

Science Needs for Determining the Effects of Climate Change on Harmful Algal Blooms in the Southeastern United States
Links
- Document: Report (7.45 MB pdf) , HTML , XML
- Download citation as: RIS | Dublin Core
Acknowledgments
Funding for this project was provided by the Southeast Climate Adaptation Science Center, SECASC Project 056.
The authors wish to thank Dr. Victor Roland, U.S. Geological Survey, for guidance on hydrology and water-quality models, and Dr. Jennifer Cartwright, U.S. Geological Survey, for guidance on climate models.
Abstract
The Southeastern United States has many lakes, streams, and reservoirs that serve as important drinking water sources with recreational, agricultural, and ecological uses. However, harmful algal blooms (HABs) are becoming more common in these waters, causing health issues for humans and animals. HABs have been listed as a contaminant of emerging concern, and the magnitude, frequency, and duration of HABs appear to be increasing at the global scale. While it is well known that nutrients stimulate algae growth, it is not clear how climate change and other parameters stimulate the development of toxin production by HABs. The scientific literature describes parameters, such as storm occurrence, temperature, dissolved metals, erosion of soils, increasing length of growing season, discharge, and hydroperiod, that may affect algae growth and toxin production. Climate and hydrologic models address many of the physical and environmental parameters that influence HABs, but no climate models directly address HABs. This report compiles information from the existing literature pertaining to HABs and the modeling and forecasting of HABS. This compilation is done through the incorporation of climate change models. HAB research that involves climate change will require multiple disciplines that bring together ecologists, hydrologists, climatologists, engineers, economists, and new technology. Resource managers could use geographic data about the occurrence and distribution of HABs to develop models that identify waterbodies more vulnerable to HAB events. Development of such models will require teams capable of integrating biological, chemical, and physical factors. Model development will require additional research that can resolve anthropogenic and climate-related environmental factors to identify trends in freshwater HABs. The complexity and interconnectedness of the parameters that influence HAB occurrences will make model development challenging and require rigorous regional calibration.
Introduction
The U.S. Geological Survey (USGS), in cooperation with the Southeast Climate Adaptation Science Center and Tennessee State University, College of Agriculture, has developed this review to organize research on what is needed to develop models that predict harmful algal blooms (HABs) in a changing world. This review could help water resource and landscape planners develop new management tools to better control and minimize HABs in the future. The Southeastern United States is a water-rich region, with many rivers that have been impounded. Large reservoir systems in the Southeastern United States have traditionally been managed for flood control, commerce, hydropower, recreation, and drinking water. Small reservoirs and streams on private property, such as livestock ponds, are also vulnerable to HABs, which may cause problems with farm agricultural health (Munn and others, 2018). Likewise, urban areas and new developments are using retention and detention ponds for stormwater runoff control from nonpoint sources. Along the southeastern coastline, studies have found that shallow ephemeral ponds are susceptible to HABs (figs. 1A, B; Lewitus and others, 2008; Grogan and others, 2023). Incorporating HAB control into future management plans for these large and small impoundments could help balance protection of ecosystem and human health while maintaining other functions. Even urban stream systems in the southeast have shown that they can produce benthic HABs and associated cyanotoxins (Loftin and others, 2016). Knowledge of historical patterns is a critical driver of risk perceptions and can motivate the development of management plans that reduce future exposure to HABs (Treuer and others, 2021). Climate change is contributing to historical patterns becoming a less reliable predictor of the future for managing episodic HABs.

Waterbodies being monitored for harmful algal blooms: A, a reservoir at J. Percy Priest Dam, Tennessee, and B, a shallow ephemeral pond. Photographs by T.D. Byl, U.S. Geological Survey.
The objective of this study was to synthesize available literature on physical, biological, and chemical conditions that stimulate HABs in the Southeastern United States and ways in which climate change will affect those conditions. This report provides background information about the ecology of HABs, followed by a description of the roles that climate change and physical, biological, and chemical parameters play in the development of HABs. The physical, biological, and chemical parameters that influence HABs are then discussed in the context of climate models to determine additional needs to improve predictions of the effects of climate change on HABs.
Background on HABs and Cyanobacteria
HABs are the result of rapid growth of cyanobacteria and (or) algae in waterbodies. Marine (saltwater) and freshwater HAB events are caused by different alga types. Marine events, commonly known as red tide events, are not discussed in this report. Some extensive reviews of marine HABs in the literature, in addition to those listed here, are from Hallegraeff and others (2003) as part of the United Nations Educational, Scientific and Cultural Organization (UNESCO) and GlobalHAB (2021) programs.
The Centers for Disease Control and Prevention (CDC) received 368 reports of HAB events from 16 States in 2021 (CDC, 2023); 90 percent of these events occurred in freshwater systems, primarily in lakes, reservoirs, and impounded rivers (note: not all States report HAB events to the CDC). Of the reported events, HABs caused 117 cases of human illness and 2,715 cases of animal illness or death. Freshwater HAB events usually occur with eutrophic algal blooms, but not all algal blooms produce harmful toxins. Eutrophication is an excessive amount of nutrients that causes dense growth of algae and bacteria, which can result in extreme diurnal fluctuations of dissolved oxygen concentrations and pH. HAB events may involve excessive algae growth but are defined by the production and release of algal toxins. Eutrophication and HAB events rarely kill adult, air-breathing organisms, but they frequently kill aquatic organisms primarily through lack of oxygen, causing suffocation. HAB events can result in the illness or death of aquatic and terrestrial organisms through the uptake of toxins directly or through the food chain. This can result in the illness or death of air-breathing organisms as the toxin moves through the food web (Breinlinger and others, 2021; CDC, 2023).
HABs have a well-established presence in the Southeastern United States. The water-rich, warm landscape creates favorable habitat to support HAB growth (Paerl and Huisman, 2008). Monitoring and tracking of HABs in the Southeastern United States has drastically improved over the past decade (Graham and others, 2016; CDC, 2023). The U.S. Environmental Protection Agency (EPA) has a national ArcGIS mapping program that follows HAB incidents around the United States, and maps are updated each month based on HAB reports from the 48 contiguous States (EPA, 2024). Studies have indicated an increase in the frequencies and amounts of HABs in the southeast, not only because of increased awareness and monitoring (Hallegraeff and others, 2020), but also because of warming temperatures, intense storms, flash droughts, and nutrient enrichment (Ho and others, 2019; Smucker and others, 2021).
Although climate change is a global issue, its effects will not be equal and proportional across the globe. The Southeastern United States has experienced an increase in temperatures and precipitation during the last 20 years (Carter and others, 2018), while other parts of the contiguous United States have experienced increased drought, heat waves, and extreme rains. In response to these changing environmental conditions, cyanobacteria that once played an important role in the balanced freshwater ecosystems are now growing out of control, leading to oxygen depletion and altered food webs (Paerl and Huisman, 2008). In 2019, the Intergovernmental Panel on Climate Change directly linked HABs to climate change, noting that since the 1980s, the expansion of HAB ranges and frequency is a response to both climatic and nonclimatic drivers (Gobler, 2020). HABs are climate change co-stressors that have complex interactions with aquatic species and watershed health (Paerl, Scott, and others, 2016; Griffith and Gobler, 2020; Wells and others, 2020).
As climate change progresses, HABs are expected to become a primary water-quality issue (Brooks and others, 2016), increasing the need for water managers to better utilize predictive models built on knowledge about the physical, chemical, and biological drivers of HABs to develop strategies that control HAB events. Adapting to climate change has been reported to involve modifying infrastructure designs and stabilizing waterways and ecosystems to withstand increasing heat waves, flash droughts, and floods (Boehlert and others, 2015; Paerl, Gardner, and others, 2016). Few previous HAB studies have involved global climate models (GCMs), such as general circulation models or other predictive models, perhaps because of the uncertainties associated with the biological responses. Predictive models that primarily build on physical and chemical parameters and use biological generalizations could be used to evaluate future scenarios and identify strategies that enable adaptation to climate-change-induced HAB events.
Drivers of HAB Events
A recent surge in HAB publications has led to some discrepancies about perceived factors that bring about HABs. One central theme of all these publications is the goal of understanding the drivers of HABs in freshwater systems with a focus on climate change and nutrients (Journey and others, 2013; Paerl, Scott, and others, 2016; Ho and Michalak, 2020; Hellweger and others, 2022). Questions addressed in the literature include the following:
-
• What physiological or ecological traits enable toxic algal blooms to flourish where they were once rare?
-
• How might climate change influence and alter these interactions of physiological and ecological traits in ways that might favor, or suppress, toxic planktonic and benthic HAB species?
-
• What knowledge is needed to develop a predictive model that can incorporate climate-change stressors and cyanobacterial responses?
Much of this information may already have been published in the scientific literature or in data archives that have not yet been compiled and transformed into mechanistic or statistical models. A brief review of the HAB literature confirms a wide range of investigative methods were used in the studies. These differences in methods impede attempts to synthesize themes across studies.
For this study, drivers of HABs, especially those related to climate change, were organized into three main categories: physical, biological, and chemical (fig. 2). Physical and chemical parameters that drive HABs tend to be more easily defined or characterized with monitoring. The complexity of biological species, such as their physiology, genetic variations, growth requirements, life cycles, tolerances, habitats, and trophic interactions, complicate efforts to summarize in a report; therefore, some generalizations have been made. Because of the biological complexity and uncertainty, some amount of oversimplification of HAB responses to an itemized list of drivers was necessary. This is a limitation of this study, and this topic should be revisited as new information becomes available.
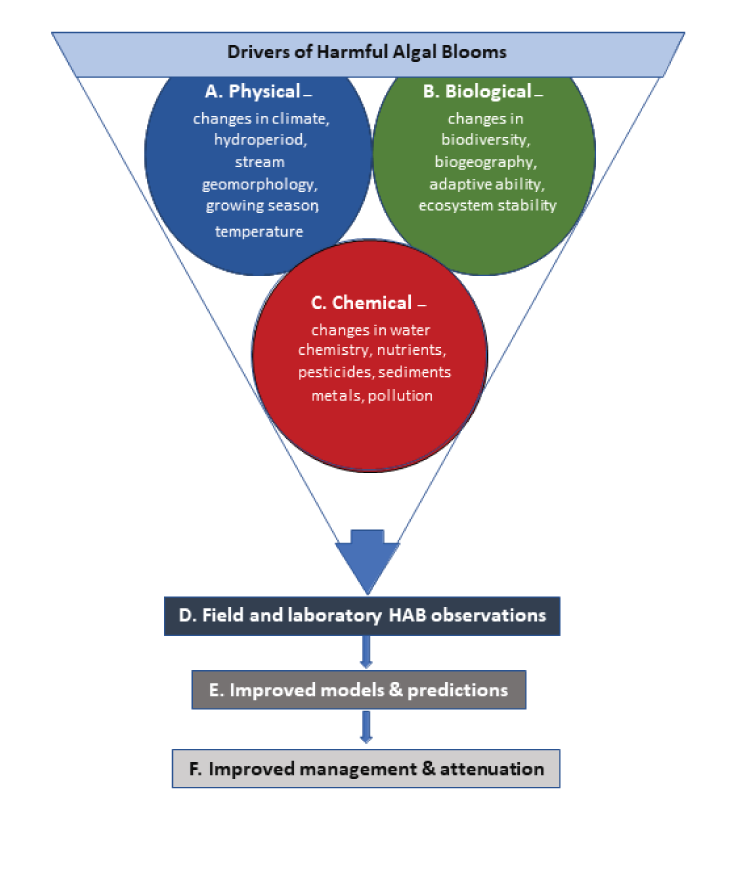
The physical, biological, and chemical drivers of harmful algal blooms (HABs) that could be used to improve models used by resource managers to reduce the negative effects of the HABs.
Biological Drivers of HABs
The cyanobacteria and algae responsible for HAB events release secondary metabolic compounds (Graham and others, 2012; Manganelli and others, 2012; Huang and Zimba, 2019). As secondary metabolites, their true ecological function is unknown. These secondary metabolites are not always present and are not required for the algae or cyanobacteria to complete their lifecycle. The secondary metabolites range in toxicity from nonthreatening bad smells and tastes to skin rashes or even tissue and organ failure (table 1). A brief discussion of toxic secondary metabolites, or algal toxins, is presented in this report to provide context and an introduction to the scope of the hazards. More rigorous toxicology discussions can be found in the literature (Manganelli and others, 2012; Dittmann and others, 2013; Huang and Zimba, 2019; Chorus and Welker, 2021).
Table 1.
Secondary metabolites, or algal toxins, produced by various freshwater cyanobacteria and their primary forms of toxicity in humans and wildlife (Mur and others, 1999; Carmichael and Boyer, 2016; Metcalf and others, 2021).The primary microalgae responsible for producing freshwater toxins are cyanobacteria, commonly referred to as blue-green algae because they have a greater abundance of phycocyanin pigment than other algae (Wheeling, 2019). Cyanobacteria biology is discussed herein to provide context for these ancient and complex organisms.
Cyanobacteria lineage can be traced back to the earliest life forms on Earth, and they helped to form the Earth’s oxygen-rich atmosphere (Taylor and Taylor, 1993; Popall and others, 2020). Cyanobacteria survived and evolved over 2 billion years, maintaining competitive ecological niches. Their evolutionary adaptations gave them an ecological advantage as environmental conditions varied in response to geological climate change. Cyanobacteria are able to adapt to use different light wavelengths. Some are capable of using far-red light (Nürnberg and others, 2018), and some have even adapted to extremely low light in show caves (Mur and others, 1999; Morton and others, 2020). These traits, among others, grant flexibility and growth advantage in slow moving waters. The capacity to develop gas vesicles has enhanced their ability to change their buoyant density and move up and down in the water column to optimize nutrient uptake, gas exchange, and photosynthesis. If growing conditions deteriorate, cyanobacteria can settle to the bottom of the water column and remain dormant for extended periods of time, reemerging when optimal growing conditions return (Preece and others, 2017).
Cyanobacteria function like algae in aquatic ecosystems; therefore, they are typically considered to be part of algal communities (Graham and others, 2016). Cyanobacteria are actually photosynthesizing prokaryotes related to bacteria that differ from eukaryotic algae like Chlorophyta, which are more closely related to plants (fig. 3) (Metcalf and others, 2021). Cyanobacteria grow in a variety of aquatic environments and even in some terrestrial environments. They can range in size from microscopic single cells to large colonies or chains of cells that work together. There are many different species of cyanobacteria, and not all cyanobacteria are capable of producing toxins (Graham and others, 2016). Cyanobacteria have different strategies for living in the water: cyanobacteria floating in the water column are referred to as planktonic; those attached to rocks, sticks, and plants in the water are periphytic; and those growing on the bottom of the waterbody are benthic. Some cyanobacteria can live in all three types of habitats. Production of toxins is not affected by how they live (planktonic, periphytic, or benthic) but is affected by the presence of the toxin-encoding genes.

Eutrophic waters with a mix of filamentous Hydrodictyon (Chlorophyta) algae floating on top and intertwined with the macrophytes, various cyanobacteria, and rooted aquatic macrophytes (Stuckenia pectinata and Ceratophyllum demersum) in a shallow body of water. Photograph by T.D. Byl, U.S. Geological Survey.
Many cyanobacteria grow faster in warmer water temperatures as compared to Chlorophyta (green algae) and diatoms (Paerl and Huisman, 2008; Visser and others, 2016a; Gobler and others, 2017; Tanvir and others, 2021). Planktonic cyanobacteria have an optimum temperature range of 27–37 degrees Celsius (°C), compared to other true algae such as Chlorophyta, which have an optimum range of 27–32 °C, or diatoms, which have an optimum range of 17–27 °C (Paerl and Huisman, 2009; Paerl, 2014). Cyanobacteria continue to function at high efficiency when temperatures exceed the growing threshold of their closest competitors. Because water temperatures in shallow lakes and streams have begun to routinely exceed 32 °C (Carter and others, 2018), an increase in cyanobacteria as part of freshwater algal communities is expected.
Nitrogen (N) and phosphorus (P) are considered the two most important nutrients for the growth of algae, including cyanobacteria, and certain cyanobacteria are capable of fixing nonreactive atmospheric N (N2) into reactive N compounds. (Newell and others, 2019). A Food and Agriculture Report by the United Nations in the 1970s promoted the idea of using certain cyanobacteria in rice fields to provide a natural N fertilizer for the rice (Hesse, 1975). They did not know at the time that some of those N-fixing cyanobacteria could produce toxins that would complicate the safety of the rice production. Dolichospermum (formerly Anabaena), Nostoc, Aphanizomenon, and Cylindrospermopsis are a few genera of the toxin-producing cyanobacteria that can fix N2 gas. This N-fixing capability by many of the cyanobacteria complicates the nutrient story. For example, reducing N2 from surface-water runoff to reduce occurrence of HABs may not have the desired effect if toxin-producing cyanobacteria are present and can obtain reactive N from atmospheric gas through N-fixation. N-fixation capability elevates the role of P in the growth of cyanobacteria and the production of toxins because P becomes the limiting element; however, not all cyanobacteria can fix N. Microcystis, which can produce the microcystin toxin, is incapable of fixing N. Several studies found that N-to-P ratios play an important role in the production of microcystin toxin for that specific cyanobacterium (Beversdorf and others, 2013; Beaver and others, 2018). Despite the N-fixing capability of some cyanobacteria, reducing N runoff from farm fields and wastewater treatment is a way to reduce eutrophication and potential HAB development (Gobler and others, 2016). This rationale is supported by recent studies that have shown cyanobacterial N-fixation does not meet phytoplankton or ecosystem N demands (Paerl and Otten, 2013). The N-fixation process requires high energy, and oxygen production during photosynthesis can inhibit the anaerobic process; turbulent mixing caused by wind can also disrupt N-fixation. Additionally, cofactors such as iron, molybdenum, and other trace metals may limit the enzymes involved in N-fixation. Recent research continues to emphasize the role of P as the main nutrient that controls the development of HABs (Glibert and Burkholder, 2006; Davis and others, 2010).
In addition to N-fixation, cyanobacteria are also efficient at fixing carbon. As carbon dioxide (CO2) concentrations increase in the atmosphere, the dissolved CO2 in water is expected to increase, partly due to gas dissolution behavior described by Henry’s gas law, which predicts that if partial pressure of CO2 in the atmosphere increases, a proportionally higher amount of CO2 will dissolve into the water column. Other factors such as increased respiration of aerobic organisms will also increase the CO2 dissolved in water. Warming water tends to increase the metabolic respiration rate of bacteria (Morita, 1974), potentially enriching the dissolved CO2 well beyond what Henry’s gas law predicts. Toxin-producing cyanobacteria, such as Microcystis and Raphidiopsis raciborskii, take advantage of this enrichment and take up CO2 from the water more efficiently than other algae by using a CO2-concentrating mechanism before the RuBisCo carbon fixation step of photosynthesis. This means that not only do the cyanobacteria prefer the warmer waters with higher CO2, but they also can assimilate the carbon and divide faster than other phytoplankton, allowing them to become dominant in the HABs (Paerl and Huisman, 2009; Paerl and Paul, 2012; Visser and others, 2016b; Ji and others, 2020).
As mentioned previously, cyanobacteria can produce a variety of secondary metabolite compounds that include toxins such as dermal toxins, hepatotoxins, cytotoxins, and neurotoxins (table 1) (Mur and others, 1999; Carmichael and Boyer, 2016; Metcalf and others, 2021). Even within a particular cyanotoxin type, such as microcystin, there are more than 150 congeners because of substitutions on positions 2 and 4 of the basic molecule (Citriglia and others, 2019). Research on cyanobacteria continues to lead to the discovery of additional toxins such as the nonprotein amino acid β-N-methylamino-L-alanine, produced by cyanobacteria with neurotoxic properties (Metcalf and others, 2021). It is unknown how common these new toxins are in the environment because routine, inexpensive, standard analytical methods have not been established for these rare or recently discovered toxins. Additional toxins are likely to be discovered as scientists continue to study the chemistry and genetics of cyanobacteria. In fact, this is an active area of medicinal research due to several secondary metabolites having therapeutic properties; therefore, the medical field is searching for additional compounds with beneficial value (Pradhan and others, 2021).
Cyanobacterial secondary metabolites are receiving increased attention with regards to their chemistry, biosynthesis, ecological role, toxicity to different organisms, and potential biomedical use (Briand and others, 2003; Graham and others, 2008, 2016; Manganelli and others, 2012). While the national attention to these compounds started quite recently (about 2000) from a scientific standpoint, their biosynthesis evolved over 1 billion years ago. Aquatic ecology textbooks from the 1970–80s suggested that toxin synthesis developed primarily as a chemical defense mechanism against organisms that eat phytoplankton (DeMott and others, 1991; Henao and others, 2020). However, new genetic research suggests that cyanotoxin production likely predated consumers of phytoplankton. For example, evolutionary gene analysis and phylogenetic investigation of primary sequences indicate that the gene cluster responsible for saxitoxin production in early Nostocales, an order of common cyanobacteria, has been present for at least 2.1 billion years, but planktivorous organisms emerged only 1.5 billion years ago (Murray and others, 2011). Phylogenetic evidence for the pre-phytoplanktivore evolution of microcystin synthesis has also been identified (Rantala and others, 2004), and indirect cues for similar phenomena in the case of cylindrospermopsin production also exist (Rzymski and Poniedziatek, 2014). These timescales imply that cyanobacterial toxins initially played ecological roles other than grazer defense and that their toxicity towards animals may only be an indirect effect of their production and release into the water column. The ecological function of the cyanobacteria’s secondary metabolites may have changed over geological time in a process known as exaptation, which would mean that the present-day ecological function of the toxin molecule could be to provide some protection to the cyanobacteria community from consumers. However, studies are ongoing to determine the ecological role these chemicals play. Knowing their ecological roles and what elicits the production of toxins could provide insight into management strategies to prevent the production of toxins. The literature provides a long list of possible functions, such as nutrient storage, chemical signals to other microorganisms, osmotic regulators, allelopathic chemicals, oxidative stress response, predator deterrent, geochemical facilitator for metals or oxygen uptake, mutualistic interaction, and antioxidant protection against ultraviolet radiation (Kaebernick and Neilan, 2001; Errera and Campbell, 2011; Paerl and Otten, 2013; Harris and others, 2014; Limardo and Worden, 2015; Fuentes and others, 2016; Wilhelm and others, 2020; Hellweger and others, 2022). Studies that combine regulation of toxin gene expression and environmental effectors could provide significant insight into determining the ecological function of these secondary metabolites (Neilan and others, 2013).
Physical and Chemical Parameters
Physical and chemical parameters related to climate change are expected to have an outsized response by cyanobacteria and HAB development. The main factor that will influence HAB occurrence is the relations among nutrients, eutrophication, and HAB development (Chorus and Zessner, 2021). Discussions among climatologists related to climate change and weather often include discussions of water resources and the volatility of rainstorms, short- and long-term droughts, and intense flooding. Unfortunately, that volatility is expected to bring more nutrients into surface waters due to more extreme rain events that result in intense storm runoff (Gudmundsson and others, 2021; Carpenter and others, 2022; Ryberg and Chanat, 2022). Several government agencies have shared results from general circulation climate models for the United States that predict more extreme rain patterns with the rising temperatures in the Southeastern United States (National Oceanic and Atmospheric Administration, 2023; National Park Service, 2023). The rationale for the extreme rains is that for every degree Celsius increase in temperature, the atmosphere can hold 7 percent more moisture (Santer and others, 2007). So, a region experiencing a heat wave with temperatures 3–6 °C above average can expect 21 to 42 percent more moisture in the atmosphere that will precipitate when the heat wave encounters cool air.
Extreme heat can cause increased evapotranspiration and flash drought conditions, such as vegetation dieback (Mohammadi and others, 2022). These conditions can also lead to more forest fires (Gao and others, 2021). Extremely dry soils with reduced vegetation or fire disturbance are less efficient at absorbing or percolating rainwater as compared to moist soils (Vogelmann and others, 2017). The end result is that heat waves can instigate flash drought conditions, making the topsoil more vulnerable to erosion, especially when the drought is punctuated by intense storms that drop large amounts of water in a short period of time. This weather pattern can lead to more nutrient and sediment runoff going into wetlands, rivers, and lakes (fig. 4) (Sinha and others, 2017; Zhai and others, 2020).

Conceptual model showing how heat waves can influence algal growth through enhanced erosion.
Nutrients in surface waters are the primary drivers of eutrophication, which is associated with HAB events (Glibert and Burkholder, 2006; Paerl and others, 2019; Ho and Michalak, 2020). Nutrients that promote eutrophication in a standing waterbody can originate from external nutrient loading in storm runoff or discharge or from historical internal loads that accumulate in the sediments. Nutrient reduction is considered the top strategy to prevent HABs (Interstate Technology and Regulatory Council [ITRC], 2022). While nutrient loading and increased eutrophication are directly connected, the development of HABs is more complicated than N and P concentrations because other physical processes that are associated with climate change influence the growth of toxin-producing cyanobacteria. For example, as temperatures rise, we can expect (1) increased frequency of intense storms; (2) increased residence times of the waters in the watershed prompted by greater time intervals between storms and altered hydroperiods; (3) decreased dissolved oxygen; (4) longer growing seasons and milder winters that lead to earlier cyanobacteria growth; (5) increased water demands from growing human populations and ecosystem requirements; and (6) climate-induced temperature stratification in the water column, which is also influenced by bathymetry and geomorphology of lakes and reservoirs (Carter and others, 2018; Terando and others, 2020; Woolway and others, 2021). These physical factors are tied to chemical factors, which ultimately influence the biology. For example, the increasing water temperatures will drive greater dynamic diurnal CO2 and oxygen fluctuations (USGS, 2018), accompanied by oscillating pH, thus provoking a shift in algal community composition.
Storm Intensity and Its Effect on Soil Erosion
Climatologists have documented an increase in the frequency and intensity of extreme daily precipitation since 1980 (Sun and others, 2021; Wood and others, 2021; Harp and Horton, 2022). An extreme precipitation event is defined as one in which the quantity of rain over a specified time exceeds a threshold or an average over some spatial region. These extreme rain events are often reported by using probability statistics, such as calling a storm a “100-year event,” which is shorthand for saying the chance that a rain event of that magnitude will occur is once in 100 years. Many regions of the country are now experiencing such events every 10–20 years. This change in frequency means the storm-probability models are having to be re-calculated based on more recent storms (Tabari, 2020). Lehmann and others (2015) reported that record-breaking rainfall events occurred 12 percent more often than expected globally from 1981 to 2010, with an estimated 26-percent chance that a record-setting rainfall event was due to anthropogenic climate change. The increased frequency of intense storms was projected by GCMs (Sillmann and others, 2013; Toreti and others, 2013). Not only has there been an increase in extreme storms, but there has also been a shift of when and where those storms occur (National Research Council, 2013). For example, the number of intense storms occurring during the dry season of summer and early fall has increased in the Southeastern United States (Walsh and others, 2014). Unfortunately, one consequence of these more intense storms is increased soil and nutrient erosion (Chapra and others, 2017; Gobler, 2020). Intense storms also cause a rapid rise in water levels and velocity in natural and engineered drainage networks, resulting in resuspending sediments rich in nutrients. The outcome of this external and internal nutrient loading is algal growth stimulation in the receiving rivers, wetlands, lakes, and reservoirs (Graham and others, 2020).
A review of soil erosion processes by Holz and others (2015) describes how antecedent soil moisture is the most important property influencing erosion during storms. The moisture content of the soil affects the structure and hydraulic infiltration rates during a storm (Cresswell and others, 1992; Ziadat and Taimeh, 2013). Extreme soil moisture, dry or wet, will influence the infiltration rate. For example, wetter soils will saturate quicker during a storm, creating overland flow and resulting in potential erosion; however, soils with less than 30 percent moisture often form a surface seal that reduces soil porosity on the surface. This seal hinders water infiltration while increasing overland flow and potential erosion. Note that soil moisture is not the only factor controlling infiltration rates; slope steepness, porosity, vegetation, and soil organisms also play a role in percolation and erosion. Climate change does not affect soil properties such as particle size, porosity, and soil texture, but it does exert a major influence on soil moisture, rain intensity, and the potential for erosion.
Soils and nutrients that erode during storms get transported into waterways and eventually end up in ponded waters, such as wetlands, lakes, and reservoirs. This external load of nutrients into the watershed can stimulate algae, cyanobacteria, and rooted aquatic macrophyte plants. Many of the large reservoirs in the Southeastern United States were built during 1930 to 1960; in the 1950s, it became apparent that these reservoirs not only controlled flow in large river networks, but they also trapped sediments (Brune, 1953). Brune (1953) calculated that United States reservoirs trapped between 30 and 90 percent of sediments that flowed into them, depending on the size and bathymetry of the reservoir and the frequency of rains. Over the many years of operation, many of the reservoirs have had to be dredged to keep from filling in. The sediments carried into the reservoirs also held a lot of nutrients and minerals. Yang and others (2017) compiled data from 157 studies and found that P accumulated faster than N in the sediments. Maavara and others (2020) used a mechanistic model to show that, on a global scale, reservoirs trap approximately 12 percent of the phosphorous being carried by major rivers, and that percentage is expected to rise to 17 percent by 2030 because of the construction of new dams. The internal trapped nutrients play a role in eutrophication and may delay recovery of waterbodies in watersheds that have been the focus of best management practices, improved wastewater treatments, or other landscape management strategies.
Changes in Hydroperiods and Flash Droughts
Climate change is expected to have a major influence on the hydroperiod, which is the number of days per year that an area is wet or inundated, further complicating the connections among trapped sediments, nutrients, and eutrophication. Evolving rain patterns caused by warming trends and increased moisture-holding capacity, coupled with fluctuating large-scale atmospheric circulation patterns, are expected to result in extreme temperatures and alter the frequency of rain events, which affects the natural hydroperiod (Terando and others, 2020).
Not only is climate change increasing the frequency and intensity of storms, but it is also increasing the frequency of flash droughts (Mohammadi and others, 2022). A flash drought is defined as a drought with a rapid onset and intensification caused by heat, lack of precipitation, and increased evapotranspiration (fig. 5). Flash droughts are considered a modification of the hydroperiod (Christian and others, 2024). As noted earlier, intense storms may provide as much or more water than average storms prior to 2000, but those rains fall in shorter, more intense bursts. Rather than facilitating optimum rain infiltration, intense storms enhance overland runoff. After intense storms subside, warm temperatures and periods without rain dry the soils through evapotranspiration, thus causing flash drought conditions.

Researchers investigating dried algae in a Tennessee wetland area during a flash drought in September 2022. Water levels are normally 2–3 feet deep where the researchers are working, but during this drought, the researchers were more than 100 feet away from any surface waters. Photograph by T.D. Byl, U.S. Geological Survey.
Not only do warm temperatures affect conditions on land, but they also stabilize thermal stratification in standing waters (fig. 6). Thermal stratification sets up two distinct water layers in lakes with sufficient depth where less dense, oxygen-rich water resides on top (epilimnion), and colder, denser, hypoxic water sinks to the bottom (hypolimnion). The thermal stratification and water density are so divergent that little mixing occurs between these two layers during the warm months. This stratification plays an important role in the biogeochemistry of internal nutrient loads, especially P (Wu and others, 2014; Rohwer and others, 2023). The term “internal nutrient load” refers to nutrients that have historically been transported into the still waters of a wetland, lake, or reservoir (lentic system) and have settled to the bottom and accumulated over years. The sediments are enriched with nutrients, soil, and organic matter, but lack oxygen. Anaerobic bacteria are active in these sediments, and during respiration they pass electrons to metals such as iron oxyhydroxides instead of oxygen. The metal oxides tend to bind P, ammonia, and other nutrients. The reduction of sedimentary metal oxides by the anaerobic bacteria, however, causes the metals to become soluble and release the bound phosphorus and other nutrients, stimulating algae growth (Orihel and others, 2016; Leung and others, 2021). This process increases the dissolved nutrients and metals in the water column and makes them accessible to cyanobacteria (fig. 7). Cyanobacteria can drift down to the hypolimnion to take in the nutrients they need and then rise back to the surface to continue doing photosynthesis (Peeters and others, 2007; Wilhelm and others, 2020). Anoxic conditions at the sediment–water interface in the hypolimnion have the potential to cause substantial nutrient release from sediments. In some situations, strong winds or cool night temperatures can cause localized upwelling of the nutrient-rich hypolimnion waters, causing a localized burst of algae growth (Visser and others, 2016a).
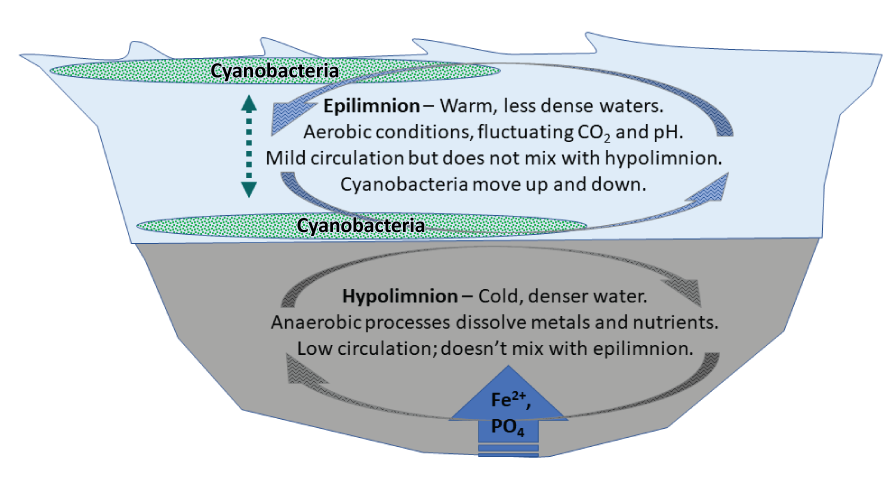
Lake and reservoir thermal stratification occurs during the warm season. The upper layer, called epilimnion, is warm and less dense than the lower layer, and cyanobacteria move up and down to optimize nutrient uptake and photosynthesis. The lower, colder, denser waters, called hypolimnion, have low dissolved oxygen concentrations, and the bacteria in the sediments help to release iron (Fe2+) and other metals through oxidation-reduction reactions, which also facilitate the release of absorbed phosphorus (PO4) and other nutrients. [CO2, carbon dioxide]
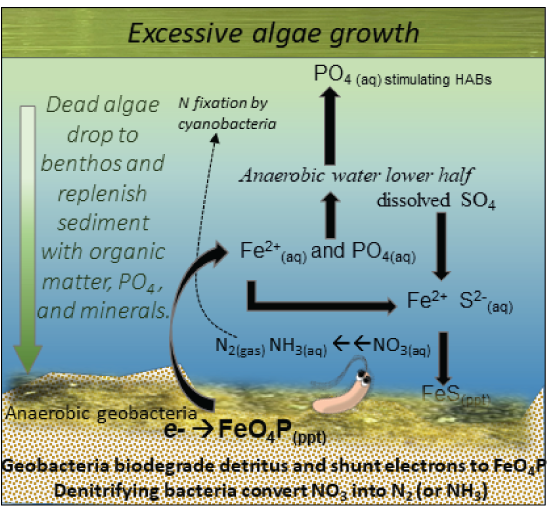
The biogeochemical cycles remobilize internal nutrient loads such as phosphorus and nitrogen (N) from depositional sediments, dissolving them back into the water column and making the nutrient loads accessible to growing cyanobacteria. [PO4, phosphorus ion; Fe2+(aq), aqueous ferrous iron; S2−(aq), aqueous sulfide; SO4, sulfate ion; FeS(ppt), iron sulfide precipitate; e−, electron transfer; FeO4P(ppt), ferric phosphorus precipitate; NO3, nitrate; N2, nitrogen; NH3, ammonia]
Longer stratification periods in lakes can cause decoupling between epilimnetic and hypolimnetic ecosystem processes and will influence the nutrient biogeochemistry (Summers and Ryder, 2024). Reservoir managers are reporting summer thermal stratification starting earlier in the spring and lasting longer into the fall. Woolway and others (2021) used the Inter-Sectoral Impact Model Intercomparison Project climate models in series with a suite of independently developed lake models to project that global warming will cause summer stratification to start 22 days earlier than 1970–99 conditions and last 11 days beyond 1970–99 conditions. Thermal stratification starting 22 days earlier will accelerate the creation of the hypolimnion, with subsequent effects on nutrient and phosphorus release from lake sediments, and will establish conditions that favor planktonic cyanobacteria over other planktonic algae.
Increasing Water Temperatures Lead to Decreased Dissolved Oxygen Levels
Climate change is expected to increase the temperature of freshwater systems which will then affect the solubility of dissolved oxygen (DO) in freshwater systems (Rajesh and Rehana, 2022). DO concentrations should decline with increasing water temperature because warmer water reduces gas solubility. Other mechanisms can also affect the concentration of DO, potentially amplifying or counteracting losses based on predicted solubility calculations. For example, the rate of heterotrophic microorganism respiration increases with temperature faster than primary production via photosynthesis. As noted earlier, surface-temperature warming can increase the strength and duration of thermal stratification, thus reducing water circulation and preventing deep-water DO replenishment. Rajesh and Rehana (2022) documented temperature trends in seven rivers over several decades, reporting a concurrent decrease in DO with an increase in temperatures. The rates at which the temperature increased and DO decreased varied in the seven catchments, probably due to oxygen release during photosynthesis and differences in water velocity. Jane and others (2021) used a large dataset from 393 freshwater lakes and found a general decline in DO that was primarily associated with reduced solubility under warmer water temperatures. The data also indicated that DO increased in a subset of highly productive warming lakes, probably because of an increase in the production of phytoplankton. This observation was supported by Secchi turbidity readings showing an increase in turbidity in the highly productive lakes. Oxygen trends in warming freshwater lakes were compared to warming ocean trends; declines in freshwater DO were 2.75 to 9.3 times greater than observed in the world’s oceans (Jane and others, 2021). The low oxygen levels in rivers and lakes can lead to a positive feedback mechanism, one where fish die due to lack of oxygen, which then causes an increase in biological oxygen demand and lower DO levels.
A potential trend of decreasing DO due to rising temperatures associated with climate change in waterbodies in the Southeastern United States is concerning because of the implications for small lakes and ponds in the region. A study of five small urban ponds in Kentucky found that DO quickly decreased with the onset of warm temperatures (de la Cruz and others, 2017). Pond size and depth seem to play a major role in warming and DO fluctuations. The small ponds also had HAB events occurring during warming spells in the winter, which is normally infrequent in large reservoirs. Common water-quality parameters, such as temperature and DO levels, associated with HABs in large lakes and rivers might need additional assessment when they are applied to smaller ponds or wetlands.
Longer Growing Seasons, Milder Winters
Climate is expected to extend the growing season considerably by delivering milder winters with earlier spring warming spells and later cooling in the fall. Because temperature is an important driver of HAB events, this is expected to influence the onset of these events and set up conditions favorable to cyanobacteria (Visser and others, 2016b). Toxin-producing cyanobacteria, such as Planktothrix, are common in the Southeastern United States, can persist in the water column of temperate lakes, and tend to bloom as temperatures rise in the early spring (fig. 8). Other toxin-producing cyanobacteria species tend to form akinetes, which are specialized cells that are resistant to cold, desiccation, and irradiation, or vegetative cells in lake sediments. Increasing temperatures in early spring may initiate earlier germination of akinetes (Tsujimura and Okubo, 2003).

Small shallow ponds are highly susceptible to early spring warming and resulting algal blooms, such as A, this small urban pond in Nashville, Tennessee, that B, experienced incredible algae growth after a 2-week warming period in early April 2023. Photographs by T.D. Byl, U.S. Geological Survey.
Bathymetry of Climate-Vulnerable Lakes and Reservoirs
The bathymetry of a reservoir includes the depth, slope of benthos, littoral zone conditions, land use activities, and shoreline-to-volume ratio. Shallow embayments and extensive shoreline-to-lake surface area ratios have a major influence on the growth of cyanobacteria and algal blooms; increased littoral zones, increased shoreline-to-volume ratios, and reduced flushing rates increased the occurrence of algae growth (Green and others, 2021; Kim and others, 2021). Kim and others (2021) reported that a combination of low-head dams and run-of-the-river impoundments in regulated rivers favored algal growth by slowing velocity, increasing water residence times, and providing improved light in shallow depths. Hydrologic alterations such as instream structures result in slower water velocity and increases in sediment deposition which can enrich the benthos with nutrients that are available for algal growth.
Six reservoir morphometric parameters—shoreline length, surface area, volume, maximum depth, flushing rate, and mean depth—played a role in the development of algal blooms in a study by Green and others (2021). Through a series of multivariate statistics, the parameters that played the most significant role in eutrophication for different reservoirs in the Eastern United States were prioritized. Because reservoirs had different bathymetries, the priority of the parameters varied, but the six parameters previously listed were the most common in driving the growth of phytoplankton. The depth of the reservoir played a major role in thermal stratification development, which set up conditions for internal nutrient recycling as discussed by Green and others (2021).
Knoll and others (2015) applied similar analysis to reservoir eutrophication and found that reservoirs with large catchment area-to-lake surface area ratios were particularly sensitive to eutrophication. They surmised that a larger drainage area has more potential for nutrient enrichment or more opportunity to warm waters flowing to the reservoir. The most important eutrophication influence identified by Knoll and others (2015) was land use and the transport of nutrients and sediments to the reservoirs. Their analysis included different types of agricultural land use and best management practices. Another study that focused on urbanizing watersheds found that increasing the impervious surfaces led to rapid, high velocity pulses of storm runoff. The high velocity pulses associated with urban storm runoff were capable of transporting sediments and nutrients to the receiving reservoir (Liu and others, 2021), making them susceptible to eutrophication.
Increasing Water Demands and Ecosystem Requirements from Growing Populations
Climate change is expected to bring more stress on water resources because of increased evapotranspiration and water demand. Changes in the distribution and quantity of, and demand for, water resources in response to climate variability and change are essential to understanding potential HAB occurrences. Population growth and warming conditions are expected to increase the demand for water for consumption, industry, and energy production (Frederick and Major, 1997; Lenton and others, 2023). An editorial in the journal Nature describes how the increasing temperatures, growing population, greater agricultural consumption, and higher evaporation will eventually put billions of people in water stress (Nature, 2023). After a study of the Tennessee River Basin, which encompasses parts of Tennessee, Virginia, North Carolina, Alabama, and Mississippi, the Tennessee Valley Authority projected that public water demand would increase by 12 percent from 2020 to 2045, and irrigation was projected to increase by 58 percent during the same timeframe (Sharkey and Springston, 2022). Other studies project a similar escalation in water demand in response to irrigation and livestock production; Wang and others (2016) describe conditions such as rising temperatures that will result in more water needs for crop production.
Just as raising crops and livestock in the future will require additional water, natural ecosystems may also require more water in response to increased risk of heat waves. Climate scientists used multiple GCMs to simulate the Earth's climate system and to develop plausible future scenarios derived with the Standardized Precipitation Evapotranspiration Index (SPEI) (Terando and others, 2020). The SPEI considers precipitation and potential evapotranspiration in determining potential drought conditions. Climatologists with the National Park Service, in collaboration with the USGS, used general circulation models, a specific type of GCM, to derive future SPEI scenarios at more than 370 parks across the United States (Monahan and Fisichelli, 2014; Battaglin and others, 2020). Running climate models for the geographically distributed park systems provides excellent regional perspective on water needs under different climate conditions. The climatologists used GCM model scenarios where mean temperatures increased 3 to 5 degrees Fahrenheit (°F), resulting in changing seasonal rain patterns and a sharp increase in evapotranspiration by the ecosystems during the summers, thereby drawing more water from the shallow aquifers and intercepting percolating rain waters.
LaFontaine and others (2019) also used GCMs, coupled with a USGS hydrologic model, to simulate water availability in the Southeastern United States under different climate conditions. They reported that runoff will generally decrease except in urban areas with impervious surfaces. Most summer streamflows were projected to be less than historical volumes across most of the Southeastern United States, with a few exceptions. The reduced streamflows across the study area were primarily affected by projected increases in air temperature and evapotranspiration, which can reduce the quantity of water storage available to generate runoff. This reduction in regional streamflow greatly affects the flushing of systems, and it will stabilize thermal stratification in many of the waterbodies. Thus, an increase in water demand and reduction in streamflow will lead to conditions that favor the development of HAB events. As more water is removed from reservoirs and decreased through increased evapotranspiration, less water will be available to maintain active flow through the rivers and to flush the lentic systems. Without adequate water to flush a system, the lentic systems become warmer, more stagnant, anoxic, and conducive to cyanobacteria growth (Jane and others, 2021; Rajesh and Rehana, 2022). The higher probability of stagnating waters will reinforce rising water temperatures and the onset of thermal stratification in deeper lakes and reservoirs, thus facilitating delivery of internal nutrients to cyanobacteria.
Other Physical and Chemical Drivers that Influence HAB Occurrence
Some additional physical and chemical drivers of HABs include the following: wind, pollution, groundwater, water circulation patterns, and variability in water depth and watershed properties. Wind can be a major factor influencing the occurrence of HABs; floating algal blooms can move across a lake, pushed by winds across the open waters, or they can be forced into the water column by turbulence (Xue and others, 2022). Rusak and others (2018) studied the effects of temperature, wind speed, and radiation on phytoplankton. They reported that temperature set up the growing conditions, but variation in wind speed was a major factor in phytoplankton abundance at all temporal scales. Sustained winds eroded density stratification, deepened mixing, and altered nutrient distribution, thus creating a complex circulation pattern resulting in intense patches of algae growth. Surface accumulation of cyanobacteria may develop and dissipate within a matter of hours as wind and weather conditions change, resulting in extreme spatial and temporal inconsistency.
Unknown Factors
Additional factors, such as urban heat disparity, could play a role in HAB development. Studies have shown that parts of an urban core can be up to 7 °C (12.6 °F) hotter than nearby suburban and rural areas with trees (Hoffman and others, 2020; Hsu and others, 2021; Witze, 2021). These urban heat traps lead to earlier temperature increases in urban ponds as compared to similar ponds in rural areas, thus allowing HABs to commence in urban areas during winter months. Few studies have focused on HAB occurrences in small, urban waterbodies, which results in a poor understanding of the relation between HABs and these conditions.
Results of one study to monitor HABs in urban environments in Kentucky indicated that the toxin-producing cyanobacteria were detected year-round (de la Cruz and others, 2017). In this study, ponds with heavy blooms also contained little to no buffer zones, whereas the ponds that had lily pads and a shoreline lined with trees, grasses, and riparian vegetation had less severe HAB events, possibly caused by increased nutrient uptake by vegetation. Additional research on urban HABs and competition with shading plants could help determine if the main driver is temperature, nutrient runoff, internal loading, migrating waterfowl, or some other parameter that facilitates the development of HABs in small ponds. Small ponds appear to be more vulnerable to HAB development than large waterbodies because the chemistry and biology can change rapidly in response to short weather patterns (de la Cruz and others, 2017), but they can also be treated much quicker and easier than a large body of water (ITRC, 2022).
Development of Models
Models are often used to describe potential future outcomes. The term “model” has been used in many ways to describe the development of HAB events or other environmental effects of climate change. A model output may be conveyed through various data visualization techniques to help communicate the results in compelling ways that allow people to use the information for education or decision making (USGS, 2019). Following are short descriptions of model types.
Types of Models
One of the most basic model types is a conceptual model. Developing a conceptual model involves reviewing and synthesizing relevant literature and then depicting the parameters, usually in a schematic or a condensed format. An example of a conceptual model for parameters that influence the development of HABs in freshwater is shown in figure 9. Conceptual models often generalize information, rarely include any quantification, and can be adapted to fit different physical settings or timelines (Beaver and others, 2018; Burford and others, 2020). They can provide a good starting point for projects but are usually not suited for supporting long-term management decisions needed to address changing climate variables. For that, more rigorous predictive models, such as statistical or mechanistic models, are needed (Safaie and others, 2016). Moving from a conceptual HAB model to a statistical or mechanistic model can be challenging and requires skills in mathematics and computer programming and an understanding of the system being modeled.
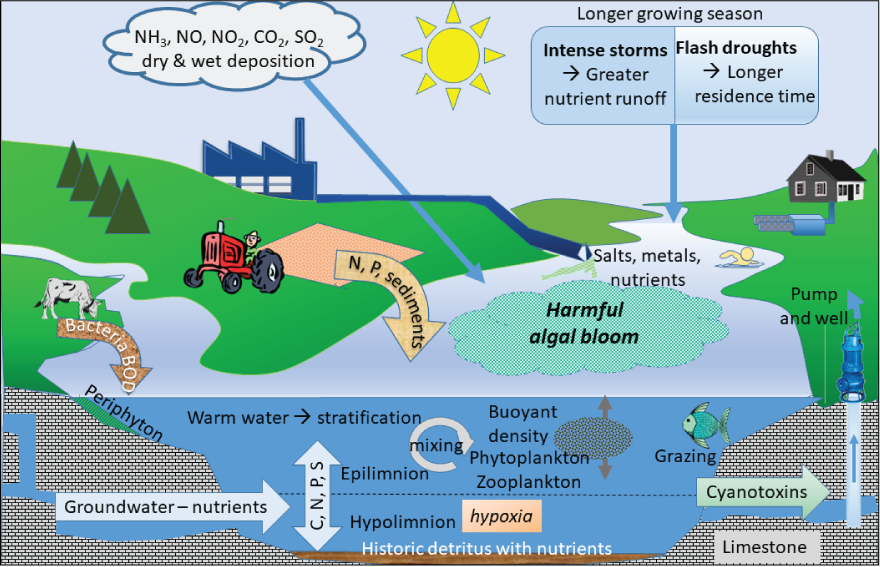
Conceptual model of parameters that influence the development of harmful algal blooms in freshwater. Notice the rates are not quantified and parameters are not prioritized in this conceptual model. (Adapted from Graham and others, 2016.) [NH3, ammonia; NO, nitric oxide; NO2, nitrogen dioxide; CO2, carbon dioxide; SO2, sulfur dioxide; C, carbon; N, nitrogen; P, phosphorous; S, sulfur; BOD, biological oxygen demand]
The fundamental difference between statistical models and mechanistic models is that statistical models use large amounts of data to look for patterns that can be described with probabilistic equations (Francy and others, 2020) and are not based on cause-and-effect mechanisms (Baker and others, 2018), whereas a mechanistic model is based on understanding the mechanisms that elicit a response in the target species. Statistical models apply algorithms to data to provide probabilistic results (Francy and others, 2020). Recently, statistical modelers have been applying machine learning and artificial intelligence to identify trends and predict potential patterns in HABs from large datasets (Song and others, 2015). An important trait of statistical models is that the outcome is based on correlation patterns and not causality. The output from statistical models is often bound to their calibration range, and the predicted results can be limited by the data breadth from which they are calibrated.
A mechanistic model is built upon a detailed understanding of the parameters in an ecosystem which can then be represented through mathematical equations. A mechanistic model compartmentalizes the different parameters, allows a user to change one or more of the physical or chemical factors, and mimics real-life events through suppositions on the prominent underlying mechanisms. For example, cyanobacteria growth rates can be evaluated in response to controlled variables, such as specific nutrient loads entering the waters, temperature changes, and other physical or chemical parameters that may influence the target species. The drawback of mechanistic models is that they require very detailed characterization and understanding of complex ecosystems, which may not be practical or achievable with limited resources and time. For example, cyanobacteria will interact with other algae, macrophytes, or grazers and will respond to changing water chemistry, all of which influence growth patterns of HAB species. It is technically infeasible to currently account for all possible mechanistic interactions with genetic diversity influencing cyanobacteria growth and production of toxins. Small- to large-scale studies continue to expand our understanding of the parameters involved in HAB development with the hope of developing a reliable mechanistic model (Glibert and Pitcher, 2001; GlobalHAB, 2021).
Statistical and mechanistic models have advantages and disadvantages. Many of the more recent hybrid models try to incorporate the best of both types of models by bringing in the causality missing from statistical approaches while building on the large datasets that are now readily available (Baker and others, 2018; fig. 10). Statistical and mechanistic modeling approaches rely on, and produce, different types of data and provide different types of understanding and output. Newer hybrid models incorporate the benefit of important hidden variables that may not be sufficiently quantified to insert in mechanistic models (Binder and others, 1997). The hybrid models incorporate mechanistic traits in the statistical outcomes based on data input. Some statistical modelers have been very intentional about identifying the cause-and-effect mechanisms by using multivariate approaches to determine which parameters have the greatest influence on the development of HABs in a system (Chapra and others, 2017; Francy and others, 2020; Ho and Michalak, 2020; Green and others, 2021). While statistical links are not definitive evidence of formal mechanistic association, the relation between large datasets and their potential for identifying mechanisms can expand our understanding of the influence of climate change on HAB development.
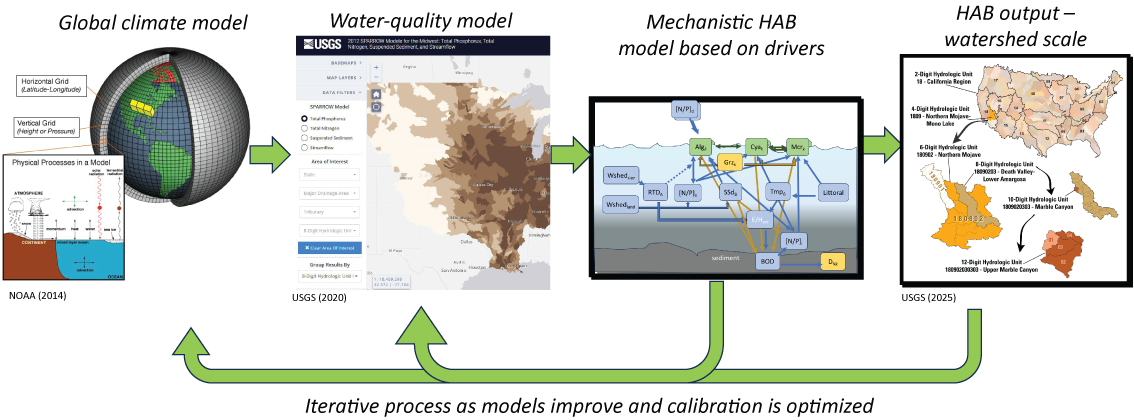
Conceptual diagram showing how combining scaled-down statistical global climate models under different climate scenarios with water-quality models and mechanistic harmful algal bloom (HAB) models can produce an iterative hybrid model that predicts HABs at different watershed scales. [NOAA, National Oceanic and Atmospheric Administration; USGS, U.S. Geological Survey]
Models that Predict HABs
Large datasets can be used to develop and evaluate statistical models that rank or prioritize parameters involved in HAB development on a large scale, and they can be used to predict the development of HABs (Beaver and others, 2014; Chapra and others, 2017; Harris and Graham, 2017; Franks, 2018; Burford and others, 2020; Ho and Michalak, 2020; Ralston and Moore, 2020; Green and others, 2021; Lin and others, 2023). The types of data used in these studies vary, and therefore, it is difficult to identify a definitive list of the top physical or chemical parameters involved in HAB development. However, these studies all conclude that nutrient availability played a major role in HAB occurrences. Because of this, most HAB forecasting models consider nutrients as a major driver of algae growth. A review of nutrient fate-and-transport models used to estimate eutrophication potential is provided by Morelli and others (2018). Some of the eutrophication models focused on rain, temperature, and nutrient loads, while others used land use, drainage basin size, bathymetry of the reservoir, impoundments on the rivers, photosynthetic pigment concentrations, salinity, or other factors to predict HABs. These multiple model parameters reflect the complex reality of HAB modeling. Partly because of the use of eutrophication data from large geographical areas and limited cyanobacteria information in the models, most models do not incorporate physiological traits for specific cyanobacteria genera, and instead only report concentrations of cyanobacteria.
Most HAB models are statistical models that base forecast development on regional knowledge concerning cyanobacteria response to area weather and reservoir conditions. These HAB models have proven effective for near-term forecasts by exploiting large, historical datasets that provide patterns of when and where the HAB will appear (Green and others, 2021). Remote sensing or field monitoring may be used to help detect and confirm HAB occurrences. As noted earlier, the numerical relations of statistical models become less predictable as conditions move outside the range of past observations, such as extreme heat waves, flash droughts, or 100-year rain events (Flynn and McGillicuddy, 2018). These climate-induced modifications to future weather patterns can be addressed in part by using a Bayesian statistical approach. In addition to incorporating historical information, Bayesian modeling is able to incorporate new knowledge into the model, such as the uncertainties of extreme weather or prior knowledge about the outcome (Moe and others, 2016). Bayesian modeling allows flexibility in dealing with missing extreme weather conditions that climate change will deliver. The model outcomes are expressed as a degree of belief in the potential solutions. Bayesian models are similar to mechanistic models in that they also allow the user to evaluate different management strategies to reduce the probability of HABs.
Mechanistic, or process-based, HAB models are typically more complex than statistical models, representing physical and biological mechanisms involved in HAB development, and they maintain authenticity when variable conditions extend outside of historical observation. A strong understanding of process variables is needed to be certain a mechanistic HAB model is accurate (Moore and others, 2008). With a mechanistic HAB model, one can compartmentalize processes important to algae growth, such as temperature, phosphorus, nitrogen, thermal stratification, and biological competition. Fennel and others (2011) developed a physical-biological model to describe eutrophication in the northern Gulf of America (formerly “Gulf of Mexico”) shelf. The model describes phytoplankton growth primarily as a function of nutrients delivered to the gulf from the Mississippi River and the Atchafalaya Basin. Nutrient and detritus loads from the rivers to the gulf, along with bathymetry, temperature, zooplankton grazing, nitrification, and other processes, were used to show that the increase in biomass is not simply a product of nutrients, but also a function of sediments and detritus transported to the gulf. By using a mechanistic model, Fennel and others (2011) were able to look at the environment to determine cause-and-effect relations. Ralston and Moore (2020) argued that mechanistic (process-based) models may be complicated, but they are worth developing for climate change HAB models because they can identify model parameters that are most sensitive to determining HAB development. Ralston and Moore (2020) recognized that development of mechanistic models requires numerous laboratory and ecosystem studies in a variety of settings to improve understanding of variables and process dependencies, including the effects of simultaneous changes to multiple driving factors. Ralston and Moore (2020, p. 6) also state: “Statistical approaches should continue to play an important role in HAB modeling, particularly for resource management and public health protection over event to seasonal time scales but extending statistical models to predict climate change response has limited merit.”
Several authors have discussed the need to incorporate more biological information, specifically cyanobacteria, competition, and predator-prey information, into HAB models (Wells and others, 2015; Glibert and Burford, 2017). Despite the limited and different information used in each HAB model study, several authors predicted HAB development for their respective geographical areas under current climate conditions with reasonable accuracy. Ralston and Moore report that several statistical models summarized in their review had less than 30 percent false positive. Fennel and others (2011) used a mechanistic model to predict algal blooms in the Gulf of America and had a corresponding coefficient of 64 percent. The model evaluations were done by using historical data to calibrate their models and then removing selected data to determine if the calibrated model predicted recent HAB events. Another way to test the model outcomes is to look back in geological history to when conditions were similar to current or future climate scenarios for evidence of HAB events. Several recent reports have quantified toxins, algae pigments, and other indicators of HABs in ancient lake sediments (Bianchi and others, 2000; Waters, 2016; Henao and others, 2020).
Water resource managers can use calibrated HAB models to identify parameters correlated with extreme algal growth and provide warnings at recreational sites, add supplemental treatment in water utilities, target specific land use for best management practices to reduce nutrient runoff in a watershed, and implement other actions that were previously infeasible. Future research could further explore whether these HAB models can be used in conjunction with climate models to project the probability of HABs based on future climate scenarios.
Combining Climate, Hydrology, and HAB Models
The Earth’s changing climate will have an outsized influence on HAB development (Wells and others, 2020). HABs are becoming more prevalent in more locations and with longer durations because of warming temperatures and human activities (National Research Council, 2020). Water resource managers are challenged to estimate the occurrence of HABs in this changing world and develop strategies to reduce HAB events. Models that accurately project HAB development based on a variety of climate scenarios could help water resource managers consider short- and long-term risks and opportunities. Integrated multiple models (multimodels) could be developed as tools for projecting potential future HAB events resulting from various climate scenarios and supporting decision making.
Even though uncertainty is associated with merging multiple models, incorporating the most recent climate information into the multimodel can improve the output. Examples of best practices associated with GCMs are discussed by Terando and others (2020). Some of their principal suggestions are as follows:
-
• Use a range of climate scenarios, such as a range of greenhouse gas emission scenarios, to account for human decisions that affect global greenhouse gas emissions.
-
• Use multiple climate models to project future effects resulting from specific scenarios to avoid regional biases of certain models.
-
• Run models to project future effects over timescales ranging from the near-term out to a century or longer, depending on the management decisions the user is trying to ascertain.
-
• Articulate clearly the uncertainties associated with any analysis.
-
• Ensure that climate-related observations and projections continue to receive rigorous peer review.
-
• Incorporate new knowledge and best practices as climate and HAB models improve.
The concept of merging climate models with other models to develop potential future scenarios under different climate conditions has been employed for a variety of environmental issues. LaFontaine and others (2019) integrated climate and hydrology models to predict streamflow and water resource conditions in the Southeastern United States under future climate scenarios. Gao and others (2021) combined climate models with hydrology and fire frequency models to establish the probability of drought conditions that could foster forest fires under different climate scenarios. Steinacher and others (2010) combined climate models and a primary production model to study the potential increase and decrease in productivity in the marine environment. Chapra and others (2017) merged climate models, a nutrient runoff model, and an algal growth model to predict HABs under different climate conditions for moderately large watersheds. These hybrid models are combinations of statistical and mechanistic models. Each of these integrated model studies started by using multiple GCMs which provided data on potential temperature, rain, evapotranspiration, rain intensity estimates, and drought intensity. Multiple GCMs were run in parallel to reduce any bias that might be present in a single GCM and to get a broader spectrum of potential outcomes. GCMs tend to use large grid cells, and the output may not be usable on a modest-sized watershed. Recent advances in technology allow GCM models to use additional information to down-scale and refine the output to smaller scales.
LaFontaine and others (2015) provide a detailed example of how a GCM output was applied to a hydrology model that incorporated land cover, geology, and topographic information. They used the USGS Precipitation-Runoff Modeling System (PRMS; Leavesley and others, 1983; Markstrom and others, 2015; Van Metre and others, 2019), which is a deterministic, distributed-parameter, process-based model system that can be integrated with the different GCM outputs to simulate the effects of precipitation, air temperature, and land use on potential hydrologic possibilities. The GCM-PRMS hybrid model provided a description of the potential duration, frequency, magnitude, rate of change, and timing of streamflow conditions in multiple basins. The hydrologic information provided by the GCM-PRMS hybrid model would be sufficient for calibration and execution of water-quality models. The purpose of these water-quality models is to predict nutrient and algal growth conditions.
Analysis of a combination of water-quality, nutrient, and algae life-cycle models can be used to gain more information. Morelli and others (2018) provide a review of potential water-quality models that can be integrated with the hydrological model output and then combined with a mechanistic algae life-cycle model. Morelli and others (2018) briefly describe 19 water-quality models and give a detailed example using the Soil and Water Assessment Tool (SWAT), Water Quality Analysis Simulation Program (WASP), and AQUATOX nutrient models (Ambose and Wool, 2009; Neitsch and others, 2011; Clough, 2014; Park and Clough, 2014). AQUATOX and WASP are models developed by the EPA to estimate water-quality concentrations and fluxes in individual stream reaches and river networks for monitoring and regulatory purposes. SWAT is a watershed model developed by the U.S. Department of Agriculture to simulate runoff and fate-and-transport processes for soil and nutrients. The Spatially Referenced Regression on Watershed attributes (SPARROW) model is a statistical process-based watershed model developed by the USGS (Schwarz and others, 2006) to predict the fate and transport of nutrients based on land use, topography, hydrogeology, and weather patterns. SPARROW is prominently used because it can predict changes in nutrient loading in response to land use changes. Each of these nutrient transport models have strengths and weaknesses that should be considered when deciding which one to use for a specifically defined purpose.
Chapra and others (2017) used five GCMs as part of the Coupled Model Intercomparison Project Phase 5 and organized the climate results with the SPARROW nutrient model and a biophysical model called QUALIDAD that simulates primary production such as cyanobacteria growth. QUALIDAD uses simple mechanistic representations linked to temperature, nutrients, photosynthetically active radiation, and growth rates to simulate growth of diatoms, green algae, and cyanobacteria. One shortcoming of the QUALIDAD model is that it is limited to planktonic algae and does not consider periphyton or benthic algae, which can be important to HAB events.
Chapra and others (2017) provides an example of how to merge climate models, a water-quality model, and a biological model to derive potential HAB scenarios on a watershed scale. The first step was to use GCMs to estimate potential hydrologic conditions. Next, SPARROW was used to further resolve potential physical and chemical drivers, such as nutrient loading and thermal stratification caused by changes in weather patterns modifying the hydrology. Based on SPARROW’s projection of potential nutrient loading under different climate scenarios, the QUALIDAD model then estimated growth of phytoplankton and HAB events. In general, this hybrid model estimates that more HABs will occur because of climate change, especially in the South and Southeastern United States (Chapra and others, 2017). Chapra and others (2017) surmise that there is a greater density of reservoirs in the Southeastern United States that are vulnerable to evolving conditions suitable for HAB development. The watershed size used in the hybrid GCM-nutrient-biophysical model was the 8-digit Hydrologic Unit Code (HUC) (Alexander and others, 1999). An 8-digit HUC watershed is large enough to have more than one reservoir, so Chapra and others (2017) used the volume of water stored in each 8-digit HUC as a surrogate for the assemblage of reservoirs in the basins. A multimodel such as this one provides preliminary insight into potential HAB events and gives water resource managers an opportunity to consider management options to treat HABs.
Model development is an iterative process that continues to improve as new data and technology become available. Future studies could use updated GCMs combined with diverse hydrology models, nutrient fate-and-transport models, and updated biological process models to obtain greater resolution (fig. 11). These hybrid models will also need to consider other factors to improve model outcome, such as increased demand by humans and ecosystems, improved remote sensing, increased understanding of the complex biological system based on genetic and ecosystem studies, and unfamiliar extreme weather patterns.

Schematic diagram showing the cyclical process of using multimodels to accurately determine process-based harmful algal bloom (HAB) response as climate change progresses and as the observed ecosystems reflect that progression. Various mechanistic and statistical models are accumulated: global climate models (GCMs), Spatially Referenced Regression on Watershed attributes (SPARROW), QUALIDAD, and Soil and Water Assessment Tool (SWAT). After these models are run and outputs are produced, the results are used to then forecast the ecosystem’s response to various scenarios. After this, the HAB response within the affected ecosystem is assessed in a model. Uncertainty is evaluated to address accuracy, and, lastly, using the model data, a plan is created to affect HAB growth. Long-term observations in the ecosystem will help to determine which models will be accumulated in future multimodels designed to assess HABs.
Climate multimodels for predicting HAB events are still in the developmental stage and will have limits but are improving with new insights and technology. Communications with water resource scientists and engineers can help resource managers and reservoir operators remain aware of new developments associated with climate change models. However, a concerted effort to disseminate the information and models is needed. A survey of 355 water treatment managers indicated that for information on how to handle HABs they rely “most on State regulators (48 percent), professional associations (18 percent), and internal sources (15 percent) … and rely least on scientific organizations (4 percent) and Internet searches (3 percent)” (Treuer and others, 2021, p. 960). Disseminating the best available science and forecasts can help inform decisions made by water resource managers, such as reservoir operators. Coordination among natural resource managers, farmers, and researchers could facilitate the development of strategies to reduce nutrient runoff and other factors important in HAB formation. The ITRC and agencies, such as the USGS, EPA, U.S. Department of Agriculture, National Oceanic and Atmospheric Administration, and U.S. Army Corps of Engineers; State agencies; academic institutions; and many private companies continue to support and report on strategies to reduce HABs and improve the models (Christensen and others, 2024). Reliable HAB models that can be adapted to different systems can help inform development of strategies that reduce the hazards and socioeconomic effects of future HABs.
Summary
Studies have predicted that harmful algal blooms (HABs) will become more frequent as climate change progresses. The conditions that promote the growth of HABs—higher water temperatures, concentrations of nutrients, shifting winds causing new patterns on the surface water—all become more frequent as global temperatures increase. In this report, the physical and chemical drivers that support harmful algal bloom growth described in published research are discussed. The production of cyanotoxins, as well as the prevalence of the algal species that secrete them, are addressed in various global climate models. The complexity of the physical parameters that promote HAB occurrence demonstrates how multifaceted a mechanistic model would have to be in order to incorporate all variables.
The different potential global climate models that have been published and could be used are discussed extensively. The benefits and risks of statistical and mechanistic models used to predict HAB development are weighed, as varying situations fit one or another better. Currently, the most accurate models tend to be statistical models based on years of data. Unfortunately, a statistical model developed for one particular waterbody does not have strong transferability to another waterbody with different bathymetry or hydrology. Thus, there is a need for development of a HAB model that can be adapted to different lakes, wetlands, and rivers. Once the strengths and weaknesses of models are described, the concept of hybrid models should be explored. Hybrid models have the potential to manipulate and process variables and output complex data and could potentially predict HAB response to climate change with increased confidence and accuracy. These hybrid models and multimodels are being developed in order to find the best model that can weigh different factors and reach an accurate result. An issue that will be difficult to address with these models is the inability to calculate current stressors, such as human and ecosystem demand and unfamiliar weather patterns, in the model. The output from hybrid models and multimodels will continue to be improved and verified with enhanced remote sensing and should one day be a valuable resource in the prevention and monitoring of HABs. The accuracy of these models could result in minimal socioeconomic effects from HAB growth in the future.
References Cited
Alexander, R.B., Brakebill, J.W., Brew, R.E., and Smith, R.A., 1999, ERF1—Enhanced River Reach File 1.2: U.S. Geological survey data release, accessed May 28, 2024, at https://doi.org/10.5066/P93JFBOE.
Baker, R.E., Peña, J-M., Jayamohan, J., and Jérusalem, A., 2018, Mechanistic models versus machine learning, a fight worth fighting for the biological community?: Biology Letters, v. 14, no. 5, accessed August 7, 2023, at https://doi.org/10.1098/rsbl.2017.0660.
Battaglin, W., Hay, L., Lawrence, D.J., McCabe, G., and Norton, P., 2020, Baseline conditions and projected future hydro-climatic change in national parks in the conterminous United States: Water (Basel), v. 12, no. 6, 24 p., accessed August 3, 2023, at https://doi.org/10.3390/w12061704.
Beaver, J.R., Manis, E.E., Loftin, K.A., Graham, J.L., Pollard, A.I., and Mitchell, R.M., 2014, Land use patterns, ecoregion, and microcystin relationships in U.S. lakes and reservoirs—A preliminary evaluation: Harmful Algae, v. 36, p. 57–62, accessed September 18, 2023, at https://doi.org/10.1016/j.hal.2014.03.005.
Beversdorf, L.J., Miller, T.R., and McMahon, K.D., 2013, The role of nitrogen fixation in cyanobacterial bloom toxicity in a temperate, eutrophic lake: PLoS One, v. 8, no. 2, article e56103, 11 p., accessed February 13, 2018, at https://doi.org/10.1371/journal.pone.0056103.
Binder, J., Koller, D., Russell, S., and Kanazawa, K., 1997, Adaptive probabilistic networks with hidden variables: Machine Learning, v. 29, p. 213–244, accessed August 7, 2023, at https://doi.org/10.1023/A:1007421730016.
Boehlert, B., Strzepek, K.M., Chapra, S.C., Fant, C., Gebretsadik, Y., Lickley, M., Swanson, R., McCluskey, A., Neumann, J.E., and Martinich, J., 2015, Climate change impacts and greenhouse gas mitigation effects on U.S. water quality: Journal of Advances in Modeling Earth Systems, v. 7, no. 3, p. 1326–1338.
Breinlinger, S., Phillips, T.J., Haram, B.N., Mareš, J., Martínez-Yerena, J.A., Houzek, P., Sobotka, R., Henderson, W.M., Schmieder, P., Williams, S.M., Lauderdale, J.D., Wilde, H.D., Gerrin, W., Kust, A., Washington, J.W., Wagner, C., Geier, B., Liebeke, M., Enke, H., Niedermeyer, T.H.J., and Wilde, S.B., 2021, Hunting the eagle killer—A cyanobacterial neurotoxin causes vacuolar myelinopathy: Science, v. 371, no. 6536, 7 p., accessed March 29, 2021, at https://doi.org/10.1126/science.aax9050.
Briand, J.F., Jacquet, S., Bernard, C., and Humbert, J-F., 2003, Health hazards for terrestrial vertebrates from toxic cyanobacteria in surface water ecosystems: Veterinary Research, v. 34, no. 4, p. 361–377, accessed May 11, 2020, at https://doi.org/10.1051/vetres:2003019.
Brooks, B.W., Lazorchak, J.M., Howard, M.D.A., Johnson, M.V.V., Morton, S.L., Perkins, D.A.K., Reavie, E.D., Scott, G.I., Smith, S.A., and Steevens, J.A., 2016, Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems?: Environmental Toxicology and Chemistry, v. 35, no. 1, p. 6–13.
Burford, M.A., Carey, C.C., Hamilton, D.P., Huisman, J., Paerl, H.W., Wood, S.A., and Wulff, A., 2020, Perspective—Advancing the research agenda for improving understanding of cyanobacteria in a future of global change: Harmful Algae, v. 91, accessed October 19, 2021, at https://doi.org/10.1016/j.hal.2019.04.004.
Carpenter, S.R., Gahler, M.R., Kucharik, C.J., and Stanley, E.H., 2022, Long-range dependence and extreme values of precipitation, phosphorus load, and cyanobacteria: Proceedings of the National Academy of Sciences of the United States of America, v. 119, no. 48, 7 p., accessed July 28, 2023, at https://doi.org/10.1073/pnas.2214343119.
Carter, L., Terando, A.J., Dow, K., Hiers, K., Kunkel, K.E., Lascrain, A.R., Marcy, D., Osland, M.J., and Schramm, P., 2018, Southeast, chap. 19 of Reidmiller, D.R., Avery, C.W., Easterling, D.R., Kunkel, K.E., Lewis, K.L.M., Maycock, T.K., and Stewart, B.C., eds., Impacts, risks, and adaptation in the United States—Fourth National Climate Assessment, Volume II: Washington, D.C., U.S. Global Change Research Program, p. 743–808, accessed July 27, 2023, at https://doi.org/10.7930/NCA4.2018.
Centers for Disease Control and Prevention [CDC], 2023, Summary Report—One Health Harmful Algal Bloom System (OHHABS), United States, 2021: Atlanta, Ga., Centers for Disease Control and Prevention web page, accessed October 15, 2023, at https://www.cdc.gov/habs/data/2021-ohhabs-data-summary.html.
Chapra, S.C., Boehlert, B., Fant, C., Bierman, V.J., Jr., Henderson, J., Mills, D., Mas, D.M.L., Rennels, L., Jantarasami, L., Martinich, J., Strzepek, K.M., and Paerl, H.W., 2017, Climate change impacts on harmful algal blooms in U.S. freshwaters—A screening-level assessment: Environmental Science & Technology, v. 51, no. 16, p. 8933–8943.
Chorus, I., and Zessner, M., 2021, Assessing and controlling the risk of cyanobacterial blooms—Nutrient loads from the catchment, chap. 7 of Chorus, I., and Welker, M., eds., Toxic cyanobacteria in water (2d ed.): Geneva, Switzerland, World Health Organization, prepared by CRC Press, Boca Raton, Fla., p. 434–498.
Christensen, V.G., Crawford, C.J., Dusek, R.J., Focazio, M.J., Fogarty, L.R., Graham, J.L., Journey, C.A., Lee, M.E., Larson, J.H., Stackpoole, S.M., Mazzei, V., Pindilli, E.J., Rattner, B.A., Slonecker, T., McSwain, K.B., Reilly, T.J., and Lopez, A.E., 2024, Interdisciplinary science approach for harmful algal blooms (HABs) and algal toxins—A strategic science vision for the U.S. Geological Survey: U.S. Geological Survey Circular 1520, 39 p., accessed February 7, 2025, at https://doi.org/10.3133/cir1520.
Christian, J.I., Hobbins, M., Hoell, A., Otkin, J.A., Ford, T.W., Cravens, A.E., Powlen, K.A., Wang, H., and Mishra, V., 2024, Flash drought—A state of the science review: WIREs Water, article e1714, accessed March 28, 2024, at https://doi.org/10.1002/wat2.1714.
Davis, T., Harke, M., Marcoval, M., Goleski, J., Orano-Dawson, C., Berry, D., and Gobler, C., 2010, Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms: Aquatic Microbial Ecology, v. 61, no. 2, p. 149–162, accessed May 2, 2019, at https://doi.org/10.3354/ame01445.
de la Cruz, A., Logsdon, R., Lye, D., Guglielmi, S., Rice, A., and Kannan, M.S., 2017, 2017, Harmful algae bloom occurrence in urban ponds—Relationship of toxin levels with cell density and species composition: Journal of Earth and Environmental Sciences, v. 25, p. 704–726, accessed May 5, 2024, at https://pubmed.ncbi.nlm.nih.gov/31750421/.
Dittmann, E., Fewer, D.P., and Neilan, B.A., 2013, Cyanobacterial toxins—Biosynthetic routes and evolutionary root: FEMS Microbiology Reviews, v. 37, no. 1, p. 23–43, accessed May 15, 2020, at https://doi.org/10.1111/j.1574-6976.2012.12000.x.
Fennel, K., Hetland, R., Feng, Y., and DiMarco, S., 2011, A coupled physical-biological model of the Northern Gulf of Mexico shelf—Model description, validation and analysis of phytoplankton variability: Biogeosciences, v. 8, no. 7, p. 1881–1899, accessed October 17, 2023, at https://doi.org/10.5194/bg-8-1881-2011.
Flynn, K.J., and McGillicuddy, D.J., 2018, Modeling marine harmful algal blooms—Current status and future prospects, chap. 3 of Shumway, S.E., Burkholder, J.M., and Morton, S.L., eds., Harmful algal blooms—A compendium desk reference: Hoboken, N.J., John Wiley & Sons, Ltd, p. 115–134, accessed February 2, 2022, at https://doi.org/10.1002/9781118994672.ch3.
Francy, D.S., Brady, A.M.G., Stelzer, E.A., Cicale, J.R., Hackney, C., Dalby, H.D., Struffolino, P., and Dwyer, D.F., 2020, Predicting microcystin concentration action-level exceedances resulting from cyanobacterial blooms in selected lake sites in Ohio: Environmental Monitoring and Assessment, v. 192, no. 8, p. 513, accessed August 10, 2023, at https://doi.org/10.1007/s10661-020-08407-x.
Franks, P.J.S., 2018, Recent advances in modelling of harmful algal blooms, chap. 19 of Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., and Zhou, M., eds., Global ecology and oceanography of harmful algal blooms, v. 232 of Wardle, D.A., Canadell, J.G., Díaz, S., Heldmaier, G., Jackson, R.B., Levia, D.F., Schulze, E., and Sommer, U., eds., Ecological studies—Analysis and synthesis: Cham, Switzerland, Springer, p. 359–377, accessed July 18, 2023, at https://doi.org/10.1007/978-3-319-70069-4_19.
Fuentes, J.L., Garbayo, I., Cuaresma, M., Montero, Z., Gonzalez-del-Valle, M., and Vilchez, C., 2016, Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds: Marine Drugs, v. 14, no. 5, p. 100, accessed June 19, 2020, at https://doi.org/10.3390/md14050100.
Gao, P., Terando, A.J., Kupfer, J.A., Varner, J.M., Stambaugh, M.C., Lei, T.L., and Hiers, J.K., 2021, Robust projections of future fire probability for the conterminous United States: Science of the Total Environment, v. 789, article 147872, accessed July 27, 2023, at https://doi.org/10.1016/j.scitotenv.2021.147872.
Glibert, P.M., and Burford, M.A., 2017, Globally changing nutrient loads and harmful algal blooms—Recent advances, new paradigms, and continuing challenges: Oceanography, v. 30, no. 1, p. 58–69, accessed August 10, 2023, at https://doi.org/10.5670/oceanog.2017.110.
Glibert, P.M., and Burkholder, J.M., 2006, The complex relationships between increases in fertilization of the earth, coastal eutrophication and proliferation of harmful algal blooms, chap. 26 of Granéli, E., and Turner, J.T., eds., Ecology of harmful algae, v. 189 of Wardle, D.A., Canadell, J.G., Díaz, S., Heldmaier, G., Jackson, R.B., Levia, D.F., Schulze, E., and Sommer, U., eds., Ecological studies—Analysis and synthesis: Berlin, Springer, p. 341–354.
Gobler, C.J., 2020, Climate change and harmful algal blooms—Insights and perspective: Harmful Algae, v. 91, 4 p., article 101731, accessed May 2, 2019, at https://doi.org/10.1016/j.hal.2019.101731.
Gobler, C.J., Doherty, O.M., Hattenrath-Lehmann, T.K., Griffith, A.W., Kang, Y., and Litaker, R.W., 2017, Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans: Proceedings of the National Academy of Sciences of the United States of America, v. 114, no. 19, p. 4975–4980, accessed December 17, 2019, at https://doi.org/10.1073/pnas.1619575114.
Graham, J.L., Dubrovsky, N.M., and Eberts, S.M., 2016, Cyanobacterial harmful algal blooms and U.S. Geological Survey science capabilities: U.S. Geological Survey Open-File Report 2016–1174, 12 p., accessed January 17, 2017, at https://doi.org/10.3133/ofr20161174.
Graham, J.L., Ziegler, A.C., Loving, B.L., and Loftin, K., 2012, Fate and transport of cyanobacteria and associated toxins and taste-and-odor compounds from upstream reservoir releases in the Kansas River, Kansas, September and October 2011: U.S. Geological Survey Scientific Investigations Report 2012–5129, 65 p.
Green, W.R., Hoos, A.B., Wilson, A.E., and Heal, E.N., 2021, Development of a screening tool to examine lake and reservoir susceptibility to eutrophication in selected watersheds of the eastern and southeastern United States: U.S. Geological Survey Scientific Investigations Report 2021–5075, 59 p., accessed November 1, 2022, at https://doi.org/10.3133/sir20215075.
Griffith, A.W., and Gobler, C.J., 2020, Harmful algal blooms—A climate change co-stressor in marine and freshwater ecosystems: Harmful Algae, v. 91, article 101590, accessed May 20, 2023, at https://doi.org/10.1016/j.hal.2019.03.008.
Grogan, A.E., Alves-de-Souza, C., Cahoon, L.B., and Mallin, M.A., 2023, Harmful algal blooms—A prolific issue in urban stormwater ponds: Water, v. 15, article 2436, accessed May 9, 2024, at https://doi.org/10.3390/w15132436.
Gudmundsson, L., Boulange, J., Do, H.X., Gosling, S.N., Grillakis, M.G., Koutroulis, A.G., Leonard, M., Liu, J., Schmied, H.M., Papadimitriou, L., Pokhrel, Y., Seneviratne, S.I., Satoh, Y., Thiery, W., Westra, S., Zhang, X., and Zhao, F., 2021, Globally observed trends in mean and extreme river flow attributed to climate change: Science, v. 371, no. 6534, p. 1159–1162.
Hallegraeff, G.M., Anderson, D.M., Belin, C., Bottein, M.Y.D., Bresnan, E., Chinain, M., Enevoldsen, H., Iwataki, M., Karlson, B., McKenzie, C.H., Sunesen, I., Pitcher, G.C., Provoost, P., Richardson, A., Schweibold, L., Tester, P.A., Trainer, V.L., Yñiguez, A.T., and Zingone, A., 2020, Perceived global increase in algal blooms is attributable to intensified monitoring and emerging bloom impacts: Communications Earth & Environment, v. 2, article 117, accessed June 11, 2021, at https://doi.org/10.1038/s43247-021-00178-8.
Harp, R.D., and Horton, D.E., 2022, Observed changes in daily precipitation intensity in the United States: Geophysical Research Letters, v. 49, no. 19, article e2022GL099955, accessed August 3, 2023, at https://doi.org/10.1029/2022GL099955.
Harris, T.D., and Graham, J.L., 2017, Predicting cyanobacterial abundance, microcystin, and geosmin in a eutrophic drinking-water reservoir using a 14-year dataset: Lake and Reservoir Management, v. 33, no. 1, p. 32–48, accessed May 10, 2018, at https://doi.org/10.1080/10402381.2016.1263694.
Harris, T.D., Wilhelm, F.M., Graham, J.L., and Loftin, K.A., 2014, Experimental manipulation of TN:TP ratios suppress cyanobacterial biovolume and microcystin concentration in large-scale in situ mesocosms: Lake and Reservoir Management, v. 30, no. 1, p. 72–83, accessed June 1, 2015, at https://doi.org/10.1080/10402381.2013.876131.
Henao, E., Rzymski, P., and Waters, M.N., 2020, A review on the study of cyanotoxins in paleolimnological research—Current knowledge and future needs: Toxins, v. 12, no. 1, article 6, accessed September 29, 2021, at https://doi.org/10.3390/toxins12010006.
Hesse, P.R., 1975, Blue-green algae for rice production: Food and Agriculture Organization of the United Nations, Soils Bulletin 46, RAS/75/004, [variously paged], accessed February 13, 2019, at https://www.fao.org/3/ar124e/ar124e.pdf.
Ho, J.C., Michalak, A.M., and Pahlevan, N., 2019, Widespread global increase in intense lake phytoplankton blooms since the 1980s: Nature, v. 574, p. 667–670, accessed December 10, 2021, at https://doi.org/10.1038/s41586-019-1648-7.
Hoffman, J.S., Shandas, V., and Pendleton, N., 2020, The effects of historical housing policies on resident exposure to intra-urban heat—A study of 108 US urban areas: Climate, v. 8, no. 1, article 12, accessed December 10, 2021, at https://doi.org/10.3390/cli8010012.
Interstate Technology & Regulatory Council [ITRC], 2022, Strategies for preventing and managing benthic harmful cyanobacterial blooms (HCB-2): Washington, D.C., Interstate Technology & Regulatory Council, HCB Team, accessed July 24, 2023, at https://hcb-2.itrcweb.org/.
Jane, S.F., Hansen, G.J.A., Kraemer, B.M., Leavitt, P.R., Mincer, J.L., North, R.L., Pilla, R.M., Stetler, J.T., Williamson, C.E., Woolway, R.I., Arvola, L., Chandra, S., DeGasperi, C.L., Diemer, L., Dunalska, J., Erina, O., Flaim, G., Grossart, H.P., Hambright, K.D., Hein, C., Hejzlar, J., Lanus, L.L., Jenny, J.P., Jones, J.R., and Rose, K.C., 2021, Widespread deoxygenation of temperate lakes: Nature, v. 594, p. 66–70, accessed January 7, 2022, at https://doi.org/10.1038/s41586-021-03550-y.
Ji, X., Verspagen, J.M.H., Van de Waal, D.B., Rost, B., and Huisman, J., 2020, Phenotypic plasticity of carbon fixation stimulates cyanobacterial blooms at elevated CO2: Science Advances, v. 6, no. 8, accessed October 5, 2022, at https://doi.org/10.1126/sciadv.aax2926.
Journey, C.A., Beaulieu, K.M., and Bradley, P.M., 2013, Environmental factors that influence cyanobacteria and geosmin occurrence in reservoirs, chap. 2 of Bradley, P.M., ed., Current perspectives in containment hydrology and water resources sustainability: IntechOpen, p. 27–55, accessed February 13, 2019, at https://doi.org/10.5772/54807.
Kaebernick, M., and Neilan, B.A., 2001, Ecological and molecular investigations of cyanotoxin production: FEMS Microbiology Ecology, v. 35, no. 1, p. 1–9, accessed December 15, 2017, at https://doi.org/10.1111/j.1574-6941.2001.tb00782.x.
Knoll, L.B., Hagenbuch, E.J., Stevens, M.H., Vanni, M.J., Renwick, W.H., Denlinger, J.C., Hale, R.S., and González, M.J., 2015, Predicting eutrophication status in reservoirs at large spatial scales using landscape and morphometric variables: Inland Waters, v. 5, no. 3, p. 203–214, accessed November 1, 2022, at https://doi.org/10.5268/IW-5.3.812.
LaFontaine, J.H., Hart, R.M., Hay, L.E., Farmer, W.H., Bock, A.R., Viger, R.J., Markstrom, S.L., Regan, R.S., and Driscoll, J.M., 2019, Simulation of water availability in the Southeastern United States for historical and potential future climate and land-cover conditions: U.S. Geological Survey Scientific Investigations Report 2019–5039, 83 p., accessed August 8, 2023, at https://doi.org/10.3133/sir20195039.
LaFontaine, J.H., Hay, L.E., Viger, R.J., Regan, V.R., and Markstrom, S.L., 2015, Effects of climate and land cover on hydrology in the Southeastern U.S.—Potential impacts on watershed planning: Journal of the American Water Resources Association, v. 51, no. 5, p. 1235–1261, accessed August 3, 2023, at https://doi.org/10.1111/1752-1688.12304.
Lehmann, J., Coumou, D., and Frieler, K., 2015, Increased record-breaking precipitation events under global warming: Climatic Change, v. 132, p. 501–515, accessed July 27, 2023, at https://doi.org/10.1007/s10584-015-1434-y.
Lenton, T.M., Xu, C., Abrams, J.F., Ghadiali, A., Loriani, S., Sakschewski, B., Zimm, C., Ebi, K.L., Dun, R.R., Svenning, J.C., and Scheffer, M., 2023, Quantifying the human cost of global warming: Nature Sustainability, v. 6, p. 1237–1247, accessed May 28, 2024, at https://doi.org/10.1038/s41893-023-01132-6.
Leung, T., Wilkinson, G.M., and Swanner, E.D., 2021, Iron availability allows sustained cyanobacterial blooms—A dual-lake case study: Inland Waters, v. 11, no. 3, p. 417–429, accessed February 13, 2022, at https://doi.org/10.1080/20442041.2021.1904762.
Lin, S., Pierson, D.C., and Mesman, J.P., 2023, Prediction of algal blooms via data-driven machine learning models—An evaluation using data from a well-monitored mesotrophic lake: Geoscientific Model Development, v. 16, no. 1, p. 35–46, accessed September 18, 2023, at https://doi.org/10.5194/gmd-16-35-2023.
Liu, S., Johnson, F., Tamburic, B., Crosbie, N.D., and Glamore, W., 2021, The effectiveness of global constructed shallow waterbody design guidelines to limit harmful algal blooms: Water Resources Research, v. 57, no. 8, article e2020WR028918, accessed June 14, 2022, at https://doi.org/10.1029/2020WR028918.
Maavara, T., Chen, Q., Van Meter, K., Brown, L.E., Zhang, J., Ni, J., and Zarfl, C., 2020, River dam impacts on biogeochemical cycling: Nature Reviews Earth & Environment, v. 1, p. 103–116, accessed February 7, 2020, at https://doi.org/10.1038/s43017-019-0019-0.
Manganelli, M., Scardala, S., Stefanelli, M., Palazzo, F., Funari, E., Vichi, S., Buratti, F.M., and Testai, E., 2012, Emerging health issues of cyanobacterial blooms: Ann Ist Super Sanità, v. 48, no. 4, p. 415–428, accessed April 8, 2022, at https://doi.org/10.4415/ANN_12_04_09.
Markstrom, S.L., Regan, R.S., Hay, L.E., Viger, R.J., Webb, R.M.T., Payn, R.A., and LaFontaine, J.H., 2015, PRMS-IV, the Precipitation-Runoff Modeling System, version 4: U.S. Geological Survey Techniques and Methods, book 6, chap. B7, 158 p., accessed February 13, 2022, at https://doi.org/10.3133/tm6B7.
Metcalf, J.S., Tischbein, M., Cox, P.A., and Stommel, E.W., 2021, Cyanotoxins and the nervous system: Toxins, v. 13, no. 9, article 660, accessed September 18, 2023, at https://doi.org/10.3390/toxins13090660.
Moe, S.J., Haande, S., and Couture, R.M., 2016, Climate change, cyanobacteria blooms and ecological status of lakes—A Bayesian network approach: Ecological Modelling, v. 337, p. 330–347, accessed September 18, 2023, at https://doi.org/10.1016/j.ecolmodel.2016.07.004.
Mohammadi, K., Jiang, Y., and Wang, G., 2022, Flash drought early warning based on the trajectory of solar-induced chlorophyll fluorescence: Proceedings of the National Academy of Sciences of the United States of America, v. 119, no. 32, article e2202767119, accessed July 20, 2023, at https://doi.org/10.1073/pnas.2202767119.
Monahan, W.B., and Fisichelli, N.A., 2014, Climate exposure of US national parks in a new era of change: PLoS One, v. 9, no. 7, article e101302, accessed August 3, 2023, at https://doi.org/10.1371/journal.pone.0101302.
Moore, S.K., Trainer, V.L., Mantua, N.J., Parker, M.S., Laws, E.A., Backer, L.C., and Fleming, L.E., 2008, Impacts of climate variability and future climate change on harmful algal blooms and human health: Environmental Health, v. 7, suppl. 2, article S4, accessed December 10, 2021, at https://doi.org/10.1186/1476-069X-7-S2-S4.
Morelli, B., Hawkins, T.R., Niblick, B., Henderson, A.D., Golden, H.E., Compton, J.E., Cooter, E.J., and Bare, J.C., 2018, Critical review of eutrophication models for life cycle assessment: Environmental Science & Technology, v. 52, no. 17, p. 9562–9578, accessed August 11, 2023, at https://doi.org/10.1021/acs.est.8b00967.
Morton, N., Hogan, B., Toomey, R., Young, D., and Byl, T.D., 2020, Characterizing the pigments in Lampenflora in Mammoth Cave National Park, Kentucky, in Proceedings of the 2020 Tennessee Water Resources Symposium, Burns, Tenn., April 22–24, 2020: Tennessee Section of the American Water Resources Association, p. P-21.
Munn, M.D., Frey, J.W., Tesoriero, A.J., Black, R.W., Duff, J.H., Lee, K., Maret, T.R., Mebane, C.A., Waite, I.R., and Zelt, R.B., 2018, Understanding the influence of nutrients on stream ecosystems in agricultural landscapes: U.S. Geological Survey Circular 1437, 80 p., accessed May 28, 2024, at https://doi.org/10.3133/cir1437.
National Oceanic and Atmospheric Administration, 2014, Climate models: National Oceanic and Atmospheric Administration web page, accessed March 3, 2025, at https://www.climate.gov/maps-data/climate-data-primer/predicting-climate/climate-models.
National Oceanic and Atmospheric Administration, [2023], Future climate projections—Graphs and maps—U.S. Climate Resilience Toolkit—Climate Explorer: National Oceanic and Atmospheric Administration web page, accessed July 25, 2023, at https://www.climate.gov/maps-data/dataset/future-climate-projections-graphs-maps.
National Park Service, 2023, Park-specific climate science: National Park Service web page, accessed July 25, 2023, at https://www.nps.gov/subjects/climatechange/parkclimatescience.htm.
National Research Council, 2013, Abrupt impacts of climate change—Anticipating surprises: Washington, D.C., The National Academies Press, 222 p., accessed January 7, 2021, at https://doi.org/10.17226/18373.
National Research Council, 2020, Climate change—Evidence and causes—Update 2020: Washington, D.C., The National Academies Press, 36 p., accessed July 27, 2023, at https://doi.org/10.17226/25733.
Nature, 2023, Water crisis—How local technologies can help solve a global problem (editorial): Nature, August 1, 2023, v. 620, no. 7, accessed September 7, 2023, at https://doi.org/10.1038/d41586-023-02442-7.
Neilan, B.A., Pearson, L.A., Muenchhoff, J., Moffitt, M.C., and Dittmann, E., 2013, Environmental conditions that influence toxin biosynthesis in cyanobacteria: Environmental Microbiology, v. 15, no. 5, p. 1239–1253, accessed May 15, 2020, at https://doi.org/10.1111/j.1462-2920.2012.02729.x.
Paerl, H.W., Hall, N.S., Hounshell, A.G., Luettich, R.A., Jr., Rossignol, K.L., Osburn, C.L., and Bales, J., 2019, Recent increase in catastrophic tropical cyclone flooding in coastal North Carolina, USA—Long-term observations suggest a regime shift: Scientific Reports, v. 9, article 10620, accessed July 27, 2023, at https://doi.org/10.1038/s41598-019-46928-9.
Paerl, H.W., and Otten, T.G., 2013, Harmful cyanobacterial blooms—Causes, consequences, and controls: Microbial Ecology, v. 65, p. 995–1010, accessed May 26, 2023, at https://doi.org/10.1007/s00248-012-0159-y.
Paerl, H.W., Scott, J.T., McCarthy, M.J., Newell, S.E., Gardner, W.S., Havens, K.E., Hoffman, D.K., Wilhelm, S.W., and Wurtsbaugh, W.A., 2016, It takes two to tango—When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems: Environmental Science & Technology, v. 50, no. 20, p. 10805–10813, accessed May 28, 2024, at https://doi.org/10.1021/acs.est.6b02575.
Popall, R.M., Bolhuis, H., Muyzer, G., and Sánchez-Román, M., 2020, Stromatolites as biosignatures of atmospheric oxygenation—Carbonate biomineralization and UV-C resilience in a Geitlerinema sp.-dominated culture: Frontiers in Microbiology, v. 11, article 948, accessed July 19, 2023, at https://doi.org/10.3389/fmicb.2020.00948.
Pradhan, B., Nayak, R., Patra, S., Jit, B.P., Ragusa, A., and Jena, M., 2021, Bioactive metabolites from marine algae as potent pharmacophores against oxidative stress-associated human diseases—A comprehensive review: Molecules (Basel, Switzerland), v. 26, no. 1, article 37, accessed December 6, 2022, at https://doi.org/10.3390/molecules26010037.
Rajesh, M., and Rehana, S., 2022, Impact of climate change on river water temperature and dissolved oxygen—Indian riverine thermal regimes: Scientific Reports, v. 12, article 9222, accessed June 13, 2023, at https://doi.org/10.1038/s41598-022-12996-7.
Rantala, A., Fewer, D.P., Hisbergues, M., Rouhiainen, L., Vaitomaa, J., Börner, T., and Sivonen, K., 2004, Phylogenetic evidence for the early evolution of microcystin synthesis: Proceedings of the National Academy of Sciences of the United States of America, v. 101, no. 2, p. 568–573, accessed January 10, 2018, at https://doi.org/10.1073/pnas.0304489101.
Rohwer, R.R., Ladwig, R., Dugan, H.A., Hanson, P.C., Walsh, J.R., and Zanden, M.J.V., 2023, The aftermath of a trophic cascade—Increased anoxia following species invasion of a eutrophic lake: bioRχiv, 15 p., accessed September 18, 2023, at https://doi.org/10.1101/2023.01.27.525925.
Rusak, J.A., Tanentzap, A.J., Klug, J.L., Rose, K.C., Hendricks, S.P., Jennings, E., Laas, A., Pierson, D., Ryder, E., Smyth, R.L., White, D.S., Winslow, L.A., Adrian, R., Arvola, L., de Eyto, E., Feuchtmayr, H., Honti, M., Istvánovics, V., Jones, D., McBride, C.G., Schmidt, S.R., Seekell, D., Staehr, P.A., and Zhu, G., 2018, Wind and trophic status explain within and among-lake variability of algal biomass: Limnology and Oceanography Letters, v. 3, no. 6, p. 409–418, accessed August 3, 2023, at https://doi.org/10.1002/lol2.10093.
Ryberg, K.R., and Chanat, J.G., 2022, Climate extremes as drivers of surface-water-quality trends in the United States: Science of the Total Environment, v. 809, article 152165, accessed January 7, 2022, at https://doi.org/10.1016/j.scitotenv.2021.152165.
Santer, B.D., Mears, C., Wentz, F.J., Taylor, K.E., Glecker, P.J., Wigley, T.M.L., Barnett, T.P., Boyle, J.S., Bruggemann, W., Gillett, N.P., Klein, S.A., Meehl, G.A., Nozawa, T., Pierce, D.W., Stott, P.A., Washington, W.M., and Wehner, M.F., 2007, Identification of human-induced changes in atmospheric moisture content: Proceedings of the National Academy of Sciences of the United States of America, v. 104, no. 39, p. 15248–15253.
Sharkey, J.K., and Springston, G.L., 2022, Water use in the Tennessee Valley for 2020 and projected use in 2045: Tennessee Valley Authority, River and Resources Stewardship, 80 p., accessed July 31, 2023, at https://tva-azr-eastus-cdn-ep-tvawcm-prd.azureedge.net/cdn-tvawcma/docs/default-source/default-document-library/site-content/environment/managing-the- river/water-use-in-the-tennessee-valley-for-2020-and-projected-use-in-2045.pdf?sfvrsn=baf315a5_2.
Sinha, E., Michalak, A.M., and Balaji, V., 2017, Eutrophication will increase during the 21st century as a result of precipitation changes: Science, v. 357, no. 6349, p. 405–408, accessed July 28, 2017, at https://doi.org/10.1126/science.aan2409.
Song, W., Dolan, J.M., Cline, D., and Xiong, G., 2015, Learning-based algal bloom event recognition for oceanographic decision support system using remote sensing data: Remote Sensing (Basel), v. 7, no. 10, p. 13564–13585, accessed April 28, 2016, at https://doi.org/10.3390/rs71013564.
Steinacher, M., Joos, F., Frölicher, T.L., Bopp, L., Cadule, P., Cocco, V., Doney, S.C., Gehlen, M., Lindsey, K., Moore, J.K., Schneider, B., and Segschneider, J., 2010, Projected 21st century decrease in marine productivity—A multi-model analysis: Biogeosciences, v. 7, no. 3, p. 979–1005, accessed August 15, 2023, at https://doi.org/10.5194/bg-7-979-2010.
Summers, E.J., and Ryder, J.L., 2024, A modified stratification index method to assess reservoir water quality trends: Journal of Environmental Management, v. 354, article 120297, accessed February 21, 2024, at https://doi.org/10.1016/j.jenvman.2024.120297.
Sun, Q., Zhang, X., Zwiers, F., Westra, S., and Alexander, L.V., 2021, A global, continental, and regional analysis of changes in extreme precipitation: Journal of Climate, v. 34, no. 1, p. 243–258, accessed July 27, 2023, at https://doi.org/10.1175/JCLI-D-19-0892.1.
Tabari, H., 2020, Climate change impact on flood and extreme precipitation increases with water availability: Scientific Reports, v. 10, article 13768, accessed June 6, 2022, at https://doi.org/10.1038/s41598-020-70816-2.
Tanvir, R.U., Hu, Z., Zhang, Y., and Lu, J., 2021, Cyanobacterial community succession and associated cyanotoxin production in hypereutrophic and eutrophic freshwaters: Environmental Pollution, v. 290, article 118056, accessed September 18, 2023, at https://doi.org/10.1016/j.envpol.2021.118056.
Terando, A., Reidmiller, D., Hostetler, S.W., Littell, J.S., Beard, T.D., Jr., Weiskopf, S.R., Belnap, J., and Plumlee, G.S., 2020, Using information from global climate models to inform policymaking—The role of the U.S. Geological Survey: U.S. Geological Survey Open-File Report 2020–1058, 25 p., accessed July 28, 2021, at https://doi.org/10.3133/ofr20201058.
Toreti, A., Naveau, P., Zampieri, M., Schindler, A., Scoccimarro, E., Xoplaki, E., Dijkstra, H.A., Gualdi, S., and Luterbacher, J., 2013, Projections of global changes in precipitation extremes from coupled model intercomparison project phase 5 models: Geophysical Research Letters, v. 40, no. 18, p. 4887–4892, accessed June 1, 2015, at https://doi.org/10.1002/grl.50940.
Treuer, G., Kirchhoff, C., Lemos, M.C., and McGrath, F., 2021, Challenges of managing harmful algal blooms in US drinking water systems: Nature Sustainability, v. 4, p. 958–964, accessed August 15, 2023, at https://doi.org/10.1038/s41893-021-00770-y.
U.S. Environmental Protection Agency [EPA], 2024, Tracking cyanoHABs—Mapping harmful algal blooms reported in U.S. fresh waters: U.S. Environmental Protection Agency Office of Water website, accessed January 19, 2024, at https://storymaps.arcgis.com/stories/d4a87e6cdfd44d6ea7b97477969cb1dd.
U.S. Geological Survey [USGS], 2018, Dissolved oxygen and water: U.S. Geological Survey Water Science School website, accessed July 27, 2023, at https://www.usgs.gov/special-topics/water-science-school/science/dissolved-oxygen-and-water.
U.S. Geological Survey [USGS], 2019, Water data visualizations: USGS Water Resources Mission Area website, accessed August 7, 2023, at https://www.usgs.gov/mission-areas/water-resources/science/water-data-visualizations.
U.S. Geological Survey [USGS], 2020, SPARROW mappers: USGS Water Resources Mission Area website, accessed March 3, 2025, at https://www.usgs.gov/mission-areas/water-resources/science/sparrow-mappers.
U.S. Geological Survey [USGS], [2025], Hydrologic unit codes (HUCs) explained: U.S. Geological Survey website, accessed March 3, 2025, at https://www.usgs.gov/mission-areas/water-resources/science/sparrow-mappers.
Van Metre, P.C., Waite, I.R., Qi, S., Mahler, B., Terando, A., Wieczorek, M., Meador, M., Bradley, P., Journey, C., Schmidt, T., and Carlisle, D., 2019, Projected urban growth in the southeastern USA puts small streams at risk: PLoS ONE, v. 14, no. 10, article e0222714, accessed September 18, 2023, at https://doi.org/10.1371/journal.pone.0222714.
Walsh, J., Wuebbles, D., Hayhoe, K., Kossin, J., Kunkel, K., Stephens, G., Thorne, P., Vose, R., Wehner, M., Willis, J., Anderson, D., Doney, S., Feely, R., Hennon, P., Kharin, V., Knutson, T., Landerer, F., Lenton, T., Kennedy, J., and Somerville, R., 2014, Our changing climate, chap. 2 of Melillo, J.M., Richmond, T.C., and Yohe, G.W., eds., Climate change impacts in the United States—The third national climate assessment: U.S. Global Change Research Program, p. 19–67, accessed September 18, 2023, at https://doi.org/10.7930/J0KW5CXT.
Wells, M.L., Karlson, B., Wulff, A., Kudela, R., Trick, C., Asnaghi, V., Berdalet, E., Cochlan, W., Davidson, K., De Rijcke, M., Dutkiewicz, S., Hallegraeff, G., Flynn, K.J., Legrand, C., Paerl, H., Silke, J., Suikkanen, S., Thompson, P., and Trainer, V.L., 2020, Future HAB science—Directions and challenges in a changing climate: Harmful Algae, v. 91, article 101632, accessed May 28, 2024, at https://doi.org/10.1016/j.hal.2019.101632.
Wells, M.L., Trainer, V.L., Smayda, T.J., Karlson, B.S.O., Trick, C.G., Kudela, R.M., Ishikawa, A., Bernard, S., Wulff, A., Anderson, D.M., and Cochlan, W.P., 2015, Harmful algal blooms and climate change—Learning from the past and present to forecast the future: Harmful Algae, v. 49, p. 68–93, accessed May 28, 2024, at https://doi.org/10.1016/j.hal.2015.07.009.
Wheeling, K., 2019, Toxic algal blooms are worsening with climate change: Eos (Washington, D.C.), v. 100, accessed November 15, 2019, at https://doi.org/10.1029/2019EO136398.
Wilhelm, S.W., Bullerjahn, G.S., and McKay, R.M.L., 2020, The complicated and confusing ecology of Microcystis blooms: mBio, v. 11, no. 3, accessed March 17, 2022, at https://doi.org/10.1128/mBio.00529-20.
Witze, A., 2021, Racism is magnifying the deadly impact of rising city heat: Nature, v. 595, no. 7867, p. 349–351, accessed June 9, 2021, at https://doi.org/10.1038/d41586-021-01881-4.
Wood, R.R., Lehner, F., Pendergrass, A.G., and Schlunegger, S., 2021, Changes in precipitation variability across time scales in multiple global climate model large ensembles: Environmental Research Letters, v. 16, no. 8, article 084022, accessed July 27, 2023, at https://doi.org/10.1088/1748-9326/ac10dd.
Woolway, R.I., Sharma, S., Weyhenmeyer, G.A., Debolskiy, A., Golub, M., Mercado-Bettín, D., Perroud, M., Stepanenko, V., Tan, Z., Grant, L., Ladwig, R., Mesman, J., Moore, T.N., Shatwell, T., Vanderkelen, I., Austin, J.A., DeGasperi, C.L., Dokulil, M., La Fuente, S., Mackay, E.B., Schladow, S.G., Watanabe, S., Marcé, R., Pierson, D.C., Thiery, W., and Jennings, E., 2021, Phenological shifts in lake stratification under climate change: Nature Communications, v. 12, no. 1, article 2318, accessed July 28, 2023, at https://doi.org/10.1038/s41467-021-22657-4.
Wu, Y., Wen, Y., Zhou, J., and Wu, Y., 2014, Phosphorus release from lake sediments—Effects of pH, temperature and dissolved oxygen: KSCE Journal of Civil Engineering, v. 18, p. 323–329, accessed October 16, 2018, at https://doi.org/10.1007/s12205-014-0192-0.
Xue, Z., Zhu, W., Zhu, Y., Fan, X., Chen, H., and Feng, G., 2022, Influence of wind and light on the floating and sinking process of Microcystis: Scientific Reports, v. 12, no. 1, article 5655, accessed July 18, 2023, at https://doi.org/10.1038/s41598-022-08977-5.
Zhai, R., Tao, F., Lall, U., Fu, B., Elliott, J., and Jägermeyr, J., 2020, Larger drought and flood hazards and adverse impacts on population and economic productivity under 2.0 than 1.5°C warming: Earth's Future, v. 8, no. 7, accessed September 18, 2023, at https://doi.org/10.1029/2019EF001398.
Conversion Factors
Temperature in degrees Celsius (°C) may be converted to degrees Fahrenheit (°F) as follows:
°F = (1.8 × °C) + 32.
Temperature in degrees Fahrenheit (°F) may be converted to degrees Celsius (°C) as follows:
°C = (°F – 32) / 1.8.
Abbreviations
CDC
Centers for Disease Control and Prevention
CO2
carbon dioxide
DO
dissolved oxygen
EPA
U.S. Environmental Protection Agency
GCM
global climate model
HAB
harmful algal bloom
HUC
Hydrologic Unit Code
ITRC
Interstate Technology and Regulatory Council
N
nitrogen
N2
Non-reactive nitrogen gas
P
phosphorus
PRMS
Precipitation-Runoff Modeling System
SPARROW
Spatially Referenced Regression on Watershed attributes
SPEI
Standardized Precipitation Evapotranspiration Index
SWAT
Soil and Water Assessment Tool
USGS
U.S. Geological Survey
WASP
Water Quality Analysis Simulation Program
For more information about this publication, contact
Director, Lower Mississippi-Gulf Water Science Center
U.S. Geological Survey
640 Grassmere Park, Suite 100
Nashville, TN 37211
For additional information, visit
https://www.usgs.gov/centers/lmg-water/
Publishing support provided by
Lafayette Publishing Service Center
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
Byl, T.D., Moore, D.M., Cunningham, C., and Young, D., 2025, Science needs for determining the effects of climate change on harmful algal blooms in the southeastern United States: U.S. Geological Survey Open-File Report 2025–1004, 29 p., https://doi.org/10.3133/ofr20251004.
ISSN: 2331-1258 (online)
| Publication type | Report |
|---|---|
| Publication Subtype | USGS Numbered Series |
| Title | Science needs for determining the effects of climate change on harmful algal blooms in the southeastern United States |
| Series title | Open-File Report |
| Series number | 2025-1004 |
| DOI | 10.3133/ofr20251004 |
| Publication Date | March 20, 2025 |
| Year Published | 2025 |
| Language | English |
| Publisher | U.S. Geological Survey |
| Publisher location | Reston, VA |
| Contributing office(s) | Lower Mississippi-Gulf Water Science Center |
| Description | vii, 29 p. |
| Online Only (Y/N) | Y |