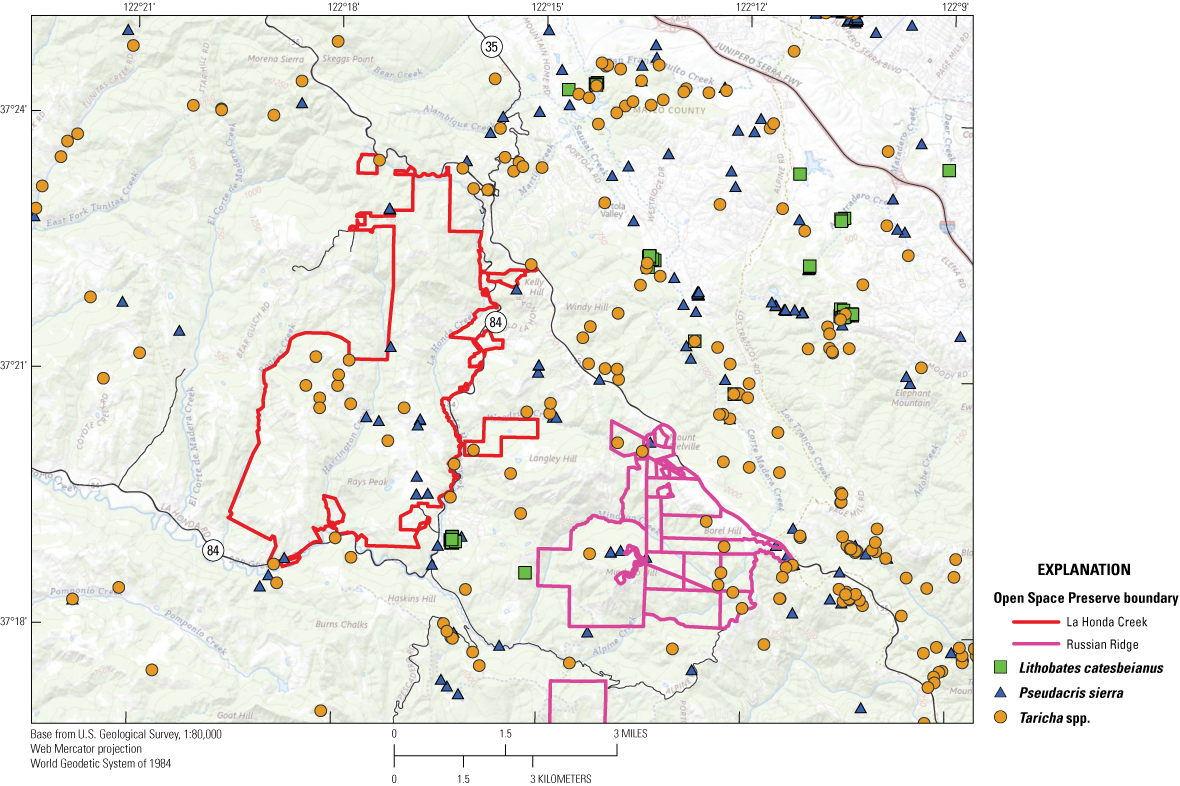
The location of American bullfrog (Lithobates catesbeianus), Sierran chorus frog (Pseudacris sierra), and Pacific newt (Taricha spp.) occurrences from 2000 to 2023 in the Global Biodiversity Information Facility database (Global Biodiversity Information Facility, 2023a, b, c, d).
Non-native American bullfrogs are not yet known to have established a breeding population at LHC. A small number of bullfrogs have been detected during amphibian surveys completed from 2008 to 2023 and removed to limit their spread into the preserve. American bullfrogs have been captured at pond DR16 and likely dispersed to this pond from the main stem of San Gregorio Creek (J. Chung, Midpeninsula Regional Open Space District, written commun., 2023). There are records of American bullfrogs from the landscape surrounding LHC, with the closest known records being a cluster of American bullfrogs recorded from 2020 to 2023 in a wetland less than 1 km east of the eastern boundary of LHC (Global Biodiversity Information Facility, 2023a; fig. 5). This cluster of American bullfrog occurrences is separated from LHC by State Highway 84, but nevertheless is a potential source for this non-native species to disperse into LHC. Based on GBIF data, another American bullfrog population seems to exist in the Windy Hill Open Space Preserve, approximately 3 km east of the northeastern boundary of LHC (fig. 5).
Comparison of Habitat and Prey Community at La Honda Creek Open Space Preserve to Cloverdale Ranch Open Space Preserve and Russian Ridge Open Space Preserve
The habitat at RR includes four water bodies with emergent vegetation, all of which are used by SFGS. The four water bodies range in size from approximately 0.1 to 1.3 ha (0.2 to 3.1 acres). One water body is a permanent lake that contains water year-round, except during extreme droughts (Kim and others, 2018). Non-native American bullfrogs previously occupied the permanent lake at RR, and eradication efforts took place in 2014 and 2015 to remove bullfrogs. The estimated adult bullfrog abundance at this lake was 16 individuals in 2014 and only 3 individuals in 2015 (Kim and others, 2018). Culling adult bullfrogs, combined with drought that caused the lake to dry up in 2014 and kill bullfrog larvae, likely extirpated bullfrogs from RR at least temporarily, with no bullfrogs detected in surveys done by USGS in 2016 and 2017 (Kim and others, 2018) or by MROSD during annual amphibian surveys from 2018 to 2023 (Matt Chaney, Midpeninsula Regional Open Space District, written commun., November 6, 2023). The existence of American bullfrogs near the boundary of RR (fig. 5) highlights the risk that this non-native species could reinvade RR in the future. Sierran chorus frogs were the most abundant amphibian at RR during surveys in 2014 and 2015, with more than 5,000 adult chorus frogs estimated to occur in the permanent water body in the spring of 2014 and more than 1,000 in the spring of 2015. Abundance estimates of adult CRLF were much lower, with only a mean estimate of 10 adult CRLF at the permanent water body in 2015. Notably, more than 20,000 recently metamorphosed CRLF were estimated at the permanent water body at RR in late summer 2015, indicating the potential for recruitment into the CRLF population (Kim and others, 2018). No updated estimates of CRLF abundance are available at RR after 2015, although annual counts of egg masses, larvae, and metamorphs have continued. Amphibian surveys by MROSD staff from 2018 to 2023 confirmed the continuing existence of CRLF populations at RR. Evidence of CRLF breeding was observed in at least two of four water bodies at RR from 2019 to 2023 (Matt Chaney, Midpeninsula Regional Open Space District, written commun., November 6, 2023).
Pacific newts were also captured near water bodies during USGS amphibian surveys at RR in 2015 and 2016, but no abundance estimates could be calculated with the data available (Kim and others, 2018). Pacific newts have been observed regularly at RR from 2018 to 2023, indicating the continued presence of this SFGS prey in and around water bodies at RR.
The Midpeninsula Regional Open Space District performed pond enhancement projects at three water bodies in RR in 2020. These projects included installing or reinforcing cattle fences around the water bodies, repairing the berm to prevent leakage, and dredging ephemeral ponds. These management implementations effectively reduced cattle effects on all three water bodies. Furthermore, these actions extended the hydroperiod and allowed establishment of dense emergent vegetation for two ephemeral ponds. Before these management actions, CRLF egg masses were not observed in one of the ephemeral ponds, and recruitment of CRLF metamorphs were not observed at the other ephemeral pond, despite the presence of egg masses. After these management actions in 2020, CRLF metamorphs were observed at both ephemeral water bodies during visual encounter surveys between 2021 and 2023 (Matt Chaney, Midpeninsula Regional Open Space District, written communication, November 6, 2023).
A potential factor that might have bolstered the outcomes of pond enhancement and the recruitment of CRLF is the “conservation grazing regime” implemented at RR. This grazing regime maintained the cattle density at RR under a management threshold, which aimed to dampen grazing pressure (J. Andersen, Midpeninsula Regional Open Space District, written commun., 2023).
The CR preserve has 24 lentic water bodies, ranging from small ephemeral wetlands covering less than 0.2 ha (0.5 acres) to the larger Lake Lucerne (14.3 ha or 35.3 acres) and upper (10.2 ha or 25.1 acres) and lower (13 ha or 32 acres) Bean Hollow Reservoirs. San Francisco gartersnakes occur in at least 15 aquatic features at CR, but have been reported most often around permanent stock ponds with ample emergent vegetation and adjacent upland habitat containing scrub and riparian woodland (Midpeninsula Regional Open Space District, 2024). Seasonal water features that dry out in the late summer or fall are also used, especially when scrub or wet meadow habitat directly connects them to more permanent aquatic features. Like LHC, CR is an actively grazed ranch. Cattle exclosures surround many of the smaller discrete stock ponds, and limited grazing directly in CR’s aquatic features has encouraged dense emergent vegetation growth and allowed woody perennials to become established and form upland habitat where snakes can bask and shelter directly adjacent to aquatic features. The presence of extensive riparian woodland and coastal scrub directly surrounding aquatic features at CR is an important habitat that was limited at LHC during our field surveys.
Biologists from the USGS have observed CRLF near at least 18 water bodies, creeks, or wetlands while trapping and surveying for SFGS at CR including all but two of the aquatic features where SFGS have been present (Schoenig and others, 2023b). The USGS surveys have documented Sierran chorus frogs breeding in several wetlands, often near funnel traps that frequently catch SFGS (Schoenig and others, 2023b). Likewise, Pacific newts have been documented breeding in several wetlands at CR and have also been captured in funnel traps in areas sampled for SFGS (Schoenig and others, 2023b). Non-native American bullfrogs are known to occur alongside SFGS in at least three wetlands in the northern quarter of CR, but they have not been documented at long-term USGS study sites in the southern half of CR (Schoenig and others, 2023b).
Potential Source Populations for San Francisco Gartersnakes to Reintroduce to La Honda Creek Open Space Preserve
The CR (referred to as Pescadero in Wood and others, 2020) and RR (Mindego in Wood and others, 2020) populations of SFGS fall within the southern genetic cluster for this subspecies, along with the population at Año Nuevo State Park (Wood and others, 2020). There is some evidence of genetic mixing between CR and RR populations, with one genotype collected from CR classified as more closely related to snakes from RR than other snakes from CR (Wood and others, 2020). The genetic effective population size at CR (Ne=60) was higher than the effective population size at RR (Ne=33), indicating lower genetic diversity and a greater risk of inbreeding depression at RR (Wood and others, 2020). San Francisco gartersnakes at Año Nuevo State Park represent another potential source population for translocating snakes to LHC. Genotyped SFGS from Año Nuevo State Park were closely related to snakes from CR and RR and were also within the southern genetic cluster (Wood and others, 2020). The Año Nuevo population of SFGS had similar genetic effective population size (Ne=40) as populations at CR and RR. Notably, the Año Nuevo SFGS population showed increased diversity over time when comparing samples collected from 2004 to 2010 and 2016 to 2018 (Wood and others, 2020). Some SFGS sequenced at Año Nuevo were admixed with SFGS from CR (Wood and others, 2020), indicating genetic connectivity between these populations. Given the low genetic diversity of SFGS in some populations in the southern genetic cluster, consideration could also be given to translocating snakes from populations in the northern genetic cluster to LHC. Admixture of snakes from the northern and southern genetic clusters could lead to increased heterozygosity in the reintroduced population, as was shown in Alpine ibex (Capra ibex) reintroductions (Biebach and Keller, 2012). Admixing individuals from multiple populations can increase the probability of a successful reintroduction while minimizing the effect of harvesting individuals from small extant populations with low genetic diversity (Onley and others, 2023). One concern with admixing individuals from multiple populations is the possibility of swamping out local adaptation by introducing individuals from different environments (Shi and others, 2018). Given the narrow geographic range of SFGS, maintaining local adaption could be less of a concern than increasing genetic diversity within small populations.
Population Simulations for San Francisco Gartersnake Populations at La Honda Creek Open Space Preserve
The highest viability for reintroduced SFGS subpopulations at LHC occurred when the duration of reintroduction was greater than or equal to 15 years, and all juvenile snakes were released in a single subpopulation. Dividing the propagule of SFGS juveniles into more subpopulations led to a higher quasi-extinction probability for each subpopulation (fig. 6). When all juvenile SFGS were released as part of a single subpopulation, mean abundance of SFGS per subpopulation at the end of the 30-year simulation was higher than if the same total number of SFGS juveniles were divided up and released into more than one subpopulation (fig. 7). Quasi-extinction probability was much higher when annual reintroductions of SFGS juveniles only took place for 5 or 10 years. A 20-year reintroduction resulted in lower quasi-extinction probabilities and higher abundance of SFGS after 30 years, but the difference between a 15-year or 20-year long reintroduction was smaller, indicating diminishing returns with increasing duration after a certain length of time.
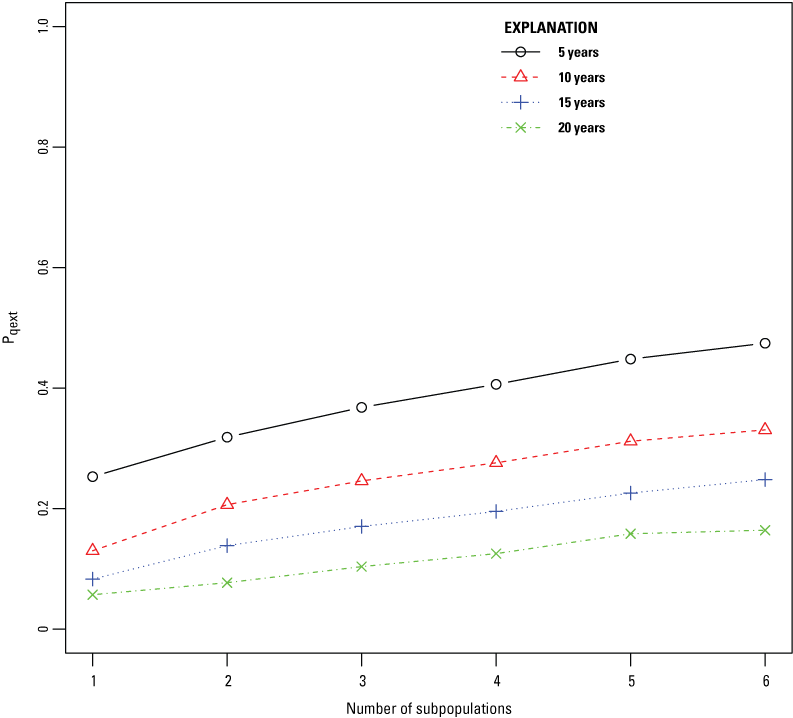
Mean quasi-extinction probability of San Francisco gartersnakes (pqext; Thamnophis sirtalis tetrataenia) as a function of the number of subpopulations that juvenile snakes were released into. The mean quasi-extinction probability is the average over all subpopulations in that scenario. Line color, style, and symbol indicate the duration of the reintroduction effort.
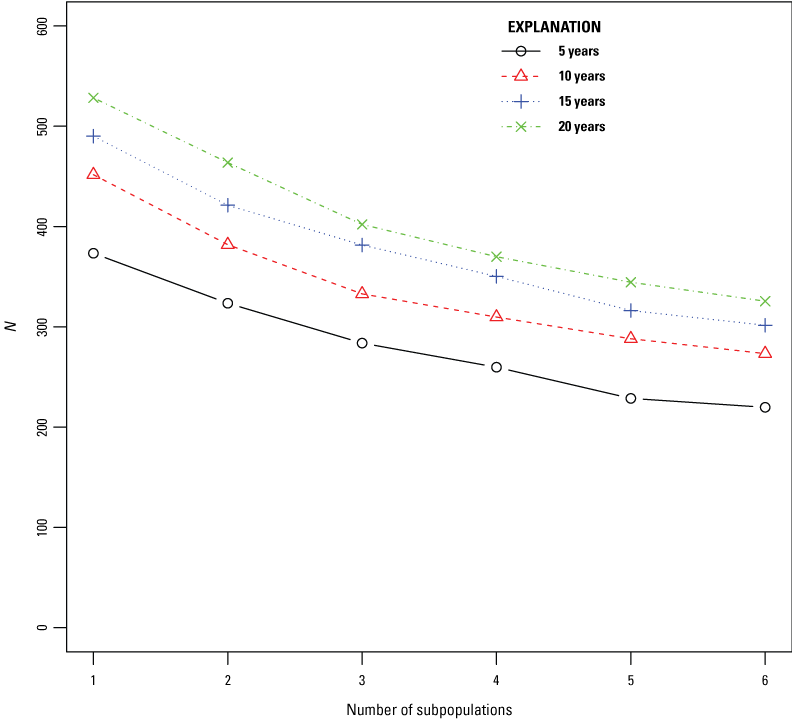
Mean abundance of San Francisco gartersnakes (Thamnophis sirtalis tetrataenia) at the end of the 30-year simulation (N) versus the number of subpopulations that juvenile snakes were released into. The mean abundance is averaged over all subpopulations in that scenario. Line color, style, and symbol indicate the duration of the reintroduction effort.
Discussion
This report has focused on the suitability of LHC for reintroducing SFGS and the best approach to translocating captive-reared juvenile snakes to their new home. Following up reintroduction efforts by monitoring the reintroduced population is essential to evaluate the success or failure of the endeavor (Ewen and Armstrong, 2007). As an active management intervention, reintroductions are ideally suited to an adaptive management approach in which data are continually analyzed and decisions reconsidered in the light of new evidence (Runge, 2013). Strategies for collecting data from a reintroduced SFGS population can follow the template used at existing populations. Trapping methods for SFGS are well-established, with drift fences and funnel traps proving effective at capturing juvenile and adult snakes (Halstead and others, 2011; Reeder and others, 2015; Kim and others, 2018). The selection of state variables to measure is an important first step in monitoring a reintroduction program (Nichols and Armstrong, 2012). The abundance of SFGS, and the survival, growth, and fecundity of individual snakes are important metrics for determining population viability (Wood and others, 2020; Rose and others, 2022, 2023). Capture-mark-recapture (CMR) methods, in which individual animals are marked such that they can be identified if recaptured, are preferred to using count data from unmarked animals for monitoring reintroductions (Nichols and Armstrong, 2012).
Capture-mark-recapture methods have been used to estimate the abundance of SFGS (Kim and others, 2018), and CMR data can be used to estimate survival and growth rates of marked SFGS if studies continue for 3 or more years (Rose and others, 2022). Based on prior CMR studies, drift fences with funnel traps could be deployed around the focal wetland(s) at which SFGS are released to capture SFGS moving to and from their foraging habitat. Trapping during the spring (April and May) for approximately 8 weeks has provided adequate numbers of recaptures of marked SFGS to estimate abundance using closed CMR models (Wood and others, 2020). Sampling during the spring also enables researchers to determine the reproductive status and fecundity of female SFGS by using a portable ultrasound to scan for the presence and number of developing embryos (Rose and others, 2023; Schoenig and others, 2023a). Capture-mark-recapture trapping would enable monitoring of the sex ratio of adult snakes, which is a key population metric listed in the species status assessment for SFGS (U.S. Fish and Wildlife Service, 2020).
Estimating growth and survival of SFGS requires sampling several consecutive years, with the precision of these vital rates generally increasing with the length of the study. Although 3 years of CMR data is sufficient to obtain an estimate of the annual survival rate, variation in growth and survival rates among years (Rose and others, 2022) demonstrates that longer periods are necessary to capture fluctuations in response to changing environmental conditions. Another metric that can be used to evaluate individual health is body condition, or the mass of animals relative to their length (Peig and Green, 2009). Lower body condition in translocated animals compared to those in established populations can be an indicator of release costs or poor habitat quality at the recipient site (Platenberg and Griffiths, 1999). Because snake body condition can also decline because of environmental fluctuations such as drought (Rose and Todd, 2017), simultaneously monitoring SFGS at extant populations at CR and RR would be useful to understand if changes in body condition at LHC are unique to the reintroduced population or representative of broader regional trends. Although collecting data from multiple populations of SFGS requires substantial investment of time and money, the information gained about vital rates from reintroduced populations can be valuable enough to outweigh the costs (Runge, 2013).
As the reintroduction of SFGS proceeds, it would also be valuable to monitor the dispersal of SFGS beyond the initial reintroduction site. The movement of reintroduced snakes between nearby wetlands connected through suitable upland habitat could increase the probability of persistence for the entire reintroduced population (Brown and Kodric-Brown, 1977). Installing drift fences and funnel traps at wetlands near the reintroduction site could aid in capturing individuals dispersing to new, unoccupied habitats and following individuals moving between established subpopulations. Capture probability of dispersing snakes might be low if few individuals emigrate from the reintroduction site and density at new wetlands is low. Another possibility for monitoring dispersal to new habitats is the use of drift fences with camera traps (Hobbs and Brehme, 2017; Amber and others, 2021) or automated passive integrated transponder (PIT) tag readers (Walkup and others, 2023), instead of funnel traps. Unlike funnel traps, camera traps and automated PIT tag readers do not need to be checked daily, and the detection of SFGS by a passive sampling method could be used as a decision point to begin using CMR trapping at a pond. Drift fences with camera traps could also be used to encircle the wetland at which SFGS were released, and the dispersal of individual snakes away from the wetland could be tracked through photographs of snakes moving through openings in the drift fence array. Alternatively, radiotelemetry could be used to track the movements of reintroduced snakes. Radiotelemetry was used to track the movements and document mortality of giant gartersnakes (Thamnophis gigas) translocated to constructed wetlands in the Sacramento Valley of California (Nguyen and others, 2023). The smaller size of SFGS compared to giant gartersnakes could necessitate smaller transmitters with shorter battery life, and limit tracking to the largest adult female snakes only. Further, the complex topography and dense vegetation in parts of LHC could make locating SFGS using radiotelemetry challenging.
The focus of this report has been the demographic viability of reintroduced SFGS populations, but maintaining genetic diversity also is key to establishing a viable, self-sustaining population (Schwartz and others, 2007; Van Rossum and Hardy, 2022). Reintroducing SFGS from CR and RR, and potentially other source populations, to LHC would provide increased genetic diversity compared to either donor population alone, and could help alleviate concerns of inbreeding depression in the reintroduced population. In other taxa, the introduction of individuals from a genetically divergent population of the same species has been shown to increase genetic diversity and adaptive potential (Chan and others, 2019). Genotyping all SFGS that are released at LHC, and following up by genotyping SFGS that are born in the reintroduced population, could enable scientists and managers to track which individuals are contributing to future generations (Van Rossum and Hardy, 2022).
The data collected during the monitoring of SFGS demography, genetic diversity, and status of CRLF populations can all be integrated into an adaptive management approach to the reintroduction of SFGS. Advocates of an active adaptive management approach often argue for an experimental approach to reintroduction, in which different methods are attempted simultaneously, and their success is evaluated with empirical data (McCarthy and others, 2012). Regarding an endangered species, there is a heightened concern for the fate of translocated animals because the effects of removing animals based on the viability of the donor population are paramount. Still, adaptive management can be used to design a monitoring plan and to make decisions in response to data collected from the reintroduced population as the reintroduction proceeds. If data on the vital rates of individual SFGS and abundance of the reintroduced SFGS population indicate the population is declining or failing to grow fast enough to be viable over the next 20 years, it might be necessary to revisit the reintroduction strategy and make changes that will lead to greater population growth. For example, although population simulations indicated that the release of captive-reared juvenile SFGS resulted in the highest viability for the donor and reintroduced populations (Rose and others, 2023), monitoring data could indicate that survival of juvenile SFGS is low at the reintroduction site and, as a result, the population is not growing as expected. If juvenile survival in the wild was lower than expected, then this updated information could be incorporated into the demographic model and stakeholders could evaluate if releasing adult SFGS or mixed-age propagules were preferable alternatives. Monitoring data on the abundance and vital rates of SFGS could also indicate the population is growing faster than expected. In that case, the decision could be made to stop the addition of new snakes at the first reintroduction site and shift reintroductions to a new wetland, if the reintroduced population seems demographically and genetically viable.
Translocating animals to a location that is unoccupied requires the commitment of resources from many stakeholders over several years. Given the proximity of LHC to a known SFGS population approximately 4 km to the east at RR, it is worthwhile to consider if SFGS could naturally colonize LHC without human assistance. The intervening habitat between the eastern boundary of LHC and the western boundary of RR includes multiple wetlands, grassland and scrub that could act as a dispersal corridor. The primary obstacle to natural SFGS recolonization of LHC is barriers to dispersal along the eastern boundary of LHC. State Highway 84 runs north to south along a steep canyon near and along the eastern boundary of LHC. The steep slope and shaded redwood forest habitat in the canyon might present a natural deterrent to dispersal for SFGS, which prefer shrub cover and less densely forested woodlands. State Highway 84 could present a barrier to dispersal of SFGS, either behaviorally if snakes avoid crossing an open road where they are exposed to predators, or demographically if vehicles might kill the occasional SFGS attempting to disperse further west. Also, the town of La Honda east of LHC has a series of smaller roads and suburban development that is inhospitable to SFGS. The additional barrier presented by State Highway 84 makes natural colonization of LHC by SFGS less likely than if LHC and RR were connected by an unfragmented landscape.
The International Union for the Conservation of Nature (IUCN) recommends that any proposal for translocating wildlife should consider the potential benefits and costs (International Union for Conservation of Nature, 2013). A primary concern for reintroducing SFGS to LHC is the potential for negative effects on the donor SFGS populations at CR and RR. In simulated reintroductions, minimizing the number of adult snakes removed from the donor populations by releasing captive-reared juvenile snakes resulted in the lowest effect on the demographic viability of simulated donor populations (Rose and others, 2023). Beyond demographic viability, the genetic effects of translocating snakes from CR and RR to LHC must be considered. The CR and RR populations have low genetic effective population sizes (Ne; Wood and others, 2020), an indicator of reduced genetic diversity and possible risk of inbreeding depression. Therefore, the removal of snakes from CR and RR for translocation to LHC could cause further decreases in genetic diversity in these existing populations. Negative effects on donor SFGS populations at CR and RR could be mitigated by only transferring a fraction of captive-born neonates to LHC and returning the rest to the donor population. Also, in the future, reciprocal translocation of SFGS between populations within the same genetic cluster (for example, CR, RR, and Año Nuevo State Park) or even among the southern and northern genetic clusters could be necessary to increase genetic diversity and stave off negative effects of inbreeding depression (Heber and others, 2013; Forsdick and others, 2017; Grueber and others, 2017).
Another potential risk from the reintroduction of SFGS to LHC is that adding these predators to wetlands could have negative effects on native CRLF populations. The small founder population of SFGS is unlikely to have dramatic effects on CRLF populations in the first 3 to 5 years following translocation. Over time, if the SFGS population increases in abundance, SFGS could represent one of the main predators on CRLF at LHC, along with resident Santa Cruz gartersnakes. Given the long coevolutionary history of CRLF coexisting with gartersnakes, including SFGS, throughout California, it is unlikely that the presence of SFGS alone would drive a population of CRLF to extinction. San Francisco gartersnakes and CRLF coexist at RR and CR, as well as other extant SFGS populations. San Francisco gartersnakes feed on a variety of amphibians and fish (Larsen, 1994; U.S. Fish and Wildlife Service, 2020), and as generalists are unlikely to deplete CRLF populations. Also, large CRLF likely exceed the gape of SFGS, and the existence of a size-refuge could aid in CRLF persistence with gartersnake predators.
To quantify if the addition of SFGS at LHC has a negative effect on CRLF populations, egg mass counts can be used to track the abundance of this threatened frog species. Collecting a time series of egg mass counts at several wetlands before reintroduction would enable the use of a before-after control-impact (BACI) study design (Clinton and others, 2022). Continuing egg mass counts at the focal wetland where SFGS are reintroduced and at surrounding wetlands where no SFGS are present would be a valuable complement to monitoring SFGS abundance. Monitoring CRLF egg masses at sites where no SFGS are present is essential for determining whether any changes in the CRLF egg mass count at the focal wetland(s) where SFGS are reintroduced are the result of broader trends (for example, in response to interannual changes in precipitation) or occurred in response to predation by SFGS. Surveying for all native and non-native pond-breeding amphibians in the focal wetland(s) and wetlands without SFGS would further allow for quantification of community-level effects of SFGS reintroduction (Clinton and others, 2022). Based on the negative effects of American bullfrogs on CRLF documented elsewhere (Fisher and Shaffer, 1996; Lawler and others, 1999; Anderson and others, 2019), the spread of American bullfrogs into wetlands at LHC would represent a much greater threat to CRLF persistence than the reintroduction of SFGS.
In addition to CRLF, other potential amphibian prey species are widespread at LHC, including Sierran chorus frogs and Pacific newts. Sierran chorus frogs are generalists that breed in a variety of wetlands including seasonal and permanent ponds (Stebbins, 2003). Sierran chorus frogs may make up the bulk of SFGS diet during the late winter and spring (Kim and others, 2021), when adult frogs migrate to water bodies to breed and metamorphs (juvenile frogs that have recently undergone metamorphosis from the larval stage) emerge from the water. Sierran chorus frogs produce large cohorts of larvae and metamorphs that leave the water in large numbers in the late spring and early summer. Because gartersnakes are gape-limited predators (Shine, 1991), small Sierran chorus frogs might be particularly important prey for juvenile SFGS (Larsen, 1994; Kim and others, 2021). The high tolerance that SFGS have for the tetrodotoxin produced by Pacific newts suggests that newts are an important part of the diet for SFGS (Brodie and others, 2002). Pacific newts breed in the winter months in ponds, reservoirs, and stream pools. Pacific newt larvae take several months to develop into adult newts and generally emerge from the water in the late summer or early fall. Water bodies that support breeding by a variety of different amphibian species, which each can have different breeding periods and larval stages of varying duration, likely benefits SFGS by providing multiple sources of prey that are abundant at different times of year. The presence of Sierran chorus frogs and Pacific newts at several wetlands within LHC further bolsters the likelihood that the preserve has suitable prey for reintroduced SFGS.
If the reintroduction of SFGS to LHC is pursued in the future, several outstanding questions remain about the methods used to rear juvenile snakes in captivity and release snakes into the wild. For captive-reared snakes, it is unclear if “enrichment” by recreating more natural environments in captivity provides benefits once snakes are released into the wild (Roe and others, 2015). A potentially important decision is whether to use a “soft release” method, in which snakes are first released into enclosures at the reintroduction site, or a hard release in which snakes are released and free to disperse immediately. Evidence for the benefits of a soft release for translocated snakes are mixed (Sacerdote-Velat and others, 2014; Cornelis and others, 2021), and building enclosures that allow for foraging by semiaquatic gartersnakes would be difficult. Finally, translocating wild animals and rearing animals in captivity raise concerns about the risk of transmitting disease (Sainsbury and Vaughan-Higgins, 2012). Snake Fungal Disease (SFD) caused by the pathogen Ophidiomyces ophiodiicola is an emerging risk to wild snake populations (Lorch and others, 2016). Screening captured and captive-reared SFGS for SFD could be important to prevent spread of the fungal pathogen between populations. These choices and other decisions could benefit from convening a panel of experts in gartersnake ecology, animal translocation, and captive-rearing to develop plans for each step of the process, data to collect to evaluate efficacy of chosen methods, and decision points to reevaluate the chosen methods should initial efforts result in negative outcomes for reintroduced SFGS.
Conclusions
La Honda Creek Open Space Preserve (LHC) is a suitable candidate site for a trial reintroduction of San Francisco gartersnakes (SFGS) to unoccupied habitat within its historical range. The climate, habitat, prey community, and land use at LHC are all similar to nearby open space preserves that support extant populations of SFGS. Based on our analysis, the following conditions could increase the likelihood of successful population establishment: annual releases of SFGS for more than 10 years, initial releases at a single wetland (in other words, founding one subpopulation of SFGS), and annual monitoring of the reintroduced management approach, where decisions can be made quickly to change management actions based on the health, genetic diversity, and vital rates of reintroduced SFGS. Furthermore, excluding cattle from the vicinity of the focal wetland at which snakes are reintroduced would likely improve the viability of the population by providing foraging and refuge habitat. Before reintroduction begins, many decisions will need to be made about the number, source, and specific destination of translocated animals. Bringing together the many stakeholders concerned with wildlife conservation, open space preservation, and maintaining working landscapes could be vital to ensuring that the reintroduction of SFGS to LHC is successful.
References Cited
Amber, E.D., Lipps, G.J., Jr., and Peterman, W.E., 2021, Evaluation of the AHDriFT camera trap system to survey for small mammals and herpetofauna: Journal of Fish and Wildlife Management, v. 12, no. 1, p. 197–207. [Available at https://doi.org/10.3996/JFWM-20-016.]
Bellis, J., Bourke, D., Maschinski, J., Heineman, K., and Dalrymple, S., 2020, Climate suitability as a predictor of conservation translocation failure: Conservation Biology, v. 34, no. 6, p. 1473–1481. [Available at https://doi.org/10.1111/cobi.13518.]
Bennett, V.A., Doerr, V.A.J., Doerr, E.D., Manning, A.D., and Lindenmayer, D.B., 2012, The anatomy of a failed reintroduction—A case study with the Brown Treecreeper: Emu—Austral Ornithology, v. 112, no. 4, p. 298–312. [Available at https://doi.org/10.1071/MU11048.]
Bertolero, A., Pretus, J.L., and Oro, D., 2018, The importance of including survival release costs when assessing viability in reptile translocations: Biological Conservation, v. 217, p. 311–320. [Available at https://doi.org/10.1016/j.biocon.2017.11.023.]
Biebach, I., and Keller, L.F., 2012, Genetic variation depends more on admixture than number of founders in reintroduced Alpine ibex populations: Biological Conservation, v. 147, no. 1, p. 197–203. [Available at https://doi.org/10.1016/j.biocon.2011.12.034.]
Brodie, E.D., Jr., Ridenhour, B.J., and Brodie, E.D., III, 2002, The evolutionary response of predators to dangerous prey—Hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts: Evolution, v. 56, no. 10, p. 2067–2082. [Available at https://doi.org/10.1111/j.0014-3820.2002.tb00132.x.]
Brown, J.H., and Kodric-Brown, A., 1977, Turnover rates in insular biogeography—Effect of immigration on extinction: Ecology, v. 58, no. 2, p. 445–449. [Available at https://doi.org/10.2307/1935620.]
Burke, V.J., and Gibbons, J.W., 1995, Terrestrial buffer zones and wetland conservation—A case study of freshwater turtles in a Carolina Bay: Conservation Biology, v. 9, no. 6, p. 1365–1369. [Available at https://doi.org/10.1046/j.1523-1739.1995.09061365.x.]
Caughley, G., 1994, Directions in conservation biology: Journal of Animal Ecology, v. 63, no. 2, p. 215–244. [Available at https://doi.org/10.2307/5542.]
Chan, W.Y., Hoffmann, A.A., and van Oppen, M.J.H., 2019, Hybridization as a conservation management tool: Conservation Letters, v. 12, no. 5, 11 p. [Available at https://doi.org/10.1111/conl.12652.]
Clinton, S.M., Hartman, J., Macneale, K.H., and Roy, A.H., 2022, Stream macroinvertebrate reintroductions—A cautionary approach for restored urban streams: Freshwater Science, v. 41, no. 3, p. 507–520. [Available at https://doi.org/10.1086/721471.]
Cornelis, J., Parkin, T., and Bateman, P.W., 2021, Killing them softly—A review on snake translocation and an Australian case study: The Herpetological Journal, v. 31, no. 3, p. 118–131. [Available at https://doi.org/10.33256/31.3.118131.]
Diffenbaugh, N.S., Swain, D.L., and Touma, D., 2015, Anthropogenic warming has increased drought risk in California: Proceedings of the National Academy of Sciences of the United States of America, v. 112, no. 13, p. 3931–3936. [Available at https://doi.org/10.1073/pnas.1422385112.]
Doak, D.F., Morris, W.F., Pfister, C., Kendall, B.E., and Bruna, E.M., 2005, Correctly estimating how environmental stochasticity influences fitness and population growth: American Naturalist, v. 166, no. 1, p. E14–E21. [Available at https://doi.org/10.1086/430642.]
Ewen, J.G., and Armstrong, D.P., 2007, Strategic monitoring of reintroductions in ecological restoration programmes: Ecoscience, v. 14, no. 4, p. 401–409. [Available at https://doi.org/10.2980/1195-6860(2007)14[401:SMORIE]2.0.CO;2.]
Fisher, R.N., and Shaffer, H.B., 1996, The decline of amphibians in California’s Great Central Valley: Conservation Biology, v. 10, no. 5, p. 1387–1397. [Available at https://doi.org/10.1046/j.1523-1739.1996.10051387.x.]
Forsdick, N.J., Cubrinovska, I., Massaro, M., and Hale, M.L., 2017, Genetic diversity and population differentiation within and between island populations of two sympatric Petroica robins, the Chatham Island black robin and tomtit: Conservation Genetics, v. 18, p. 275–285. [Available at https://doi.org/10.1007/s10592-016-0899-1.]
Golden Gate National Parks Conservancy, Tukman Geospatial LLC, Aerial Information Systems, National Park Service, Midpeninsula Regional Open Space District, County of San Mateo, San Francisco Public Utilities Commission, Peninsula Open Space Trust, and San Mateo City/County Association of Governments, 2022, San Mateo County fine scale vegetation map (FileGDB): ArcGIS digital data, accessed August 18, 2023, at https://vegmap.press/San_Mateo_vegmap.
Global Biodiversity Information Facility, 2023a, GBIF occurrence download Lithobates catesbeianus: Global Biodiversity Information Facility web page, accessed October 27, 2023, at https://doi.org/10.15468/dl.v5ebnd.
Global Biodiversity Information Facility, 2023b, GBIF occurrence download Pseudacris regilla: Global Biodiversity Information Facility web page, accessed October 27, 2023, at https://doi.org/10.15468/dl.6p88y3.
Global Biodiversity Information Facility, 2023c, GBIF occurrence download Pseudacris sierra: Global Biodiversity Information Facility web page, accessed October 27, 2023, at https://doi.org/10.15468/dl.xwsssh.
Global Biodiversity Information Facility, 2023d, GBIF occurrence download Taricha: Global Biodiversity Information Facility web page, accessed October 27, 2023, at https://doi.org/10.15468/dl.4skm24.
Germano, J.M., and Bishop, P.J., 2009, Suitability of amphibians and reptiles for translocation: Conservation Biology, v. 23, no. 1, p. 7–15. [Available at https://doi.org/10.1111/j.1523-1739.2008.01123.x.]
Gershunov, A., Shulgina, T., Clemesha, R.E.S., Guirguis, K., Pierce, D.W., Dettinger, M.D., Lavers, D.A., Cayan, D.R., Polade, S.D., Kalansky, J., and Ralph, F.M., 2019, Precipitation regime change in western North America—The role of atmospheric rivers: Scientific Reports, v. 9, article no. 9944, 11 p. [Available at https://doi.org/10.1038/s41598-019-46169-w.]
Grueber, C.E., Sutton, J.T., Heber, S., Briskie, J.V., Jamieson, I.G., and Robertson, B.C., 2017, Reciprocal translocation of small numbers of inbred individuals rescues immunogenetic diversity: Molecular Ecology, v. 26, no. 10, p. 2660–2673. [Available at https://doi.org/10.1111/mec.14063.]
Halstead, B.J., Kleeman, P.M., Rose, J.P., and Fellers, G.M., 2023, Sierra Nevada amphibians demonstrate stable occupancy despite precipitation volatility in the early 21st century: Frontiers in Ecology and Evolution, v. 10, 19 p. [Available at https://doi.org/10.3389/fevo.2022.1040114.]
Halstead, B.J., Thompson, M.E., Amarello, M., Smith, J.J., Wylie, G.D., Routman, E.J., and Casazza, M.L., 2019, Effects of prescribed fire on San Francisco gartersnake survival and movement: The Journal of Wildlife Management, v. 83, no. 1, p. 231–240. [Available at https://doi.org/10.1002/jwmg.21585.]
Halstead, B.J., Wylie, G.D., Amarello, M., Smith, J.J., Thompson, M.E., Routman, E.J., and Casazza, M.L., 2011, Demography of the San Francisco gartersnake in Coastal San Mateo County, California: Journal of Fish and Wildlife Management, v. 2, no. 1, p. 41–48. [Available at https://doi.org/10.3996/012011-JFWM-009.]
Heber, S., Varsani, A., Kuhn, S., Girg, A., Kempenaers, B., and Briskie, J., 2013, The genetic rescue of two bottlenecked south island robin populations using translocations of inbred donors: Proceedings of the Royal Society B, Biological Sciences, v. 280, no. 1752, 8 p. [Available at https://doi.org/10.1098/rspb.2012.2228.]
Hobbs, M.T., and Brehme, C.S., 2017, An improved camera trap for amphibians, reptiles, small mammals, and large invertebrates: PLoS One, v. 12, no. 10, 15 p. [Available at https://doi.org/10.1371/journal.pone.0185026.]
International Union for Conservation of Nature, 2013, Guidelines for reintroductions and other conservation translocations (ver. 1): Gland, Switzerland, International Union for Conservation of Nature Species Survival Commission, 57 p. [Available at https://portals.iucn.org/library/efiles/documents/2013-009.pdf.]
Jachowski, D.S., Gitzen, R.A., Grenier, M.B., Holmes, B., and Millspaugh, J.J., 2011, The importance of thinking big—Large-scale prey conservation drives black-footed ferret reintroduction success: Biological Conservation, v. 144, no. 5, p. 1560–1566. [Available at https://doi.org/10.1016/j.biocon.2011.01.025.]
Jensen, J.B., Bailey, M.A., Blankenship, E.L., and Camp, C.D., 2003, The relationship between breeding by the gopher frog, Rana capito (Amphibia: Ranidae) and rainfall: The American Midland Naturalist, v. 150, no. 1, p. 185–190. [Available at https://doi.org/10.1674/0003-0031(2003)150[0185:TRBBBT]2.0.CO;2.]
Kim, R., Halstead, B.J., Routman, E.J., and Andersen, J., 2021, When introduced prey violates trophic hierarchy—Conservation of an endangered predator: Biological Conservation, v. 256, 12 p. [Available at https://doi.org/10.1016/j.biocon.2021.109019.]
Kim, R., Halstead, B.J., Wylie, G.D., and Casazza, M.L., 2018, Distribution and demography of San Francisco gartersnakes (Thamnophis sirtalis tetrataenia) at Mindego Ranch, Russian Ridge Open Space Preserve, San Mateo County, California: U.S. Geological Survey Open-File Report 2018–1063, 80 p., accessed September 18, 2019, at https://doi.org/10.3133/ofr20181063.
Lawler, S.P., Dritz, D., Strange, T., and Holyoak, M., 1999, Effects of introduced mosquitofish and bullfrogs on the threatened California red-legged frog: Conservation Biology, v. 13, no. 3, p. 613–622. [Available at https://doi.org/10.1046/j.1523-1739.1999.98075.x.]
Lorch, J.M., Knowles, S., Lankton, J.S., Michell, K., Edwards, J.L., Kapfer, J.M., Staffen, R.A., Wild, E.R., Schmidt, K.Z., Ballmann, A.E., Blodgett, D., Farrell, T.M., Glorioso, B.M., Last, L.A., Price, S.J., Schuler, K.L., Smith, C.E., Wellehan, J.F.X., Jr., and Blehert, D.S., 2016, Snake fungal disease—An emerging threat to wild snakes: Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, v. 371, no. 1709, 8 p. [Available at https://doi.org/10.1098/rstb.2015.0457.]
McCarthy, M.A., Andelman, S.J., and Possingham, H.P., 2003, Reliability of relative predictions in population viability analysis: Conservation Biology, v. 17, no. 4, p. 982–989. [Available at https://doi.org/10.1046/j.1523-1739.2003.01570.x.]
McCarthy, M.A., Armstrong, D.P., and Runge, M.C., 2012, Adaptive management of reintroduction, chap. 8 of Ewen, J.G., Armstrong, D.P., Parker, K.A., and Seddon, P.J., eds., Reintroduction biology—Integrating science and management: Oxford, Wiley-Blackwell, p. 256–289. [Available at https://doi.org/10.1002/9781444355833.ch8.]
Melbourne, B.A., and Hastings, A., 2008, Extinction risk depends strongly on factors contributing to stochasticity: Nature, v. 454, p. 100–103. [Available at https://doi.org/10.1038/nature06922.]
Moss, W.E., McDevitt-Galles, T., Muths, E., Bobzien, S., Purificato, J., and Johnson, P.T.J., 2021, Resilience of native amphibian communities following catastrophic drought—Evidence from a decade of regional-scale monitoring: Biological Conservation, v. 263, 8 p. [Available at https://doi.org/10.1016/j.biocon.2021.109352.]
Nguyen, A.M., Todd, B.D., and Halstead, B.J., 2023, Survival and establishment of captive‐reared and translocated giant gartersnakes after release: The Journal of Wildlife Management, v. 87, no. 3, 17 p. [Available at https://doi.org/10.1002/jwmg.22374.]
Nichols, J.D., and Armstrong, D.P., 2012, Monitoring for reintroductions, chap. 7 of Ewen, J.G., Armstrong, D.P., Parker, K.A., and Seddon, P.J., eds., Reintroduction biology—Integrating science and management: Oxford, Wiley-Blackwell, p. 223–255. [Available at https://doi.org/10.1002/9781444355833.ch7.]
Onley, I.R., White, L.C., Moseby, K.E., Copley, P., and Cowen, S., 2023, Disproportionate admixture improves reintroduction outcomes despite the use of low-diversity source populations—Population viability analysis for a translocation of the greater stick-nest rat: Animal Conservation, v. 26, no. 2, p. 216–227. [Available at https://doi.org/10.1111/acv.12812.]
Peig, J., and Green, A.J., 2009, New perspectives for estimating body condition from mass/length data—The scaled mass index as an alternative method: Oikos, v. 118, no. 12, p. 1883–1891. [Available at https://doi.org/10.1111/j.1600-0706.2009.17643.x.]
Platenberg, R.J., and Griffiths, R.A., 1999, Translocation of slow-worms (Anguis fragilis) as a mitigation strategy—A case study from south-east England: Biological Conservation, v. 90, no. 2, p. 125–132. [Available at https://doi.org/10.1016/S0006-3207(99)00023-3.]
PRISM Climate Group, 2022, PRISM 30-year normal precipitation—Annual: Corvallis, Oreg., PRISM Climate Group dataset, accessed October 18, 2023, at https://prism.oregonstate.edu.
Reeder, N.M.M., Byrnes, R.M., Stoelting, R.E., and Swaim, K.E., 2015, An endangered snake thrives in a highly urbanized environment: Endangered Species Research, v. 28, no. 1, p. 77–86. [Available at https://doi.org/10.3354/esr00677.]
Roe, J.H., Frank, M.R., Gibson, S.E., Attum, O., and Kingsbury, B., 2010, No place like home—An experimental comparison of reintroduction strategies using snakes: Journal of Applied Ecology, v. 47, no. 6, p. 1253–1261. [Available at https://doi.org/10.1111/j.1365-2664.2010.01886.x.]
Rose, J.P., Kim, R., Schoenig, E.J., Lien, P.C., and Halstead, B.J., 2022, Integrating growth and survival models for flexible estimation of size-dependent survival in a cryptic, endangered snake: Ecology and Evolution, v. 12, no. 4, 15 p. [Available at https://doi.org/10.1002/ece3.8799.]
Rose, J.P., Kim, R., Schoenig, E.J., Lien, P.C., and Halstead, B.J., 2023, Comparing reintroduction strategies for the endangered San Francisco gartersnake (Thamnophis sirtalis tetrataenia) using demographic models: PLoS One, v. 18, no. 10, 21 p. [Available at https://doi.org/10.1371/journal.pone.0292379.]
Rose, J.P., and Todd, B.D., 2017, Demographic effects of prolonged drought on a nascent introduction of a semi-aquatic snake: Biological Invasions, v. 19, no. 10, p. 2885–2898. [Available at https://doi.org/10.1007/s10530-017-1491-4.]
Runge, M.C., 2013, Active adaptive management for reintroduction of an animal population: The Journal of Wildlife Management, v. 77, no. 6, p. 1135–1144. [Available at https://doi.org/10.1002/jwmg.571.]
Sacerdote-Velat, A.B., Earnhardt, J.M., Mulkerin, D., Boehm, D., and Glowacki, G., 2014, Evaluation of headstarting and release techniques for population augmentation and reintroduction of the smooth green snake: Animal Conservation, v. 17, no. S1, p. 65–73. [Available at https://doi.org/10.1111/acv.12151.]
Sainsbury, A.W., and Vaughan-Higgins, R.J., 2012, Analyzing disease risks associated with translocations: Conservation Biology, v. 26, no. 3, p. 442–452. [Available at https://doi.org/10.1111/j.1523-1739.2012.01839.x.]
Schoenig, E.J., Lien, P.C., Rose, J.P., and Halstead, B.J., 2023a, San Francisco gartersnake fecundity data from San Mateo County, California collected from 2019–2022: U.S. Geological Survey data release, accessed August 30, 2023, at https://doi.org/10.5066/P9U5NGXY.
Schwartz, M.K., Luikart, G., and Waples, R.S., 2007, Genetic monitoring as a promising tool for conservation and management: Trends in Ecology & Evolution, v. 22, no. 1, p. 25–33. [Available at https://doi.org/10.1016/j.tree.2006.08.009.]
Semlitsch, R.D., and Bodie, J.R., 2003, Biological criteria for buffer zones around wetlands and riparian habitats for amphibians and reptiles: Conservation Biology, v. 17, no. 5, p. 1219–1228. [Available at https://doi.org/10.1046/j.1523-1739.2003.02177.x.]
Shi, J., Joshi, J., Tielbörger, K., Verhoeven, K.J.F., and Macel, M., 2018, Costs and benefits of admixture between foreign genotypes and local populations in the field: Ecology and Evolution, v. 8, no. 7, p. 3675–3684. [Available at https://doi.org/10.1002/ece3.3946.]
Shine, R., 1991, Why do larger snakes eat larger prey items?: Functional Ecology, v. 5, no. 4, p. 493–502. [Available at https://doi.org/10.2307/2389631.]
Swain, D.L., 2021, A shorter, sharper rainy season amplifies California wildfire risk: Geophysical Research Letters, v. 48, no. 5, 5 p. [Available at https://doi.org/10.1029/2021GL092843.]
Swain, D.L., Langenbrunner, B., Neelin, J.D., and Hall, A., 2018, Increasing precipitation volatility in twenty-first-century California: Nature Climate Change, v. 8, no. 5, p. 427–433. [Available at https://doi.org/10.1038/s41558-018-0140-y.]
Taylor, G., Canessa, S., Clarke, R.H., Ingwersen, D., Armstrong, D.P., Seddon, P.J., and Ewen, J.G., 2017, Is reintroduction biology an effective applied science?: Trends in Ecology & Evolution, v. 32, no. 11, p. 873–880. [Available at https://doi.org/10.1016/j.tree.2017.08.002.]
U.S. Fish and Wildlife Service, 1973, Endangered species act of 1973—As amended through the 108th Congress: U.S. Fish and Wildlife Service report, 41 p. [Available at https://www.fws.gov/law/endangered-species-act.]
U.S. Fish and Wildlife Service, 1985, Recovery plan for the San Francisco garter snake (Thamnophis sirtalis tetrataenia): Portland, Oreg., U.S. Fish and Wildlife Service, Endangered Species Program report, 77 p., accessed February 21, 2014, at https://www.fws.gov/node/68304.
U.S. Fish and Wildlife Service, 1996, Endangered and threatened wildlife and plants—Determination of threatened status for the California red-legged frog: Federal Register, v. 61, no. 101, p. 25813–25833. [Available at https://www.federalregister.gov/documents/1996/05/23/96-12901/endangered-and-threatened-wildlife-and-plants-determination-of-threatened-status-for-the -california.]
U.S. Fish and Wildlife Service, 2020, Species status assessment for the San Francisco gartersnake (Thamnophis sirtalis tetrataenia) version 1.0. May 2020: Sacramento, Calif., U.S. Fish and Wildlife Service report, 95 p. [Available at https://www.fws.gov/node/70313.]
Van Rossum, F., and Hardy, O.J., 2022, Guidelines for genetic monitoring of translocated plant populations: Conservation Biology, v. 36, no. 1, 13 p. [Available at https://doi.org/10.1111/cobi.13670.]
Walkup, D.K., Ryberg, W.A., Pierce, J.B., Smith, E., Childress, J., East, F., Pierce, B.L., Brown, P., Fielder, C.M., and Hibbitts, T.J., 2023, Testing the detection of large, secretive snakes using camera traps: Wildlife Society Bulletin, v. 47, no. 1, 16 p. [Available at https://doi.org/10.1002/wsb.1408.]
Wood, D.A., Rose, J.P., Halstead, B.J., Stoelting, R.E., Swaim, K.E., and Vandergast, A.G., 2020, Combining genetic and demographic monitoring better informs conservation of an endangered urban snake: PLoS One, v. 15, no. 5, 27 p. [Available at https://doi.org/10.1371/journal.pone.0231744.]
Abbreviations
CMR
capture-mark-recapture
CR
Cloverdale Ranch Open Space Preserve
CRLF
California red-legged frog
DR
Driscoll Ranch
GBIF
Global Biodiversity Information Facility
LHC
La Honda Creek Open Space Preserve
MROSD
Midpeninsula Regional Open Space District
OSP
Open Space Preserve
RR
Russian Ridge Open Space Preserve
SFGS
San Francisco gartersnake
SVL
snout-vent length
USGS
U.S. Geological Survey
For more information concerning the research in this report, contact the
Director, Western Ecological Research Center
U.S. Geological Survey
3020 State University Drive East
Sacramento, California 95819
https://www.usgs.gov/centers/werc
Publishing support provided by the Science Publishing Network,
Sacramento Publishing Service Center
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
Rose, J.P., Schoenig, E.J., Kim, R., Nguyen, A.M., and Halstead, B.J., 2025, Assessing the feasibility of reintroducing San Francisco gartersnakes (Thamnophis sirtalis tetrataenia) to La Honda Creek Open Space Preserve, San Mateo County, California: U.S. Geological Survey Open-File Report 2024–1073, 25 p., https://doi.org/10.3133/ofr20241073.
ISSN: 2331-1258 (online)
Study Area
| Publication type | Report |
|---|---|
| Publication Subtype | USGS Numbered Series |
| Title | Assessing the Feasibility of Reintroducing San Francisco Gartersnakes (Thamnophis sirtalis tetrataenia) to La Honda Creek Open Space Preserve, San Mateo County, California |
| Series title | Open-File Report |
| Series number | 2024-1073 |
| DOI | 10.3133/ofr20241073 |
| Publication Date | January 22, 2025 |
| Year Published | 2025 |
| Language | English |
| Publisher | U.S. Geological Survey |
| Publisher location | Reston, VA |
| Contributing office(s) | Western Ecological Research Center |
| Description | viii, 25 p. |
| Country | United States |
| State | California |
| County | San Mateo County |
| Other Geospatial | La Honda Creek Open Space Preserve |
| Online Only (Y/N) | Y |