

Climate Change Impacts on Plant Communities in the Sagebrush Region—A Science Synthesis to Inform Bureau of Land Management Resource Management
Links
- Document: Report (54.0 MB pdf) , HTML , XML
- Download citation as: RIS | Dublin Core
Acknowledgments
This synthesis benefited from valuable early input about structure and content provided by several Bureau of Land Management (BLM) staff, including Pat Deibert, Chris Domschke, Fred Edwards, Nikki Grant-Hoffman, Doug Havlina, Dominika Lepak, Don Major, Meg McLachlan, James Miller, David Mueller, Karen Prentice, Emmet Pruss, Justin Shirley, Erik Vernon, Aaron Wilkerson, and David Wood as well as Lindy Garner from the U.S. Fish and Wildlife Service. We are especially grateful to Aaron Wilkerson (BLM), Sarah Carter (U.S. Geological Survey [USGS]), Maddy Case (USGS), and Peter Adler (Utah State University) for detailed comments on the full synthesis. This synthesis project was supported in part by the USGS Ecosystems Mission Area Land Management Research Program, the USGS Northwest Climate Adaptation Science Center, and the Yale University School of the Environment. Land Management Research Program scientists work with resource managers to provide science and data products to help Federal, State, and local agencies and Native American Tribes conserve, manage, and restore our Nation’s diverse socioecological systems, often focused on management of public lands and resources at landscape scales. The Yale University School of the Environment is leading the world toward a sustainable future with cutting-edge research, teaching, and public engagement on society’s evolving and urgent environmental challenges.
Executive Summary
Background: Artemisia tridentata Nutt. (big sagebrush) plant communities once covered 62 million hectares of North America across the Western United States and Canada. Since the mid-19th century, the distribution of native intact big sagebrush plant communities has been reduced by nearly 50 percent threatening the habitat of greater than (>) 350 plant and animal species of conservation concern. Some of these species are under consideration for threatened or endangered classifications under the Endangered Species Act (16 U.S.C. 1531 et seq.), notably Centrocercus urophasianus (greater sage grouse) and Brachylagus idahoensis (pygmy rabbits). The abundance of intact sagebrush plant communities has diminished substantially since approximately 1970 in response to a variety of threats, primarily wildfire and invasive species. As of 2024, big sagebrush vegetation in the United States is largely on public land managed by Federal agencies. These agencies, especially the Bureau of Land Management (BLM) and the U.S. Department of Agriculture Forest Service, are tasked with overseeing multiple uses and activities on native sagebrush habitat. These uses and activities include extensive low-impact uses, such as livestock grazing, and localized high-impact uses, such as energy development and mining. Parts of the sagebrush region are being converted to annual grassland or to conifer woodland. In addition to the existing challenges of wildfire, invasive species, and land-use pressures, Federal agencies must now also consider the present and potential future impacts of climate change on sagebrush vegetation.
How this report can inform natural resource planning and prioritization: This synthesis is designed to support resource management decisions related to conservation and restoration of sagebrush ecosystems in the context of climate change. Contents of this report include a brief overview of climate projections for the sagebrush region, synthesized information about potential effects of climate change on sagebrush plant communities, an overview of potential implications of these climate impacts on current and potential future land uses across the region, and information about additional resources that may be helpful for specific climate-related decisions (fig. ES1).

Figure ES1. Conceptual diagram describing the synthesis structure of climate change impacts on sagebrush plant communities in the sagebrush region. The synthesis summarizes potential impacts on individual plant species, plant functional types, and big sagebrush plant community responses. Scientific information is partitioned based on the data source type (field studies or modeling) to highlight each methods advantages and disadvantages. The synthesis then reviews potential implications for decisions and land management and identifies additional resources for evaluating climate change impacts. CO2, carbon dioxide; C4, warm season; C3, cool season
Climate change may enhance the potential for annual grass invasion into sagebrush plant communities. Annual grass invasion is promoted by wildfire but does not depend entirely on wildfire. Invasion by nonnative annual grasses and other climate-mediated changes could, in turn, affect decisions about land uses such as recreation and habitat management. Changes in plant phenology (for example, changes resulting from earlier meteorological springs and later falls) may change patterns of forage availability, potentially affecting migration timing for ungulates, nesting for greater sage grouse, and the optimal timing for livestock grazing. Changes in the recovery potential of sagebrush communities following disturbances such as energy development or wildfire will likely cause increased challenges for restoration, especially as droughts become more frequent and likely more severe. As warmer potential future conditions promote more frequent wildfire in most of the sagebrush region, postfire rehabilitation efforts will become increasingly important for sustaining intact sagebrush plant communities. Finally, greater amounts of bare ground (promoted by hotter conditions with longer dry periods) will increase the potential for erosion, particularly considering the projected increase in heavy rain.
Sagebrush ecosystems are a type of dryland vegetation. Drylands are characterized by high evaporative demand and limited precipitation and are sensitive to shifts in the amounts and timing of precipitation. In these conditions, vegetation growth and reproduction are limited by access to water. This water limitation is well described by the strong effect of precipitation on productivity and species richness in drylands. Shifts in the timing and amount of available water, or the effects of temperature shifts on evaporative demand, will have direct consequences for the composition and extent of sagebrush plant communities.
Current and Projected Climate Futures: Big sagebrush plant communities are widespread across Western North America and exist within a broad gradient of climatic conditions. As of 2024, the sagebrush region is characterized by a mean annual precipitation between 220 and 620 millimeters. Peak water availability occurs during the fall and winter because of low evaporative demand, which allows water to percolate to deep soil layers where it is accessible to big sagebrush plant communities during the warmer growing season. As precipitation and temperature shift, the variability in the climate is expected to interact with these changes to differentially affect big sagebrush plant communities across the region.
Although there is variability in the projections of global climate models, most models agree that temperature in the sagebrush region will increase by >1.5 degree Celsius (°C) by the mid-21st century and by >2.5 °C by the end of the 21st century under a moderate climate change scenario (Coupled Model Intercomparison Project Phase 5 [CMIP5], representative concentration pathway [RCP] 4.5; U.S. Global Change Research Program, 2017). Further, there is agreement that the magnitude of variability within and among years in precipitation and temperature will increase.
Compared to projections about temperature, potential future changes in precipitation amounts and seasonality are much less consistent among climate models. Although models generally agree that projected precipitation will increase slightly by the middle and end of the 21st century, this outcome is not consistent among climate models. Seasonality of precipitation is projected to shift toward winter and fall according to many models, but this outcome is variable, and in some places summer precipitation is expected to increase. The lack of consistent predictions about the magnitude and fine-scale spatial distribution of precipitation changes in regions, such as the Southwest, may be attributed to unpredictable precipitation patterns with low overall annual rainfall.
Potential Climate Change Impacts on Plant Communities in the Sagebrush Region: The consequences of rising temperatures for plants are expected to vary across the sagebrush region. In areas where the growing season starts later in the spring because of lower temperatures and snowpack, temperature increases will likely have a positive effect on plant growth in sagebrush plant communities. In parts of the sagebrush region characterized by limited (for example, shorter or fragmented) growing seasons because of high summer temperatures, such as low-elevation basins and the southern extent of the region, increased temperatures will limit the growing season. This effect may especially be true for cool-season (C3) perennial grasses, which may be replaced by warm-season (C4) perennial grasses as climatic suitability for warm-season grasses increases.
Although there is high uncertainty in projected changes to total precipitation across the region, the most robust signal is an increase in precipitation in the north, leading to a potential increase in plant productivity and increased suitability for native shrubs, perennial grasses, and forbs. In the southern part of the region, decreases in total precipitation will likely reduce productivity and cause declines for perennial, herbaceous species and lead to an increase in bare ground. Shifts in precipitation may also decrease the suitability of Artemisia tridentata ssp. vaseyana (Rydb.) Beetle (mountain big sagebrush), favoring the basin adapted subspecies Artemisia tridentata ssp. wyomingensis Beetle & Young (Wyoming big sagebrush) and Artemisia tridentata ssp. tridentata Nutt. (basin big sagebrush). Typically, such shifts are correlated with lower productivity and biodiversity associated with decreased water availability. Increases in fall and winter precipitation will favor the persistence and success of big sagebrush in the northern part of the region. This trend does not extend across the entire region, and there is less agreement among climate models in the southern parts of the region. Increases in the variability of precipitation are likely to lower the total abundance of herbaceous species and increase variability in productivity.
An increased frequency of extreme climate events including high-temperature drought and increased prevalence of extreme precipitation events are projected to have immediate effects on sagebrush plant communities. Additionally, multiyear droughts are projected to become more frequent across the Western United States. The primary threat of these conditions is the loss of understory cover and biodiversity, which can lead to more bare ground, lower productivity, and a higher susceptibility to invasion by nonnative annual grasses. Some high temperature and multiyear drought conditions may cause big sagebrush mortality. Mortality can also be attributed to heavy rainfall events that lead to saturated soil and anoxic (low oxygen) root conditions and an increase in the risk of erosion.
Shifts in climate will also interact with disturbance regimes. Increasing drought risk and interannual variability in precipitation can increase fine fuel loads and increase the severity of wildfires, particularly in areas already dominated by Bromus tectorum L. (cheatgrass). Warming temperatures may also increase the probability of nonnative grass invasion, which would expand their distribution and areal cover. Such expansion has been observed in some parts of the Great Basin. Even when cheatgrass areal cover is low, cheatgrass is linked to greater fire probabilities that further increase the risk of shorter fire return intervals that promote greater cheatgrass invasion.
Implications of Potential Vegetation Changes for Management Decisions: The projected shifts in sagebrush communities will have widespread implications for management, land use, and restoration and rehabilitation efforts across the region. Although the exact nature of big sagebrush plant community response to climate change will vary depending on location, a few general trends can be expected. The northern extent of the range and higher-elevation sites are expected to remain stable and potentially expand in sagebrush community suitability. Meanwhile, the southern extent of the range and low-elevation sites in warm intermountain basins are projected to experience decreases in the ecohydrological growing season (warm conditions with sufficiently wet soil to support plant growth) and total water availability. These changes are likely to reduce overall productivity and stability of sagebrush vegetation.
More variability in temperature and precipitation and an increased likelihood of extreme weather events will further alter these communities. Greater interannual variability and precipitation may promote the expansion of conifers into sagebrush region, but overall conifer expansion may become less prevalent. More variability in climate may have the largest negative impact on sagebrush community productivity. Many livestock and wildlife species are dependent on these communities for forage and habitat, including greater sage grouse. More variability in climate will likely cause widespread plant mortality, lower forage availability, and increase bare ground. Shifts in the growing season length will alter the timing of plant productivity and the ability to recover from disturbances. Many of the projected changes will increase the climatic suitability for warm-season perennial and annual grasses and effect the timing of wildlife migrations and livestock grazing.
Consequently, climate change will likely necessitate a re-evaluation of decisions about livestock grazing intensity, frequency, season, and duration and appropriate target population levels for feral horses and burros and wild ungulates. Changes in precipitation amounts and timing will affect plant phenology and forage production. Plant phenology will also be affected by changes in temperature, which will affect the start of the livestock grazing season. The amount of forage will be most heavily affected by precipitation, which will in turn affect stocking rates and the duration of the livestock grazing season.
Climate change is also expected to impact fuel management. Warming and drying will lead to earlier and longer wildfire seasons, increased probability of wildfire, and larger wildfire size. Drought conditions characterized by high temperatures also increase the probability of wildfire. Enhanced variability in precipitation will alter fuel dynamics such that years with high productivity may be followed by dry periods and may lead to increased fuel loads with a high probability of ignition.
Recreation and other public uses of Bureau of Land Management-managed sagebrush will also be affected by climate change. The effects will include an increased threat of erosion because of increased bare ground, frequency of extreme precipitation and temperature events, and vulnerability to invasion by nonnative plant species in the sagebrush region, primarily along trails and other heavily used areas. As temperatures rise and droughts increase in frequency, human-caused wildfire ignition will become more prevalent, especially as recreation numbers increase. Finally, seasonal recreation uses in sagebrush landscapes, notably Odocoileus spp. (deer) and Cervus canadensis (elk) hunting, may shift as wildlife migration timing changes in response to altered seasonal forage availability patterns.
Restoration and rehabilitation of native plant communities, which are already challenging in the big sagebrush region, will likely become more difficult as temperatures, precipitation variability, and the frequency of extreme temperature and precipitation events increase. These challenges will affect land treatments designed for postfire restoration and postdevelopment rehabilitation. Treatment strategies may need to be re-evaluated to assure that they will succeed under new conditions.
Purpose of This Report
Artemisia tridentata Nutt. (big sagebrush) ecosystems cover 62 million hectares across 11 Western U.S. States and Canada and are largely managed by Federal agencies (fig. 1; Miller and others, 1994; West and Young, 2000). In accordance with the National Environmental Policy Act (42 U.S.C. 4332(c); NEPA), Federal agencies are required to assess the potential environmental impacts of management actions that may substantially affect the quality of the human environment. In the United States, these agencies are tasked with managing landscapes with sagebrush plant communities for multiple resources, uses, and values. Specific management actions in the sagebrush region include permitting land uses like livestock grazing; development of oil, gas, and renewable energy resources; hard rock mining; and recreation and implementing land treatments to promote ecological restoration and rehabilitation of degraded areas.
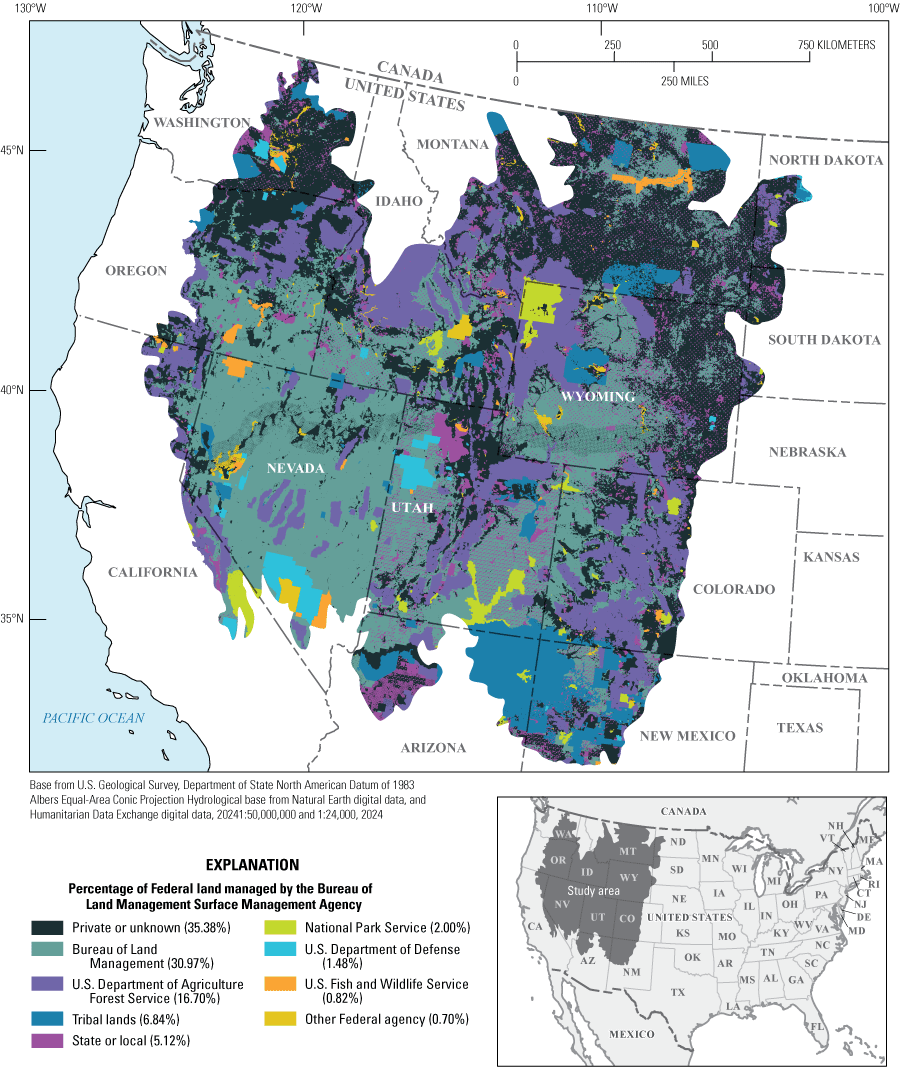
Map showing the big sagebrush region in the Western United States and percentage of land that is Federal, State, Tribal, or privately owned or managed. The area percentages by the Bureau of Land Management Surface Management Agency (SMA) dataset sagebrush biome polygon from Jeffries and Finn (2019). %, percent
A changing climate is expected to alter ecosystems, plant communities, and wildlife populations in the sagebrush region and thus may affect specific land management decisions (Stein and others, 2014). Recognizing potential climate change impacts on sagebrush plant communities is a first step toward understanding how land management in the sagebrush region may need to change to ensure continued compliance with the NEPA, Executive Orders related to climate adaptation in natural resource management (for example, Executive Order 13990) and U.S. Department of the Interior policies (for example, U.S. Department of the Interior, 2008, 2023192)
The goal of this report is to synthesize available information about how a changing climate, particularly elevated atmospheric carbon dioxide (CO2) concentrations, rising temperature, altered precipitation patterns, enhanced variability, and increasing frequency of extreme weather events, will impact plant community dynamics across the sagebrush region of the Western United States (fig. ES1). This report reviews potential effects on individual species, plant functional types, and expected consequences for whole plant communities in different areas of the sagebrush region. This report also considers the potential implications of climate impacts on sagebrush plant communities for several types of permitted uses and activities on Bureau of Land Management (BLM)-managed sagebrush habitats.
How to Use This Report
This report can be used to help natural resource management staff in the sagebrush region understand the potential effects of climate change on plant communities and related management decisions. It may be useful when evaluating the vulnerability of species and ecosystem processes and when making determinations about the potential effects of management alternatives. It may also aid in the development of environmental decision documents, such as environmental assessments (NEPA). This report may also be useful for the development of internal policy or guidance regarding land treatments in the sagebrush region. This report can also serve as a reference, supplementary information, or as a synthesis of currently (2024) available, peer-reviewed literature related to climate change in the sagebrush region. Please cite this report as Carpenter and others (2025) when appropriate.
For NEPA analyses, this synthesis can function in two main ways. First, this synthesis can serve as a foundation for recognizing the potential effects of climate change on sagebrush plant communities to assess the future potential resistance and resilience in response to specific uses (40 CFR §1502.15). Second, this synthesis considers the implications of these climate impacts for specific land uses and may inform how land management decisions can affect sagebrush vegetation in conjunction with climate change (40 CFR §1502.14). Beyond NEPA analyses, this synthesis can inform considerations of how different areas of the sagebrush region may respond to ecological restoration and rehabilitation treatments (notably postfire seeding treatment). This synthesis may also help identify areas of high conservation priority in conjunction with ecological site descriptions (Doherty and others, 2022) and ongoing assessment, inventory, and monitoring efforts (for example, the BLM Assessment, Inventory and Monitoring [AIM] Strategy; https://www.blm.gov/aim). This report can serve as a reference for understanding the response of sagebrush plant communities to climate in different geographic regions as climate continues to change throughout the 21st century. Appendix 1 provides maps of projected climate conditions showing mean annual temperature (fig. 1.1A–I), mean annual precipitation (fig. 1.2A–I), standard deviation of annual temperature (fig. 1.3A–I), coefficient of variation of annual precipitation (fig. 1.4A–I), mean day of year with the last spring frost (fig 1.5A–I), mean day of year with the first fall frost (fig. 1.6A–I), mean growing season length (fig. 1.7A–I), and mean seasonal timing of precipitation (fig. 1.8A–I).
Synthesis Approach and Objectives
This synthesis offers a perspective on the multifaceted impacts of climate change on big sagebrush plant communities. Two aspects of changing climate conditions were considered—shifting averages and extreme weather events—and results were synthesized from observational studies, manipulative experiments, and modeling. Each of these types of studies has its own set of strengths and weaknesses, but this report highlights the salient implications for management.
Section 1.2. “Considerations for Assessing Climate Effects—Changing Mean Conditions and Enhanced Variability” presents an overview of the projected changes in climate (fig. 2; table 1) under two representative concentration pathways (RCPs), RCP4.5 and RCP8.5 from the 5th iteration of the Intergovernmental Panel on Climate Change (IPCC) coupled model intercomparison project (CMIP5; IPCC, 2014). These two pathways represent a moderate greenhouse gas (GHG) emission scenario (RCP4.5) and a higher emission scenario (RCP8.5). Both scenarios provide a wide range of possible climate futures that enables a robust understanding of potential effects. These pathways are possible futures that do not represent predictions but show the consequences of what happens if humanity were to emit GHGs as described by RCP4.5 or RCP8.5.
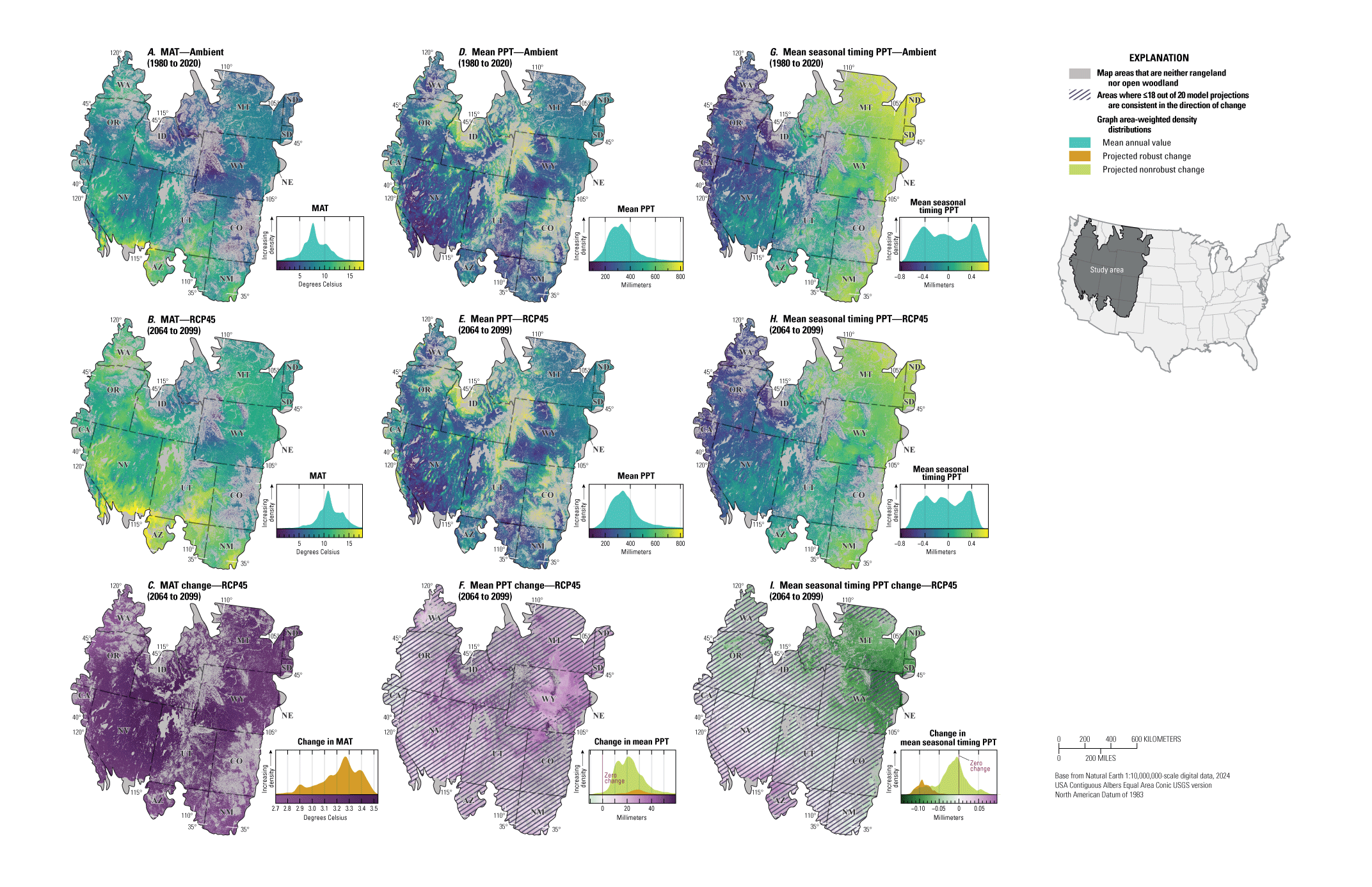
Maps showing the ambient, projected future, and projected changes in A–C, mean annual temperature (MAT; in degrees Celsius), D–F, mean annual precipitation (PPT, in millimeters), and G–I, mean precipitation seasonal timing for the sagebrush region. Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Ambient conditions include the years from 1980 to 2020 and future projections show median values across 20 global circulation models (GCMs) from the Coupled Model Intercomparison Project Phase 5 (CMIP5) representative concentration pathway (RCP) 4.5 during the years from 2064 to 2099, and the median difference (change) between future and historical projections. Cross-hatching of projected changes shows areas where less than or equal to (≤) 18 of 20 of GCM projections are consistent in the direction of change (nonrobust). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019).
Table 1.
Projected changes in climate and drought for the Artemisia tridentata Nutt. (big sagebrush) region.[Refer to appendix 1 for related maps. Confidence is expressed as high (H), medium (M), or low (L). Wet degree days (WDD) refers to plant growth potential quantified by growing degree days when soils are wet. CO2, carbon dioxide; CWD, climatic water deficit]
The CMIP5 is not the most recent climate change model comparison version (CMIP6), but it remains the dominant source of climate change projections in ecological research because of the wide availability of downscaled products (O’Neill and others, 2016). The general trends projected in CMIP5 are consistent with projections from CMIP6 (Thorarinsdottir and others, 2020; Martel and others, 2022). There are also a wide range of projections from the different global climate models (GCMs) within CMIP5, which result in variability of predicted responses. The goal of this report is to assess the expected impacts of climate change under both RCPs based on the median response of multiple GCMs, where possible.
This report assesses the consequences of projected changes in climate for big sagebrush plant communities, ranging from individual plants to entire landscapes, by reviewing the impacts of climate change on key plant functional types. Understanding how different functional types respond to changing temperature, precipitation, and CO2 is the first step in understanding the overall resilience and adaptability of big sagebrush communities. Then habitat modeling efforts that link predicted changes in climatic variables to projected distributions of plant species and functional types were reviewed. Shifts in species distributions have implications for understanding potential alterations in community composition and structure.
Section 1.3. “Climate Effects on Individual Plant Functional Types” synthesizes information and focuses on the interactions and feedback that may occur within the community. By considering the responses of multiple species, a better understanding of how climate change may reshape the functioning and stability of sagebrush ecosystems can be developed. Finally, the report examines how process-level modeling has described the impact of climate change on big sagebrush communities. Modeling results help to assess the effects of climate change on big sagebrush ecosystems, including potential changes in species distributions, community composition, and landscape-level dynamics. Additionally, exploring different modeling approaches helps to identify uncertainties and gaps in the understanding of the interactions between climate and sagebrush plant communities.
Caveats to Use of This Report
This synthesis is not a complete source for NEPA analyses of the effects of alternative management actions. More specific and detailed information will be helpful to understand potential climate changes impacts in local areas or for individual species or plant communities, and the references identified here may provide a first step. This report does not address the effects of climate change for wildlife other than through the potential implications of shifts in the quality and quantity of forage and habitat. This report does not address changing multitrophic interactions that could affect sagebrush communities, such as the effects of changing arthropod or vertebrate population dynamics. It also does not address literature regarding the effects of climate change for biocrusts or carbon and nutrient cycling.
Science Synthesis—Climate Change Impacts on Sagebrush Plant Communities
1. Characterizing Climate Change Impacts on Big Sagebrush Ecosystems
Section 1 Highlights
● Big sagebrush plant communities are widespread across the Western United States and are in areas with cold winters and relatively dry summers.
● Throughout the remainder of the 21st century, increasing temperatures across the region are projected by all climate models.
● Projected precipitation changes are variable, although precipitation may increase slightly during the cool season, particularly in the northeastern part of the region.
1.1. Basics About Big Sagebrush Ecosystems
Big sagebrush plant communities are characterized by limited water availability (fig. 2), temperate climates with cold winters, and primarily cool-season precipitation, which promotes the dominance of woody species (Schlaepfer and others, 2012a). Understanding how climate impacts big sagebrush plant communities is necessary to assess changes in vegetation (Maestre, Salguero-Gómez, and Quero, 2012; Huang and others, 2016). One of the main factors affecting dryland vegetation is moisture availability, and annual precipitation is often a key determinant of primary productivity (McNaughton and others, 1989; Hsu and others, 2012). In addition to total precipitation, the seasonal timing and the size distribution of precipitation events (many small compared to few large occurrences) exert significant effects on productivity, vegetation structure, and species richness (Bates and others, 2006; Felton and others, 2021; Hou and others, 2021). This effect is further evidenced by the control seasonality has on dryland ecosystems, which dictates whether woody or herbaceous species dominate the landscape (Wiegand and others, 2006; Renne, Bradford, and others, 2019; Holdrege and others, 2021).
The dominance of woody plants is promoted by relatively greater water available in deep soil layers compared to shallow soil layers—a condition promoted in areas that receive a large proportion of precipitation during the cool season (Bates and others, 2006; Pennington and others, 2019). As water reaches deep soil strata, it provides a favorable environment for woody roots to outcompete most herbaceous species (Walter, 1973; Sala and Lauenroth, 1982). In big sagebrush ecosystems, the productivity and richness of the herbaceous community are contingent on the magnitude of spring precipitation (Liang and others, 2021), which emphasizes the critical role of water availability in shaping vegetation.
1.2. Considerations for Assessing Climate Effects—Changing Mean Conditions and Enhanced Variability
Considering the importance of climatic variables in shaping sagebrush vegetation, climate change has the potential to alter sagebrush plant communities and impact their sensitivity to threats like wildfires and biological invasions (Coates and others, 2016; Adler and others, 2018; Zimmer and others, 2021). This section focuses on synthesizing information about the potential effects of altered precipitation patterns, increasing temperature, elevated CO2, and increased interannual precipitation variability on sagebrush plant communities. This section also distinguishes among the effects of shifting average conditions, increased interannual variability, and frequency of extreme weather events. This categorization allows for a better understanding of the complexity of climate change effects on big sagebrush ecosystems.
Shifting averages refers to long-term trends in average temperature, precipitation, and atmospheric CO2 and other GHG emissions. Although all GHGs affect the general warming trend, this section focuses on CO2 because of its direct effect on photosynthesis and its potential to affect vegetation dynamics (Ainsworth and Long, 2005). Evidence for rising temperature and CO2 is unambiguous (IPCC, 2022), and both will affect plant communities. Annual and seasonal temperature increases are important determinants of soil water availability and consequently shape the effects of temperature increases on plant communities (Kleinhesselink and Adler, 2018). As temperatures rise, the distribution and phenology of specific grass or forb species may be altered, which affects the overall structure and functioning of sagebrush ecosystems (Perfors and others, 2003; Roundy and others, 2018; Bloom and others, 2022). Plant water-use efficiency may be increased by rising atmospheric CO2 concentrations, potentially improving plant response to drought and increased temperatures (Hamerlynck and others, 2000; Lee and others, 2011). However, the effects of CO2 are less clear across long treatment periods (Ainsworth and Long, 2005; Morgan and others, 2011).
Observations and potential future expectations for long-term precipitation trends are variable across the sagebrush region (U.S. Global Change Research Program, 2017; Zhang and others, 2021). This variability will lead to differential responses among plant species and communities and make forecasting and adaptation strategies at the local scale complex. Regardless of the directionality, changes in annual precipitation will affect soil water availability, which exacerbates drought conditions or introduces novel challenges to the stability of big sagebrush plant communities (Hsu and others, 2012).
Most GCMs project an increase in the frequency of extreme weather events, which can have lasting impacts on ecological functioning (Smith, 2011; IPCC, 2022). The consequences of increased climatic variability make long-term planning and management decisions uncertain (Hou and others, 2021). In the sagebrush region, extreme weather events are likely to manifest as high-temperature droughts, multiyear droughts, and heavy precipitation that lead to erosion or anoxic (low oxygen) root conditions (U.S. Global Change Research Program, 2017). Although historical droughts have affected the region for millennia, the magnitude and severity of potential future heat, drought, and precipitation events are expected to affect abiotic conditions and community interactions not within the observed range of variability. Such events can affect the structure, composition, and stability of sagebrush plant communities (Maxwell and others, 2019; Renne, Schlaepfer, and others, 2019).
1.2.1. Regional Climate Projections and Uncertainties for the Sagebrush Region
Compared to temperatures from the baseline period from 1980 to 2010, predicted future temperatures for the sagebrush region include a median annual increase under representative concentration pathway (RCP) 4.5 of 1.5 °C by 2050, and 2.0 °C by 2100 (Palmquist and others, 2021). Under RCP8.5, the projected increases in temperature are more than double those increases expected under RCP4.5, which emphasizes the importance of considering various emission scenarios when assessing climate impacts (U.S. Global Change Research Program, 2017).
Regional precipitation projections are more complex than those projections for temperature. Under RCP4.5, the median change in annual precipitation is relatively modest and has an increase of less than 20 millimeters (U.S. Global Change Research Program, 2017). However, some parts of the sagebrush range are projected to experience substantial increases in precipitation and exceed 100 millimeters by the end of the century (Bradford and others, 2020; U.S. Global Change Research Program, 2017). Other parts, especially in the southern part of the region, may have a decline in precipitation that further decreases soil water availability; when coupled with increases in temperature, this decline will decrease habitat suitability for sagebrush (Palmquist and others, 2021; Doherty and others, 2022).
Rising temperatures and shifting precipitation will alter seasonal patterns of soil moisture availability (Bradford and others, 2020). Interannual variability and the frequency of extreme weather events represent large sources of uncertainty among GCMs. Although there is no consensus across GCMs with respect to changes in precipitation, most agree that the intensity and frequency of extreme weather events will increase with rising temperatures (Breshears and others, 2016; Armal and others, 2018). Under RCP8.5, the impact of extreme weather events will be more severe than RCP4.5 (U.S. Global Change Research Program, 2017).
All climate projections are made with some degree of uncertainty, and the projected changes differ across emissions scenarios further amplify that uncertainty. However, managing natural resources has always involved uncertainty about fluctuations in climate and biotic populations. The uncertainty about changes in precipitation and temperatures and enhanced variability presents additional challenges. Resource management professionals may want to focus on conserving existing diversity and ecosystem functionality while being aware of the degree of uncertainty and planning for timelines consistent with expected responses.
1.3. Climate Effects on Individual Plant Functional Types
Section 1.3 Highlights
● The following are main plant functional types in big sagebrush plant communities: the three subspecies of big sagebrush, other shrubs, perennial grasses and forbs, annual grasses and forbs, and invasive, cool-season annual grasses.
● The effects of climate change on plants in the sagebrush region will vary among and within plant functional types. However, it is likely that shrubs will benefit from increased cool-season precipitation and are likely to remain climatically viable across much of the region.
● Effects of climate change on plants will also vary across the climatic gradients within the sagebrush region. Plants in the southern part of the region may be more sensitive to increasing temperatures that may reduce abundance of perennial herbaceous species.
● Adverse effects of climate change on plants in the sagebrush region may be exacerbated by increasing disturbance frequency, including droughts and wildfires.
● Adverse effects of climate change on plants in the sagebrush region may also be moderated by higher plant water use efficiency driven by rising CO2 concentrations.
Climate change will initially affect individual plants, and their growth and reproductive success, within the sagebrush region. Although these effects vary among plant species, they will align with well-defined patterns based on plant functional types (Díaz and Cabido, 1997). The effects of climate change on the following plant function types were considered: big sagebrush, other shrubs, perennial bunchgrasses, and perennial forbs. This characterization follows the functional types for dryland plant communities described by Sala and others (1997). A separate discussion to the effects of climate change on nonnative species, such as Bromus tectorum L. (cheatgrass), Taeniatherum caput-medusae (L.) Nevski (medusahead), and Alyssum desertorum Staph (desert madwort), examines the effects of warming, precipitation changes, elevated atmospheric CO2 concentration, and interannual variability on each plant functional type.
1.3.1. Big Sagebrush
Big sagebrush is the dominant species in big sagebrush plant communities and is especially abundant in sagebrush ecosystems. Limited research has been done on the response of big sagebrush to CO2 enrichment. Big sagebrush may improve water-use efficiency when CO2 levels are temporarily elevated (Lucash and others, 2005); however, the implications of prolonged CO2 enrichment on long-term big sagebrush growth and survival and ecosystem dynamics are unknown. The long-term effects of elevated CO2 concentrations on big sagebrush may be restricted as plant growth becomes limited by other resources (for example, water and nutrients) after plants have acclimated to elevated CO2 conditions. This limitation by other resources has been observed in other ecosystems (Idso and Idso, 2001; Ainsworth and Long, 2005).
Experimental temperature manipulations on big sagebrush have shown different results depending on ambient climate conditions. In high elevations and in cool sites, experimental warming tends to increase aboveground biomass and growth rate of big sagebrush (Harte and Shaw, 1995; Perfors and others, 2003). However, some studies suggest that temperature increases do not necessarily improve sagebrush photosynthetic performance (Loik and Harte, 1996). In contrast, at low-elevation sites, warming has had a negative effect by reducing the reproductive capacity of big sagebrush (Karban and Pezzola, 2017). This decrease in reproductive success can limit seedling recruitment and, ultimately, its survivability in these areas.
Precipitation has a large effect on big sagebrush performance (table 2). Sagebrush may benefit from increasing cool-season precipitation. Bates and others (2006) found that fall precipitation had a significant positive effect on big sagebrush growth. Furthermore, large precipitation events have been associated with increased sagebrush growth (Holdrege and others, 2021). Precipitation reduction experiments have shown declines in sagebrush growth and reproductive capacity after multiple years of treatment. Seedling establishment experiments with reduced precipitation found that seedlings are less likely to sprout and survive past a year or two (Booth and Bai, 2000; Shriver and others, 2018; Schlaepfer and others, 2021).
Table 2.
Expected effects of different climate change impacts on specific plant functional group in Artemisia tridentata Nutt. (big sagebrush) plant communities.[CO2, carbon dioxide; C3, cool season; C4, warm season]
| Changing condition | Expected impacts | Citations |
|---|---|---|
| Elevated CO2 | Improved water-use efficiency that potentially mitigates drought threat. | Lucash and others (2005) |
| Elevated temperature | Increased growth rate and aboveground biomass in high-elevation sites, increased mortality at low elevation and warm sites. | Harte and Shaw (1995), Perfors and others (2003), Karban and Pezzola (2017) |
| Changing precipitation | Increases in fall and winter precipitation are likely to increase sagebrush growth. The increased size of precipitation events are also expected to increase sagebrush growth. Seedlings at the hot and dry south edge of big sagebrush’s distribution are less likely to establish because of decreased overall precipitation. | Bates and others (2006), Holdrege and others (2021), Shriver and others (2018) |
| Enhanced variability | Expected shifts in seasonality toward increased proportion of precipitation in the fall and winter is expected to benefit sagebrush growth. Shifts toward summer precipitation at the south edge1 are likely to decrease sagebrush regeneration and limit growth. | Bates and others (2006), Schlaepfer and others (2021) |
| Elevated CO2 | Likely divergent, species-specific responses. Some expectations are elevated CO2 will improve water-use efficiency and some drought resistance. | Hamerlynck and others (2000) |
| Elevated CO2 | Improved water-use efficiency but may not improve growth during droughts. | Morgan and others (2011), Lee and others (2011), Ainsworth and Long (2005) |
| Elevated temperature | At warmer sites, species may experience earlier senescence and shorter growing seasons. At high-elevation sites, warming is likely to have a small impact. | Williams and others (2007), Munson and others (2011) |
| Changing precipitation | Dryland perennial grasses tend to be resistant to short-term precipitation reduction treatments, although extended drought may induce perennial grass mortality events. Decreases in overall precipitation are expected to decrease abundance in drier parts of the region. | Adler and others (2009), Winkler and others (2019), Shi and others (2020) |
| Enhanced variability | Increases in winter precipitation particularly under warmer temperatures may increase perennial grass productivity. | Bates and others (2006) |
| Elevated CO2 | Improved water-use efficiency that may improve growth during high temperature periods. | Reich and others (2018), Morgan and others (2011) |
| Elevated temperature | Higher temperatures can increase productivity if water is available. Increased temperatures may also extend the growing season for C4 species and delay flowering times. Temperature increases are also projected to increase overall suitability of C4 species across the region. | Du and others (2011), Hartman and Nippert (2013) |
| Changing precipitation | More total precipitation are likely to benefit C4 perennial grasses, particularly increases in spring and summer precipitation. | Munson and others (2013), Munson and Long (2017), Havrilla and others (2023) |
| Enhanced variability | Despite relative resistance to short-term drought and fluctuations in precipitation, increases in precipitation variability are likely to decrease overall productivity and abundance. | Seleiman and others (2021), Gherardi and Sala (2015) |
| Elevated CO2 | Minimal overall impacts dependent onsite location. | Zavaleta and others (2003), Lee and others (2011) |
| Elevated temperature | Earlier flowering times and potential replacement by grass species. Overall responses will be highly variable depending on functional role, although there is a general trend toward species loss as temperature increases. | Bloom and others (2022), Morgan and others (2011), Lee and others (2011), Cross and Harte (2007) |
| Changing precipitation | Variable response based on species and functional role. Projected decreases in perennial forb suitability across the region, and a potential loss of forb species in dry areas where precipitation is reduced. | Hsu and others (2012), Palmquist and others (2021) |
| Elevated CO2 | Potentially minimal effects. | Blumenthal and others (2016) |
| Elevated temperature | Observational studies have shown higher biomass under warmer conditions. Warming may increase suitability in parts of the region, although it may not at the northernmost edge of the region. | Boyte and others (2016), Howell and others (2020), Smull and others (2019) |
| Enhanced variability | Resilience of annual populations is determined by co-occurring factors and will vary based on the extremity and timing of drought and other weather events. One indirect mechanism that may increase annual abundance is the increased frequency of wildfires. | Williamson and others (2020), Coates and others (2016) |
Refer to figure 3 of this report.
Extreme weather events and climatic variability can play a large role in shaping sagebrush plant communities. Interannual variability in precipitation can strongly affect seedling establishment in mature sagebrush stands, and an increase in drought frequency may lead to population declines (Karban and Pezzola, 2017). Karban and Pezzola (2017) also found an increase in flower production during a multiyear drought followed by branch die-off. High-temperature drought conditions can even lead to stand mortality, particularly when extreme droughts are followed by heavy precipitation (Renne, Schlaepfer, and others, 2019). Additionally, the increasing frequency of heavy downpours may cause mortality by inducing anoxic root conditions through flooding (Renne, Schlaepfer, and others, 2019). Seedling recruitment is also affected by current and recent past conditions, primarily temperatures and soil moisture (Bishop and others, 2020).
The response of big sagebrush to climate change is likely multifaceted and contingent on location and climate conditions. Although big sagebrush in some regions may experience benefits from temperature and precipitation shifts, the southern part of its range may have population declines because of reduced water availability. Moreover, increasing variability and extreme weather events are likely to have negative effects on sagebrush.
1.3.2. Other Evergreen (Semideciduous) Shrubs
In addition to big sagebrush, other shrub species, such as Purshia tridentata (Pursh) DC. (antelope bitterbrush) or Grayia spinosa (Hook) Moq. (spiny hopsage) contribute to early successional communities and provide key forage resources for wildlife within sagebrush ecosystems. Nondominant shrubs include many species with different life histories in various climatic and soil conditions within big sagebrush landscapes. Consequently, the response to climate change likely varies among species. This section discusses the results of studies on these nondominant shrub species in big sagebrush communities; however, there is a large knowledge gap in the understanding of how climate impacts these species.
Increased atmospheric CO2 can improve water-use efficiency in other evergreen shrubs, which suggests a potential positive effect on their water conservation abilities (Hamerlynck and others, 2000). Hamerlynck and others (2000) found that elevated CO2 levels may lead to some resistance to high-temperature droughts. This resistance indicates a potential advantage for these shrubs to adapt to changing climate conditions. Knowledge about other shrub responses to temperature changes is also limited. Based on general principles, their responses will vary depending on their location, local climatic conditions, and other species present (Kopp and Cleland, 2015). In high-elevation, cool sites, they may benefit from warming, similar to the observed effects in big sagebrush. Conversely, in low-elevation, hot sites, these species could experience detrimental effects on reproductive capacity and overall performance.
The response of other evergreen shrubs to changes in precipitation remains uncertain. Their responses may differ depending on the specific location and the extent of changes in precipitation patterns. Understanding how they adapt to shifts in water availability will be crucial in predicting their survival and ecological role within sagebrush ecosystems. The impacts of extreme weather events and interannual variability on other evergreen shrubs are also understudied. However, given the importance of water availability and temperature for plant growth and survival, it can be assumed that extreme weather events and fluctuations in precipitation and temperature will likely affect their performance and distribution. Although some studies suggest potential benefits, such as improved water-use efficiency and resistance to high-temperature droughts under elevated CO2 levels (Hamerlynck and others, 2000), further research is needed to fully understand their adaptive strategies in changing climate conditions.
1.3.3. Perennial Bunchgrasses
Grasses are the primary forage resources for livestock, ungulates such as elk, other herbivorous wildlife, and feral horses. Grasses are also the dominant herbaceous plant functional type in most big sagebrush plant communities and vital components of sagebrush ecosystems (Pennington and others, 2019). Understanding the response of bunchgrasses to climate change (table 2) can aid predictions regarding the overall resilience of big sagebrush plant communities.
Elevated CO2 experiments have shown mixed results and outcomes vary depending on the photosynthetic type of grasses. Initially, studies suggested that elevated CO2 levels benefited cool-season (C3) grass species (Ainsworth and Long, 2005; Lee and others, 2011). However, after prolonged exposure to elevated CO2 levels, C3 and warm-season (C4) grasses show similar performance in water-use efficiency and photosynthetic rate (Hunt and others, 1996). In addition, some studies found that sustained exposure to elevated CO2 effectively mitigated drought for C4 grass species (Reich and others, 2018), an effect that was not observed in C3 grass species (Morgan and others, 2011). Overall, CO2 enrichment has a relatively minor role in determining grass responses to climate change compared with changes in temperature and precipitation.
The expected shifts in precipitation seasonality in areas with little change to annual precipitation or projected increases may cause increases in water availability beyond depths that are accessible to most perennial grass species (Palmquist and others, 2016; Jordan and others, 2020). However, the effects may differ in wet areas of the sagebrush region, particularly in high-elevation sites where shorter snowpack durations and increased rainfall can increase water availability for perennial bunchgrasses (Bates and others, 2006). Other studies have found that in the short-term, perennial grasses are relatively resistant to changes in precipitation regime (Adler and others, 2009). It is also possible that the hydraulic redistribution of water by woody species may buffer perennial grass response to increases in deep soil water availability (Richards and Caldwell, 1987; Lee and others, 2018). However, no experimental study has looked at the role of hydraulic redistribution under altered precipitation regimes and warmer temperatures.
Cool-season grasses are generally the dominant functional type across the northern part of the sagebrush region. Warm-season grasses tend to have higher proportional cover in the warmer southern parts of the region, but some grasses are present in pockets across the region (Havrilla and others, 2023). Cool-season species are generally negatively affected by temperature increases, particularly towards the end of their growing season (Williams and others, 2007); however, grasses show resilience to the adverse effects of high temperatures (Munson and others, 2011). Warm-season species benefit from increased temperatures, and elevated temperatures can increase productivity provided that water availability remains sufficient (Du and others, 2011). Temperature changes can also affect flowering time. Increased temperature can advance C3 grass flowering time and potentially delaying C4 species (Hartman and Nippert, 2013).
Perennial bunchgrasses show some degree of acclimation to short periods of high temperature with extreme weather and climate variability. They also show resistance to moderate drought conditions but not to extreme droughts (Seleiman and others, 2021). Elevated CO2 has been found to alter the resilience of grasses to extreme droughts. However, the resilience is species specific and related to how different species store sugars, which suggests that some species will be more adapted to increased interannual variability (Bushey and others, 2023). Additionally, the type of sustained or repeated drought can affect grass responses (Winkler and others, 2019). Increasing variability in climate conditions may ultimately decrease overall productivity because of decreased abundance (Gherardi and Sala, 2015).
The responses of perennial bunchgrasses to climate change are complex and depend on their photosynthetic pathway type, as well as overarching climate and soil conditions. Although CO2 enrichment may have positive effects on perennial bunchgrasses, changes in temperature and precipitation appear to be more significant drivers of responses. These responses are likely to favor C4 species with increasing annual temperatures (Du and others, 2011; Morgan and others, 2011). However, decreases in precipitation in dry parts of the region are likely to negatively affect all grasses (Shi and others, 2020). Finally, increased interannual variability and drought conditions will have a negative effect on productivity and abundance of perennial bunchgrasses, and resilience will vary at the species level (Havrilla and others, 2023).
1.3.4. Perennial Forbs
Perennial forbs constitute the largest number of species in sagebrush ecosystems, and their responses to climate change will shape the potential future biodiversity of these communities. Perennial forbs have variable responses to climate change factors (table 2). Their interactions with CO2, temperature, precipitation, and extreme weather events are affected by their functional role and regional conditions. One general trend is a shift toward earlier flowering dates, particularly for spring species, such as Phlox hoodii Richardson (spiny phlox) or Balsamorhiza sagittata (Pursh) Nutt. (arrowleaf balsamroot). Outside of core sagebrush areas, especially in the southern part of the region, there may be a decline in perennial forb productivity and species richness because of a decrease in water availability (Pennington and others, 2019). Finally, although perennial forbs may be resilient to low levels of drought and increased variability, the projected increase in the frequency of extreme weather events and multiyear droughts is likely to have negative effects on productivity and species richness across the region (Munson and others, 2013).
Although the understanding of forb responses to CO2 enrichment is limited, studies of dryland forb species show that CO2 enrichment can have a range of effects, especially when considered with other climate variables and human-caused disturbances. Notably, one study found a reduction in forb productivity and fractional cover when exposed to elevated CO2 and nitrogen deposition (Crous and others, 2010), whereas the same treatment had a positive effect on forb species in a Mediterranean-climate annual grassland system (Zavaleta and others, 2003). However, across most studies, forb response was not related to CO2 treatment; instead, it was dominated by the variability of temperature and precipitation among years (Zavaleta and others, 2003, Lee and others, 2011).
Observational studies suggest that perennial forbs in big sagebrush communities are already beginning to have phenological responses to increases in temperature. Flowering has advanced as much as 17 days for early-season species (Bloom and others, 2022). Although temperature has shown negligible effects on photosynthetic rates in some forb species (Loik and Harte, 1996), prairie warming experiments revealed long-term negative effects on forb cover (Morgan and others, 2011). The response of forbs to temperature increases will likely be variable as a functional group; some species benefit from warmer temperatures, whereas others respond negatively, depending on their ecological niche (Lee and others, 2011). One potential general trend for forb species is the reduction of total species number in parts of the sagebrush region where temperature increases will shorten the overall growing season (Cross and Harte, 2007). This trend is consistent with relationships that show species richness decreases in big sagebrush communities as mean annual temperature increases and water availability declines (Jordan and others, 2020).
Responses to precipitation regimes may also be species specific and depend on functional roles. In general, forbs seem to be more responsive to changes in year-to-year precipitation variability than average precipitation levels (de Valpine and Harte, 2001). However, long-term reductions in precipitation in the southern part of the sagebrush region may reduce overall species richness and productivity of forbs, which is consistent with the general species richness and productivity relationship for dryland ecosystems (Maestre and others, 2012).
Interannual variability of precipitation and temperature are controls on forb cover and apparent richness (Zavaleta and others, 2003; Lee and others, 2011). As climate change progresses, the persistence of forb species in big sagebrush ecosystems is likely to be strongly affected by climatic variability.
1.3.5. Invasive Species
The spread of invasive species, including annual grasses and short-lived forbs, is becoming an important topic when discussing the potential future structure and function of sagebrush ecosystems. However, the understanding of invasive species responses to climate change remains limited. Elevated CO2 studies specifically focused on invasive annual grasses and how CO2 enrichment may affect the performance and dynamics of invasive species in sagebrush ecosystems are limited. Consequently, responses of invasive annual grasses to elevated CO2 levels have not been extensively studied, although Blumenthal and others (2016) found no positive effect of CO2 enrichment on cheatgrass in a grassland system.
Temperature changes can profoundly affect the phenology and growth patterns of invasive annual grasses. Observational studies have shown that temperature increases are already affecting the flowering and growth timing of these species (Boyte and others, 2016). These studies have also shown that increases in temperature are correlated with higher invasive annual grass biomass (Howell and others, 2020), which is consistent with experimental warming studies (Blumenthal and others, 2016). Temperature increases will also decrease snowpack, which has been linked to an increase in cheatgrass survivability and establishment (Compagnoni and Adler, 2014a, b; Smull and others, 2019).
The timing of precipitation often determines the success and abundance of invasive annual grasses. Although the response of invasive annual grasses to average precipitation levels remains understudied, their population dynamics are affected by interannual variability in precipitation (Copeland and others, 2019). Variability in year-to-year precipitation patterns can have positive and negative effects on growth, reproductive success, and overall abundance of invasive annual grasses within sagebrush ecosystems (Shriver and others, 2018). Because annuals are dependent on seeds between generations, the presence of a seed bank can facilitate the survival of invasive annual grasses through unfavorable weather (Copeland and others, 2019).
Invasive annual grasses may also be resilient to extreme weather events, such as droughts and fluctuations in temperature. For example, drought may indirectly increase cheatgrass abundance by increasing wildfire probabilities and perpetuating the cheatgrass–wildfire cycle (Coates and others, 2016). The ability of invasive annual grasses to cope with these extremes may depend on the availability of moisture resources, timing and duration of weather, and the overall sensitivity of the invasive annual grass populations (Williamson and others, 2020). Understanding how invasive annual grasses respond to extreme weather events is critical for predicting climate change impacts on their dynamics and persistence.
Understanding the responses of invasive annual grasses to climate change in sagebrush ecosystems is primarily limited to the most prevalent invader, cheatgrass. More research is needed to explore how CO2 enrichment, temperature changes, alterations in precipitation patterns, and extreme weather events will affect the dynamics of other invasive plant populations. Similar to cheatgrass, other invasive annual grasses are likely to spread into core sagebrush habitat as climate change progresses and impacts the ecological functioning of sagebrush ecosystems (Doherty and others, 2022). Potential future research on the responses of other invasive annual grass species to climate change will be essential for effective conservation and management strategies for big sagebrush plant communities.
1.4. Evidence for Climate Change Impacts on Sagebrush Plant Communities from Various Research Approaches
1.4.1. Insights About Climate Effects From Species Distribution Models
Section 1.4.1 Highlights
● Species distribution models (SDMs) describe and compare species observations with environmental conditions to define suitable conditions for a species.
● SDMs can be applied with future projections of environmental conditions to assess how climate change will shift the range of areas that have suitable conditions to support a species.
● Big sagebrush SDMs indicate range shifts to the north and to higher elevations.
● Perennial grass SDMs indicate similar geographic shifts, potentially increased suitability for C4 grasses, and decreased suitability for C3 grasses.
● Cheatgrass SDMs indicate increasing suitability across much of the region.
● SDMs for other plant functional types are often not available.
Species distribution models integrate observed locations of plant functional types or species with environmental variables (primarily climate and potentially soil) to characterize environmental suitability for a species. Projections of environmental suitability can be contrasted with these definitions of environmental suitability for a species to determine how the geographic distribution of areas that can support that species may change under altered climate conditions (table 3).
Table 3.
Summary of species distribution modeling to map potential future distributions of suitable climatic conditions for major plant functional types.[Species distribution models for the current (2024) distribution of suitable climate for invasive annuals and perennial forbs are included in this table because we did not locate any efforts to map potential future distribution of suitable climate. The “Notes” column refers to estimated future shifts or information about current distributions. Potential future disruption refers to studies that estimate potential future (typically middle or late-21st century) changes in suitability, whereas current disruption refers to studies that assess geographic patterns of suitability only under current or recent climate conditions. C3, cool season; C4, warm season]
| Species | Notes | Citations |
|---|---|---|
| Artemisia tridentata Nutt. (big sagebrush) | Declining suitability in the southern part of range and increasing suitability at high elevations and in the northeast edge. | Schlaepfer and others (2012b) |
| Big sagebrush | Declining suitability in the southern and the eastern part of the range; increasing suitability at high elevations and in the northern parts of the range. | Still and Richardson (2015) |
| Big sagebrush | Contraction of the sagebrush range across most of the United States except at high elevations, and an expansion in the northern parts of the range. | Shafer and others (2001) |
| Big sagebrush | In Wyoming, higher temperatures may have a positive effect on sagebrush cover, and less precipitation may decrease suitability in the southwestern part of the State. Overall projections suggest sagebrush distribution is stable for most areas across the State. | Tredennick and others (2023a) |
| Big sagebrush | Projected increase in sagebrush cover across Wyoming because of the relatively low average annual temperature. | Tredennick and others (2016) |
| Big sagebrush | In Nevada, 97 percent of the big sagebrush is vulnerable to one or more of the following risk factors: climate change, land use change, cheatgrass invasion, and conifer expansion. | Bradley (2010) |
| Sarcobatus vermiculatus (Hook.) Torr. (greasewood) | Range expansion into sagebrush habitat at the southern part of the distribution. | Shafer and others (2001) |
| Perennial grasses | Reductions in C3 grass distributions and large increases in suitability for C4 grasses except in the southernmost part of the big sagebrush region. | Havrilla and others (2023) |
| Bromus tectorum L. (cheatgrass) | Increases in suitability for cheatgrass in the eastern and northern parts and reductions in the southwestern parts of the sagebrush region. | Bradley (2009), Bradley and others (2016) |
| Invasive annuals | Expected distributions of multiple invasive, annual species. | Tarbox and others (2022), Shyvers and others (2022) |
| Forbs | Models for multiple species in the Western United States that are important big sagebrush communities. | Barga and others (2018) |
In the sagebrush region, big sagebrush is the most frequently modeled species, and multiple efforts map its current and potential range using various factors such as climate, ecohydrology, and existing land use (Shafer and others, 2001; Bradley, 2010; Schlaepfer and others, 2012b; Still and Richardson, 2015; Tredennick and others, 2023a). Often, the distribution of sagebrush ecosystems is based on the range of big sagebrush and the assumption that it is an umbrella species representing other components of the plant community, such as perennial bunchgrasses and forbs (Tredennick and others, 2023a).
Big sagebrush is in a diverse range of temperature conditions and seasonal precipitation patterns. However, the species is primarily in regions where a substantial amount of precipitation falls during the cool season and can percolate to the deep soil layers (Schlaepfer and others, 2012b). Schlaepfer and others (2012b) defined this ecohydrological niche as conditions that are critical for suitable habitat to support big sagebrush and found that results from their SDM built upon ecohydrological variables generally aligned with traditional climatic SDM results. Ecohydrological and climatic SDMs indicate that the south edge of the sagebrush range is expected to move northward because of higher temperatures and decreased precipitation (Still and Richardson, 2015; Tredennick and others, 2016). The east edge of the big sagebrush range that borders the Great Plains may move westward because of an increase in summer precipitation that favors herbaceous species (Still and Richardson, 2015). Consistent predictions from these models indicate that big sagebrush is likely to expand its range in the northern part of its distribution (Renwick and others, 2018). As temperatures increase, big sagebrush is also projected to expand into high elevations (Bradford and others, 2014). These results are consistent with state-level distribution modeling efforts (Homer and others, 2012) and provide insights into how big sagebrush populations may respond to ongoing climate change. All the SDMs emphasize the importance of considering ecohydrological and climatic conditions when predicting potential future species distributions in the sagebrush region.
The use of big sagebrush as a representative and an umbrella species allows researchers to model and infer the potential climate change impacts on sagebrush ecosystems and other species such as perennial bunchgrasses and forbs. Scientists can use SDMs to begin exploring how climate-induced changes in sagebrush distribution may cascade through the entire ecosystem and impact the ecological dynamics of these valuable habitats. However, understanding the response of a single species may provide only limited information on the potential shifts in distribution for other key functional types.
Species distribution models for subdominant species have received limited attention, and further research in this area is needed. The SDMs for perennial C3 grass species indicate a range expansion at the north edge of their distributions (Havrilla and others, 2023). The SDMs for C4 grass species indicate a more substantial increase in expected range, and particularly in areas dominated by C3 perennial grass species (Havrilla and others, 2023). This predicted shift from C3 suitability to C4 suitability is consistent with process-based modeling projections (Palmquist and others, 2021). The distribution of forb species has been overlooked, and there have been no attempts to create SDMs that incorporate climate change-induced range shifts for species in this functional type. There are few SDMs available for many of the abundant forb species in the sagebrush region (Barga and others, 2018) despite their relevance as forage species for greater sage grouse (Pennington and others, 2016) and pygmy rabbits (Germaine and others, 2020).
There have been more attempts to develop SDMs for invasive species because of their ecological effect and high economic cost (Maher and others, 2013; Chambers and others, 2019). However, most of these SDMs focus on the current distribution of individual species rather than modeling their potential range under climate shifts (Tarbox and others, 2022). Modeling products for invasive annual grass species were reviewed by Tarbox and others (2022) and have an accompanying dataset collection in Shyvers and others (2022). Cheatgrass is one invasive species for which SDMs have been developed; two SDMs developed by Bradley (2009) and Bradley and others (2016) predict the shift in suitability of cheatgrass under projected climate changes across the sagebrush region. Both SDMs indicate an expansion in cheatgrass habitat into parts of the Intermountain West because of a decrease in summer precipitation and increased temperatures (Bradley, 2009; Bradley and others, 2016). The most recent SDM projects a decline in cheatgrass suitability in the southern part of the Great Basin where temperature increases and precipitation declines will reduce habitat for C3 annual grasses (Bradley and others, 2016).
Limitations of Species Distribution Models Results and Research Needs
1.4.1.1
Despite the high conservation value of big sagebrush plant communities, their responses to disturbances such as grazing and, to a lesser extent, climate change, is oversimplified. This oversimplification is evident in SDMs that primarily focus on the current (2024) distribution of big sagebrush. However, sagebrush ecosystems are composed of a diverse array of species, and each contribute to the ecological functioning of these habitats. Nondominant grass species in the sagebrush region are essential for wildlife and livestock forage, and understory grass or forb species represent the largest component of overall biodiversity of these ecosystems (Pennington and others, 2019). Thus, there is a need for modeling to focus on mapping the projected future distributions of relevant nondominant species in the sagebrush region. Expanding SDMs to include future climate projections for nondominant species, including invasive grass and forb species other than cheatgrass, may be the most effective way to incorporate consideration of climate change into management decisions. Potential future research on subdominant species will contribute to a more nuanced understanding of sagebrush plant communities and their responses to changing environmental conditions.
1.4.2. Plant Community Response Types to Climate Change Based on Observed or Experimental Studies
Section 1.4.2 Highlights
● Big sagebrush plant communities may benefit from climate change in some areas, notably at high elevations and in the northern Great Plains.
● Increasing winter moisture in some locations may benefit woody plants, notably big sagebrush and conifers, whereas perennial grasses may increase in areas with elevated summer moisture and increased spring temperatures.
● Warmer conditions with more cool-season moisture may accelerate C3 annual grass invasion and increase wildfire frequency and size.
● As temperatures rise, cover of bare ground is likely to increase in areas with stable or decreasing precipitation.
This section reviews climate change impacts on sagebrush plant communities by examining experimental studies that manipulate single or multiple climate variables. Unlike the potential climate change effects estimated in SDMs, experimental studies provide insights into local to landscape-scale expectations of climate change effects and potential threats that shape community-level responses to climate change.
This section includes various experimental approaches (fig. 3), such as space-for-time substitutions, experimental warming, precipitation shifts, and CO2 enrichment experiments (Harte and Shaw, 1995; Bates and others, 2006; Jordan and others, 2020). Multiple response types observed for plant communities that are affected by local environmental factors including the prevailing climate were identified. Different response types are framed by their resistance and (or) resilience to compositional shifts in big sagebrush plant communities. The following response types were defined:
-
1. Native big sagebrush plant communities with decreased climate sensitivity,
-
2. Conifer expansion and increased sagebrush dominance,
-
3. Increased susceptibility to invasive annual grasses,
-
4. Increased herbaceous cover, and
-
5. Increased bare ground.
These response types help describe how big sagebrush plant communities may resist or adapt to climate-induced changes and identify the pathways through which these communities may change because of ongoing environmental shifts.


Photographs showing two examples of experimental methods to assess sagebrush plant community response to climate change: A, water addition treatments to simulate changes in precipitation seasonality (Photograph by Scott Carpenter, Yale University, School of the Environment) and B, rainout shelters which simulate increased drought frequency or decreases in total precipitation (Photograph by Martin Holdrege, contractor to the U.S. Geological Survey)
Native Big Sagebrush Plant Communities With Decreasing Climate Sensitivity
1.4.2.1
Big sagebrush communities with high resistance to changing climate tend to be at the highest elevations and in the parts of the Great Plains that include core big sagebrush habitat (Doherty and others, 2022). In these communities, changes to soil moisture availability resulting from altered climate conditions that extend the growing season are likely to have minimal negative and potentially positive effects on community composition and functioning. However, no precipitation manipulations as of 2024 have shown a consistent increase in herbaceous fractional cover.
All big sagebrush plant communities may benefit from the rising levels of CO2, which can improve water-use efficiency for big sagebrush as well as herbaceous species (Lucash and others, 2005; Lee and others, 2011). Moreover, in certain areas with cooler climate conditions, increased temperature has shown to promote the growth and reproductive success of big sagebrush and some C4 grass species and indicates the potential for a shift in perennial grass dominance near the end of the 21st century (Perfors and others, 2003; Williams and others, 2007).
Under experimental warming, these resistant and benefiting big sagebrush plant communities have demonstrated increased seed production and better seeding establishment rates that contribute to the expansion of native perennial plant populations in high-elevation mountain meadows (Harte and Shaw, 1995). The response potential of these resistant high-elevation communities is evident in their ability to adapt to shifts in interannual variability of precipitation, and, despite losses in productivity, maintain overall abundance of perennial species (Tredennick and others, 2018). Further, these communities may be resistant and resilient to cheatgrass establishment even under increased temperatures (Larson and others, 2017), although decreased snowpack may decrease this resistance (Compagnoni and Adler, 2014b). The high cover of native perennial bunchgrasses in these sagebrush communities likely provides high levels of resistance to invasion by cheatgrass and other invasive annual grasses (Compagnoni and Adler, 2014a). Additional experimental evidence is needed to assess the continued resistance of big sagebrush plant communities to invasive annual grasses under various aspects of climate change other than increased temperatures, including shifting precipitation patterns and enhanced weather variability.
These resistant and resilient high-elevation communities show a notable capacity to withstand the challenges posed by climate change, and sometimes, capitalize on the changing environmental conditions. Their ability to maintain or even improve their resistance to invasive annual species suggests that these communities may be able to persist and resist invasion even as the climate changes. However, the resistance and benefits observed in these communities may not occur under all climate change scenarios. The actual response of these communities will vary depending on regional characteristics, local climate conditions, levels of current and potential future herbivory, and the specific composition of each community. Additionally, the mechanisms driving the resilient responses of these big sagebrush plant communities warrant further investigation to determine whether these communities will continue to show resistance and benefits as climate change progresses.
Conifer Expansion and Increased Sagebrush Dominance
1.4.2.2
Conifers, notably Pinus edulis Engelm. (pinyon pine) and P. juniperous spp. (juniper), are predicted to increase in cover across some parts of the sagebrush region (Remington and others, 2021). In certain big sagebrush plant communities, climate change is projected to favor an increase in woody plant growth and cover, including sagebrush, particularly in response to shifts in climate that increase the availability of soil moisture deep in the soil profile (Holdrege and others, 2023). Under conditions where deep soil moisture availability increases, projected increases in precipitation during the late fall through spring are likely to benefit big sagebrush as well as coniferous tree species in pinyon-juniper woodlands (Weisberg and others, 2007).
Another possible driver of increased woody plant cover is the rise in atmospheric CO2, which promotes enhanced growth and biomass accumulation in woody plants (Curtis and Wang, 1998). Elevated CO2 may boost the productivity and competitive advantage of woody species (Cabral and others, 2003). Experimental warming studies have found that rising temperatures can create more favorable conditions for woody plant establishment and growth at high-elevation sites (Harte and Shaw, 1995; Perfors and others, 2003). As temperatures increase, woody plants in the sagebrush region may experience improved reproductive success, earlier budbreak, and increased seedling recruitment particularly at high-elevation sites (Perfors and others, 2003). The combination of these responses can facilitate conifer expansion or increases in sagebrush cover.
Changes in precipitation patterns can also affect the distribution and abundance of woody plants in sagebrush ecosystems. Increases in winter or spring precipitation will increase water availability, which will increase growth of woody species and enable them to establish more successfully (Maier and others, 2001). Although shifts in the seasonality of precipitation may favor the expansion and persistence of sagebrush species, increases in total precipitation may promote the expansion of pinyon pine and juniper into sagebrush plant communities (Weisberg and others, 2007).
The increase of woody plant cover has important implications for the overall structure and function of big sagebrush plant communities. Where conifer species are established, there will be shifts in plant species composition and alterations in plant community structure. These changes in moisture resource availability for other plant and animal species can have cascading effects on ecosystem dynamics, including shifts in wildlife habitat and altered fire regimes. If the increase in woody plants enhances big sagebrush dominance, the changes in structure and function will be small. By contrast, if the increase in woody plants includes pinyon pine or juniper, much larger changes in the structure and composition of big sagebrush plant communities may be expected. It is also possible for increased sagebrush dominance to reduce forage availability for livestock, wildlife, feral horses and burros.
Increased Susceptibility to Invasion
1.4.2.3
In parts of the Great Basin and lower-elevation sites across the Intermountain West, climate change is projected to create conditions that enhance the vulnerability of these big sagebrush plant communities to invasion by nonnative annual grasses (Zimmer and others, 2021). Conditions that may be associated with increased susceptibility to invasive annual grasses include decreases in soil moisture availability and snowpack, which likely negatively affect native perennial grasses.
Increased climatic suitability for invasive species can increase the probability that they will outcompete native plants, disrupt the natural fire cycle of sagebrush ecosystems, and limit sagebrush regeneration (Davies and Svejcar, 2008; Pilliod and others, 2017). Additionally, wildfires, which are expected to increase in frequency and intensity under climate change, can facilitate the spread of invasive species like cheatgrass (Balch and others, 2013; refer to sec. 2.4. “Potential Climate Change Effects on Recreation in Big Sagebrush Plant Communities”).
Increased vulnerability to invasion can be facilitated by decreases in moisture availability, which improves conditions for invasive annual grasses because of their resilience in low soil water conditions (D’Antonio and Vitousek, 1992). These low soil moisture conditions can also decrease native, perennial herbaceous abundance (Lozano-Parra and others, 2018). Because of reduced competition from native species, cheatgrass may be able to spread rapidly across the landscape. Enhanced susceptibility to invasive annual grasses may also be facilitated by feral horses (King and others, 2019). Further, the increased length of the growing season and warmer temperatures can provide an extended window for invasive annual grass establishment that enhances their ability to outcompete native species in colder parts of the sagebrush regions.
Big sagebrush plant communities are vulnerable to invasive annual grass species that have negative effects on these communities. As cheatgrass and other invasive plant species expand into regions, such as the Great Plains, they can alter the structure and functioning of communities, displace native vegetation, and alter the disturbance regime by increasing fire return intervals (Pilliod and others, 2017; Molvar and others, 2024). Although there are calls to continue cheatgrass eradication efforts in large parts of the region, the potential permanence of cheatgrass is also being recognized in some areas, particularly in the northern Great Basin (Davies and others, 2021). This recognition of long-term cheatgrass persistence is likely to expand in parts of the sagebrush region as climate change progresses.
This section primarily focuses on the increased vulnerability of sagebrush plant communities to cheatgrass invasion because most of the research to date (2024) has focused on cheatgrass and its interactions with climate and the fire cycle. However, the potential for changing climate to enhance threats from other invasive annual grasses such as medusahead, is unknown. Future experimental work would be helpful to begin exploring the response of less prevalent invasive plant species to projected climate changes.
Increased Herbaceous Dominance
1.4.2.4
In parts of the Intermountain West, climate change is expected to facilitate a transition from sagebrush dominance to perennial grass dominance, where grasses may replace sagebrush as the highest cover component (Doherty and others, 2022). One suggested driver of this transition is the well-documented increase in temperature under climate change (IPCC, 2022). As temperatures rise, high-elevation sites may experience more favorable conditions for the growth and reproduction of perennial herbaceous species. For example, perennial grasses and forbs that are adapted to warm temperatures may become more competitive in these areas and lead to an expansion of their distribution and overall abundance (de Valpine and Harte, 2001; Dolezal and others, 2021).
Changes in seasonal patterns of precipitation is the aspect of climate change that is most likely to facilitate increased dominance of perennial grasses. In regions where an increase in precipitation during the summer is projected, conditions will become more favorable for perennial grasses (Skinner and others, 2002; Volder and others, 2013). Most experimental studies show small, slow changes in grass cover after 2–3 years of treatment (Adler and others, 2009). However, changes in water availability that increase the proportion of soil water available in the upper part of the soil can enhance the productivity and competitive ability of perennial grasses (Walter, 1973). A shift from sagebrush to perennial grass dominance will affect ecosystem structure and function. For example, communities with increasing dominance by perennial grasses will have a higher forage yield for livestock species but a reduced habitat value for sagebrush obligate wildlife species like greater sage grouse.
Increased Bare Ground
1.4.2.5
As temperatures increase and precipitation decreases in the southern Great Basin, native perennial species in big sagebrush plant communities and invasive annual grasses are expected to decline in abundance. This decline will lead to increased bare ground. An increase in bare ground was observed in parts of Wyoming, Nevada, and other States that represent the driest parts of the region (Bradley, 2010; Xian and others, 2012; Shi and others, 2018, 2020). Decreased precipitation and increased temperatures will decrease soil water availability, lead to lower production and eventually decreased sagebrush and perennial herbaceous species abundance, and leave more areas of bare soil exposed (Bates and others, 2006).
Increased frequency and intensity of disturbances associated with climate change, such as wildfires and droughts, also contribute to increased bare ground. More frequent and severe wildfires can lead to widespread vegetation loss that creates bare landscapes (Coates and others, 2016). Additionally, droughts can increase vegetation stress and mortality, which further reduces plant cover and increases bare ground (Winkler and others, 2019). The presence and high density of feral horses and burros can also increase the prevalence of bare ground (Beever and others, 2008). There is a potential for a substantial increase in soil erosion because of less plant cover. Decreased plant cover can also affect wildlife habitat by reducing vegetation for food, shelter, and nesting sites. Changes in precipitation patterns, increased disturbances, rising temperatures, and human activities collectively contribute to the loss of plant cover and the increase in bare soil. Minimizing grazing pressure from managed livestock and feral horses and burros may improve soil retention and biotic crust formation (Muscha and Hild, 2006).
Limitations of Observational or Experimental Studies
1.4.2.6
There are few manipulative or observational field experiments about the impact of changing climate on big sagebrush plant communities. As a result, these studies cannot provide a comprehensive understanding of long-term impacts. Field studies are valuable but do not fully capture the complex and interconnected dynamics of big sagebrush plant communities across extended periods. Section 1.4.3. “Modeling Region-Wide Effects of Climate Change on Big Sagebrush Plant Communities” addresses this limitation by reviewing modeling studies that project changes in community compositions by modeling the response of sagebrush habitat or the response of multiple functional groups. These models provide predictions for the persistence of core sagebrush habitat across the region (Doherty and others, 2022). The analysis can be extended beyond individual field experiments by including model projections to gain insights into how climate change may affect sagebrush plant communities on larger spatial and temporal scales.
1.4.3. Modeling Region-Wide Effects of Climate Change on Big Sagebrush Plant Communities
Section 1.4.3 Highlights
● Most big sagebrush plant communities are expected to experience small changes under future conditions across much of the region.
● Under future climate scenarios with higher temperatures, sensitive areas that are unlikely to maintain big sagebrush plant communities are more extensive than they are under scenarios with lower potential future temperatures.
● Shifts in the composition and abundance of understory plant functional types are more likely to affect management decisions related to livestock grazing and recreation than shifts in big sagebrush abundance.
● The persistence of big sagebrush plant communities will likely be diminished where annual grass invasion and enhanced wildfire frequency are most probable.
Five response types identified in section 1.4.2. “Plant Community Response Types to Climate Change Based on Observed or Experimental Studies” highlight possible trajectories of plant communities within the sagebrush region. However, because of the limited spatial and temporal scope of field studies, these response type categories do not completely capture the potential effects of climate change on big sagebrush plant communities at regional (for example, multi-State) and long-term (for example, multidecadal) scales.
There are multiple methods for modeling how shifts in climate will impact the suitability of big sagebrush plant communities, including process-based and statistical models (refer to sec. 3.3. “Analytical Approaches for Assessing Potential Long-Term Effects of Climate Change on Plant Communities” for further description). Studies assessing potential geographically broad shifts in climatic suitability for big sagebrush plant communities can complement findings from individual field studies. This section discusses how the results of these plant community-level efforts overlap and identifies aspects of consistency or divergence with other types of studies described previously.
All available studies that examine community-level climate change effects in big sagebrush ecosystems consider the effects on big sagebrush suitability. A few studies focus on modeling climatic suitability for the big sagebrush plant community (Doherty and others, 2022). Some studies use big sagebrush as an umbrella species (Schlaepfer and others, 2021; Tredennick and others, 2023a) to represent potential effects on the broader plant community. Other studies model projected shifts in climate suitability for individual plant functional types within big sagebrush plant communities (Palmquist and others, 2021; Rigge, Shi, and Postma, 2021; Zimmer and others, 2021).
For most models, big sagebrush is expected to remain viable across much of the sagebrush region under the moderate emissions RCP4.5 and the higher emissions RCP8.5 (Palmquist and others, 2021; Rigge, Shi, and Postma, 2021; Zimmer and others, 2021) and under the new shared socioeconomic pathway (SSP) climate scenarios SSP4.5 and SSP8.5 (Tredennick and others, 2023a). The areas of sustained climatic suitability for big sagebrush plant communities overlap with the areas identified as core habitat (for example, abundant big sagebrush and perennial grasses with minimal annual grasses, human effects, or conifers) by Doherty and others (2022). Schlaepfer and others (2021) found that regeneration across some of the areas of stability will be negatively affected under RCP8.5.
Projections of geographic patterns of big sagebrush response to climate change can vary among studies. Most models project declines in climatic suitability at the southern part of the sagebrush range and are consistent with most field experiments and observations (Palmquist and others, 2021; Schlaepfer and others, 2021; Zimmer and others, 2021). However, this pattern is not fully supported by Rigge, Shi, and Postma (2021) projections for sagebrush cover under RCP8.5, which showed moderate declines in cover in the northern part of the range and little change in the southern part of the range. Rigge, Shi, and Postma (2021) project a loss in sagebrush cover in parts of Montana under RCP8.5, which is consistent with other models using this climate scenario (Palmquist and others, 2021). Other projections suggest either minimal changes to sagebrush suitability (Zimmer and others, 2021) or increases in climatic suitability by mid-21st century (Palmquist and others, 2021), which may indicate that the RCP8.5 for the end of the century is an extreme scenario. Models are also split on the expansion of big sagebrush climatic suitability at high-elevation sites (Palmquist and others, 2021; Zimmer and others, 2021). Despite the differences in the projected responses of climatic suitability for big sagebrush across models, most conclude that shifts in suitability will be small. Instead, the interactive effects of climate with disturbances and invasive species may affect most of the region. All models suggest that these interactive effects are important areas for potential future research.
Beyond shifts in climatic suitability for big sagebrush, climate change is likely to impact the composition of grasses and forbs across the region. Only Rigge, Shi, and Postma (2021) and Palmquist and others (2021) independently project shifts in the suitability for herbaceous functional types. Zimmer and others (2021) project shifts in total forage availability but do not differentiate plant functional types within that group. Rigge, Shi, and Postma (2021) and Palmquist and others (2021) project increases in suitability for perennial herbaceous species across most sagebrush plant communities in Wyoming and Colorado, but project decreases in the southern part of the range. Zimmer and others (2021) project consistent forage increases. All three studies project decreases in forage or perennial herbaceous species across most of the southern parts of the big sagebrush region except in some high-elevation areas (Palmquist and others, 2021; Rigge, Shi, and Postma, 2021; Zimmer and others, 2021), which is consistent in areas with observed increases in bare ground and similar climatic conditions (Shi and others, 2018). Palmquist and others (2021) is the only study to separate C3 and C4 perennial grasses, which make up the largest component of forage in most of the sagebrush region. They projected decreases in the biomass of C3 species and potential increases in the biomass of C4 species, which may indicate that replacement of C3 species as climate change progresses (Palmquist and others, 2021). This potential regional shift in relative biomass of C3 compared to C4 grasses is consistent with Havrilla and others (2023) species distribution models for C3 and C4 perennial grasses.
Climate change may also affect the severity of invasive or locally nonnative species that alter big sagebrush plant communities. Although little is known about how cheatgrass and conifers will interact to affect big sagebrush plant communities in the future, Zimmer and others (2021) suggested that potential future climate conditions may limit the expansion of conifers, but the potential for conifer expansion varies substantially among the climate models evaluated in their study.
1.4.3.1.Tying Climate Change to Vegetation Across Scales and Disturbances
The effects of projected changes in climate on big sagebrush plant communities are likely to vary across the sagebrush region. Despite this geographic variability, and variation among studies and methods, a few general trends emerge:
-
1. Big sagebrush and other woody species are likely to benefit from increased fall and winter precipitation that promote species growth and continued stability (Perfors and others, 2003; Schlaepfer and others, 2012a; Palmquist and others 2016). This benefit is especially true in high-elevation sites where temperature increases may extend the ecohydrological growing season (warm conditions with sufficiently wet soil to support plant growth; de Valpine and Harte, 2001).
-
2. Warming is likely to have the opposite effect at the south edge of the region and in low-elevation mountain basins where temperature increases during the summer may be linked to decreases in water availability and lower sagebrush growth (Schlaepfer and others, 2012b; Renwick and others, 2018). Extreme temperatures may cause stand-level mortality, although no studies have modeled sagebrush mortality under future climate conditions (Renne, Schlaepfer, and others, 2019).
-
3. Herbaceous functional types are expected to follow a similar trend as woody species and have productivity increases in northern and high-elevation areas and productivity decreases at southern and low-elevation intermountain areas (Shi and others, 2018; Palmquist and others, 2021; Rigge, Shi, and Postma, 2021).
This review of the available scientific literature (refer to the “Methods for Developing this Science Synthesis” section) describes uncertainties about the interactive effects of climate change, wildfire, and predicted co-occurrence of high temperatures and drought. The most well-understood interaction is between climate change and wildfire in areas already invaded by annual grasses like cheatgrass (Bradley and others, 2018; Pastick and others, 2021). Cheatgrass senesces earlier in the growing season than native grasses and creates dry, fine fuels. Fire return intervals have shortened substantially in some parts of the sagebrush region because of cheatgrass invasion, and the area burned each year increased between approximately 1980 and 2020 (Baker, 2013; Balch and others 2013; Shinneman and others, 2023). Plant community degradation occurs when postfire establishment of sagebrush is hindered by the presence of cheatgrass (Coates and others, 2016). Climate change is projected to increase the prevalence of extreme fire weather (Bowman and others, 2020; Coop and others, 2022). How climate change and cheatgrass will affect potential future wildfire frequency is uncertain and will likely vary across the region. Areas where annual grass invasion becomes more common under hotter conditions are likely to have the largest increase in wildfires (Smith and others, 2023). It is also possible that with climate change, some hot and dry areas, primarily in the southern part of the region, which already support little plant growth, may become more fuel limited, and therefore experience fewer wildfires through time.
The role that wildfire and invasive species play in big sagebrush ecosystems is one example of how disturbance dynamics can affect potential climate effects on plant communities. The combination of wildfire and invasive species has transformed large parts of the big sagebrush region into plant communities dominated by invasive species rather than native plant species (Doherty and others, 2022), and climate change is expected to amplify this process (Holdrege and others, 2024a) by increasing the probability that native big sagebrush plant communities are replaced by invasive species.
In addition to wildfire, other disturbances may interact with climate change to affect big sagebrush plant communities. Enhanced frequency and severity of extreme weather may increase the prevalence of perennial plant mortality (Winkler and others, 2019). Likewise, longer and hotter summer growing seasons may decrease the resilience of forage resources to land-use practices (Klemm and others, 2020) and may exacerbate the existing challenge of restoring native big sagebrush plant communities (Germino and others, 2018).
2. Implications of Climate-Driven Shifts in Sagebrush Plant Communities for Decisions and Land Uses
Research suggests that changing climate conditions may lead to an expansion of suitable habitat for big sagebrush in the northeastern part of the sagebrush region (Schlaepfer and others, 2015; Palmquist and others, 2021; Rigge, Shi, and Postma, 2021) and at high-elevation parts of its range (Perfors and others, 2003) and a decrease in southwestern and low-elevation parts of the region (Xian and others, 2012; Homer and others, 2015; Palmquist and others, 2021). However, regional patterns may not always align with local responses (Levin, 1992). This section describes how climate-induced changes in big sagebrush plant communities may relate to BLM land use and resource management decisions and identifies likely effects of climate change that can inform decision making in individual BLM field offices.
2.1. Implications of Climate Change for Forage Resources and Grazing Decisions in Sagebrush Ecosystems
Section 2.1 Highlights
● Potential future changes in forage resources will vary across the sagebrush region, and much of this variation can be explained by climate-driven variation in two variables: total precipitation and length of the warm, wet growing season.
● Areas with more precipitation and longer warm, wet growing seasons may have forage increases, whereas forage may be relatively unchanged in areas with stable or decreasing precipitation and longer warm, wet growing seasons.
● Areas with increased precipitation and shorter warm, wet growing seasons may experience stable forage amounts but shifting seasonality. Forage availability is likely to decline in areas with decreased precipitation and shorter warm, wet growing seasons. These changes will have different implications for grazing management.
Climate change impacts on forage resources are likely to vary substantially across the broad big sagebrush region. Although most models indicate that climate change will likely enhance the long-term viability of most native species in sagebrush plant communities in cool regions, and reduce long-term viability in warm areas, the response of perennial grasses is less likely to follow the same large-scale pattern (Palmquist and others, 2021). Potential future changes in the timing and quantity of forage production in big sagebrush plant communities will be largely dictated by climate-driven changes in two ecologically relevant variables: the ecohydrological growing season, and total precipitation. In this section, we identify four categories of climate change impacts that summarize these divergent impacts. Variation among climate models in potential future ecohydrological growing season length and total precipitation means that the exact locations of these response categories cannot be precisely mapped; however, general expectations for geographic patterns for each response types are identified. The four impact categories are the following:
-
1. Category One—Increased ecohydrological growing season length and increased precipitation.
-
2. Category Two—Increased ecohydrological growing season length and static or decreased precipitation.
-
3. Category Three—Decreased ecohydrological growing season length and increased precipitation.
-
4. Category Four—Decreased ecohydrological growing season length and decreased precipitation.
For each of these impact categories, the expected changes in the production of forage are outlined and how these shifts may affect livestock grazing are assessed. The impact categories are associated with different threats to forage species such as increased suitability for invasive annual grasses (that is, cheatgrass) or conifer encroachment. Invasive annual grasses and conifer encroachment are discussed within the following subsections exploring each impact category. This section concludes by discussing how interannual variability in precipitation and the frequency of anomalous climate conditions, such as intense droughts, will modify regional patterns for forage timing and production.
2.1.1. Category One—Increased Ecohydrological Growing Season Length and Increased Precipitation
In areas that are projected to have a Category One impact, the overall consequence will be positive for livestock grazing with increased production and an extended growing season. However, management will need to monitor the woody component of plant communities to be aware of possible increases. The primary high confidence consequence of climate change is increasing temperatures, which is the expected main driver of any projected shifts in areas with climate conditions that support big sagebrush (Kleinhesselink and Adler, 2018; Renwick and others, 2018). At the north edge of their range and in high-elevation sites, forage grass and forb species are expected to benefit from higher temperatures because of an extended ecohydrological growing season. This benefit is because of an increase in growing degree days that leads to an increase in aboveground production (Schlaepfer and others, 2012a; Palmquist and others, 2016; Renwick and others, 2018). Additionally, the phenology of herbaceous species is likely to advance as snowpack melts earlier (Bloom and others, 2022). This advancement in phenology is likely to lengthen the livestock grazing season.
The northeast edge of the big sagebrush region and some high-elevation sites are projected to experience increased precipitation (fig. 2), which can also result in an increase in forage production. The timing and amount of precipitation determine water availability at different depths within the soil profile and the competitive advantage of either herbaceous or woody species (Schlaepfer and others, 2012a; Lauenroth and others, 2014; Renne, Schlaepfer, and others, 2019). Most precipitation increases are expected to occur in the late fall and early spring (Palmquist and others, 2016). As precipitation occurrences become larger or happen during cooler periods, a greater proportion of water will percolate to deep soil layers and make it less accessible to forage species and more favorable for woody species (Holdrege and others, 2021, 2023). In locations such as northern Montana and the upper Green River Basin in Wyoming, the projected increase in production would benefit forage species, but it may also increase big sagebrush cover and abundance. Northern Montana and the upper Green River Basin in Wyoming represent locations with expected forage increases. Although other areas across the big sagebrush region may experience increases in productivity, it is also possible that slow plant response to rapid climate change may lead to lower overall productivity by the end of the 21st century (Felton and others, 2022).
2.1.2. Category Two—Increased Ecohydrological Growing Season Length With Decreased or Static Precipitation
In areas that are projected to have a Category Two impact may likely be the most stable areas within the big sagebrush region. It is expected that the longer growing season length, through increased plant growth potential during the early spring season (Bradford and others, 2020) will benefit herbaceous species, thereby promoting the production of forage and reducing bare ground. The largest threat to big sagebrush plant communities under these conditions is an increase in suitability for invasive annual grasses. Cheatgrass completes its seasonal growth cycle early in the year (Harris, 1967), and benefits from an advanced growing season and reduced snowpack (Compagnoni and Adler, 2014b; Smull and others, 2019181). Consequently, high-elevation sites that have previously been resistant to cheatgrass establishment may experience an increase in cheatgrass abundance, which would reduce forage availability. However, high cover of C3 perennial bunchgrasses can sometimes decrease the probability of cheatgrass invasion (Chambers and others, 2014). Management of high-elevation communities, such as in the intermountain parts of Colorado, Nevada, and Utah, may benefit from monitoring to identify areas of nonnative plant establishment before they reach critical abundance levels (Chambers, Maestas, and others, 2017).
For livestock grazing, areas with a Category Two impact could have an increase in the growing season length that advances the phenology of forage species such that grazing may begin earlier in the year to capitalize on earlier spring production. Thus, an increase in forage production may enable increases is animal stocking rates in these areas. However, any decision to increase grazing intensity may need to consider perennial bunchgrass cover and monitor to ensure that cover either increases or is stable to minimize the establishment of nonnative species.
2.1.3. Category Three—Decreased Ecohydrological Growing Season Length and Increased Precipitation
Areas that are projected to have a Category Three impact will likely represent a smaller part of the sagebrush region than Categories One or Two impacts. Similar to Category Two impacts, Category Three impacts represents relative stability in potential future forage production. Areas likely to experience these conditions include parts of the Great Basin and the Colorado Plateau. The decrease in the ecohydrological growing season is because of the extended time when potential evapotranspiration exceeds precipitation in the spring and fall, which decreases water availability for plant growth. This decrease is likely to reduce the overall production of forage species. In addition, most of the precipitation increase is expected to occur in the late fall and early spring (Palmquist and others, 2016) and the amount of precipitation is likely to increase (IPCC, 2022). Studies that simulate both changes have demonstrated that this precipitation regime is most likely to favor woody species more than perennial grasses similar to the Category One impact (Bates and others, 2006; Holdrege and others, 2021).
Shifts in seasonal and soil depth patterns of soil moisture availability under the Category Three impact conditions suggest the primary consequence will be a shift in growing season rather than an increase in forage production. This shift toward warmer, drier conditions may increase climatic suitability for C4 perennial grass species (Palmquist and others, 2021). If a shift toward C4 grasses occurs, managers may need to consider the later green up of C4 species in their planning. Under this response category, cheatgrass persistence and establishment is still a threat.
Considering these potential changes in the composition of big sagebrush plant communities, managers may consider three steps when monitoring livestock grazing allotments in areas that are experiencing decreased ecohydrological growing season length and increased precipitation:
-
1. Adopting a woody plant monitoring protocol that focuses on shifting fractional components of woody and forage species, particularly perennial grasses.
-
2. Take steps to promote the establishment of C4 grasses, particularly in areas with low perennial bunchgrass cover.
-
3. In conjunction with efforts to establish C4 grasses, consider the potential for a new early season period for grazing to target early annual species such as cheatgrass.
2.1.4. Category Four—Decreased Growing Season Length and Decreased or Static Precipitation
Areas that have a projected Category Four impact represent the south edge of the big sagebrush distribution, where most modeling efforts indicate that temperature increases will likely decrease the growing season by increasing evaporative demand at the beginning and end of the thermal growing season. This decrease can thereby reduce soil water availability and accelerate senescence (Bradford and others, 2020; Maurer and others, 2020126). In northern New Mexico and Arizona, forage species are likely to decrease in production (Kleinhesselink and Adler, 2018; Shi and others, 2018) and ultimately reduce available forage for livestock grazing. It is probable that a reduction in livestock grazing may be necessary to ensure the persistence of native species and to prevent the establishment of nonnative species. Furthermore, the shortened ecohydrological growing season may necessitate a reduction in grazing period length.
Overall climatic suitability for big sagebrush and other native perennial species is also expected to diminish under the Category Four impact category. Monitoring efforts may benefit from focusing on quantifying the extent bare ground cover increases that results from the loss of biocrust and perennial grass species.
2.1.5. Interannual Variability and Extreme Events
The growing likelihood of extreme climate is an added complexity to the four impact categories (Parmesan and others, 2000; Smith, 2011). More extreme weather events can directly impact forage production and have lasting effects (Hoover and others, 2014, 2015). Although extreme weather events may affect woody species, big sagebrush has some drought resistance (K.J. Clause and J. Randall, Bureau of Land Management, written commun., 2014; Hoover and others, 2015). By comparison, most herbaceous forage species do not have similar drought resistance; for example, C3 perennial grasses recover slowly following mortality events (Hoover and others, 2015, Breshears and others, 2016), although they may be resilient on decadal scales.
Beyond extreme weather events, interannual variability in precipitation can affect the production of perennial grasses within big sagebrush communities (Hsu and others, 2012; Gherardi and Sala, 2019; Hou and others, 2021), although this variability may not affect the coexistence of herbaceous and woody species (Adler and others, 2009). Consistent forage production is unlikely under response categories that have increasing interannual variability in precipitation, and managers might anticipate fluctuations in production as interannual variability rises (Klemm and others, 2020).
2.1.6. Overall Implications for Forage Resources
The impact categories follow the general expectation that the northeastern part and high-elevation areas within the range of big sagebrush will likely experience overall increases in forage production, whereas the southeastern part and low-elevation areas will experience decreases. Despite the positive outcomes in the northeastern and high elevation areas, the anticipated changes in seasonality and the increasing amounts of precipitation are more likely to favor big sagebrush more than forage species. Moreover, increased interannual variability and the increasing rate of extreme weather events is expected to have negative effects on forage production. Given these circumstances, a one-size-fits-all future management plan for livestock grazing in big sagebrush ecosystems is not feasible. Instead, managers may need to carefully consider the specific climate projections for forage species in their region to determine if forage species in grazing allotments will benefit or suffer from rising temperatures and changing precipitation.
In favorable conditions, vegetation monitoring in grazing allotments may incorporate a focus on the growth of woody and invasive species. This approach will enhance the understanding of how changing precipitation is affecting the balance between shrubs and herbaceous species, particularly perennial grasses. In unfavorable conditions, managers may need to consider shifting the grazing dates for allotments and potentially eventually reducing the overall animal use months as well. Vegetation monitoring would benefit from focusing on the persistence of C3 and C4 grasses. Finally, managers may need to continuously monitor drought indicators and remain prepared to adjust grazing intensities in the aftermath of drought periods.
2.2. Implications of Climate Change for Wildfire Management in Sagebrush Ecosystems
Section 2.2 Highlights
● Warmer temperatures enhance fire frequency, the length of the fire season, and fire size, although in dryland regions wildfire is also limited by fuel availability.
● Changes in the abundance of invasive annual grasses and perennial grasses will affect wildfire dynamics.
● Management strategies for addressing increases in wildfire risk may include invasive grass suppression techniques, mechanical fuels treatments, and targeted grazing.
Wildfire frequency and severity are projected to increase across much of the sagebrush region (Holdrege and others, 2024b). Although there is extensive work reviewing the effects of fire on sagebrush ecosystems, most studies focus on the interactions of fire and invasive annual grasses (Davies and others, 2011; Chambers and others, 2019). Few studies have considered the interaction of climate change and fire within big sagebrush plant communities (Abatzoglou and Kolden, 2011). The projected increase in wildfires within the big sagebrush range emphasizes a need for adaptive management practices to limit the accumulation of fine fuels and prepare for increased fire probability in the future (Chambers and others, 2014; Coates and others, 2016).
Projected shifts in climate across the sagebrush region are already affecting wildfire frequency and severity. Increases in annual temperature and variability of precipitation across the region are expected to alter fuel loads (Applestein and others, 2021), enhance fuel continuity because of increased cheatgrass (Germino and others, 2016), and lower fuel moisture content (Turco and others, 2017). These shifts are especially likely in some short droughts during which lower fuel moisture can make vegetation more susceptible to ignition and promote fire spread (Flannigan and others, 2016). Interannual variability can also cause an increase in fuel buildup during wet years, which can lead to increased fire severity during dry periods when ignition risk is greater (Pilliod and others, 2017).
Climate associated shifts in vegetation are also likely to affect fire cycles (Bradley, 2009; Shi and others, 2020). In particular, invasive annual grasses have increased since the 1990s throughout much of the region (Smith and others, 2023), and fire probability is greatest in areas with high annual grass abundance (Holdrege and others, 2024b). As sagebrush communities shift in response to climate change, lowered resistance to invasion by nonnative annual grasses and decreased resilience to wildfire are expected (Chambers and others, 2023; Schlaepfer and others, 2024). These decreases in resistance and resilience can promote cheatgrass establishment or increase the vulnerability of big sagebrush communities ecological transformation toward altered states following fire (Boyte and others, 2016; Barker and others, 2019). Cheatgrass is strongly linked to altered fire cycles because it provides fine fuels and increases fire continuity (Germino and others, 2016). As an annual species, its rapid growth under favorable conditions creates fine fuel buildup necessary for shorter fire return intervals (Pilliod and others, 2017). Shortened fire return intervals do not allow sagebrush time to reestablish (Simler-Williamson and others, 2022).
Historically practices such as controlled burns, mechanized thinning, and targeted grazing have been used to lower fuel loads and decrease ignition risk (Ellsworth and others, 2022). However, as fire frequency and severity increase, research is now trying to understand the potential future utility of these practices (Shinneman and others, 2023). The question of treatment efficacy is further compounded by public resistance to the use of herbicides and the costs of mechanical interventions as fire frequency increases (Brunson and Shindler, 2004; Gordon and others, 2014). One potential fire management practice is targeted grazing, which could require flexible use grazing permits to match grazing periods to annual growing seasons (Davies and others, 2022; Gornish and others, 2023). Complications with public resistance to historical practices and elevated fire activity has increased the need for effective postfire restoration strategies in big sagebrush plant communities (Brabec and others, 2017; Applestein and others, 2021; Ellsworth and others, 2022).
There is a consensus that climate change will exacerbate fire dynamics already altered by invasive annual grasses in the sagebrush region (Abatzoglou and Kolden, 2011; Chambers and others, 2014). Further, high interannual weather variability in postfire conditions can affect native plant establishment and will likely make restoration more challenging (Shriver and others, 2018). Effective wildfire management will need to consider the threat of invasive annual grasses, the cost of fuel treatments, and the ability of targeted grazing to lower fuel loads.
Finally, evidence suggests that there is a high incidence of human-caused wildfire ignitions in the Intermountain West (Bradley and others, 2018). Public education on the consequences of fire in sagebrush vegetation may have decrease wildfire incidences.
2.3. Habitat Management—Restoration Targets and Wildlife
Section 2.3 Highlights
● Habitat fragmentation is likely because of increasing abundance of invasive annual grasses and conifers and potential declines in big sagebrush and perennial grasses.
● Combating fragmentation may require successful techniques for postfire vegetation restoration and active treatments to reduce the abundance of invasive annual grasses and conifers.
Many of the preceding sections established the important link between climate and big sagebrush plant communities. This section discusses how climate-induced shifts in vegetation may alter wildlife habitat and behavior and how wildlife habitat management strategies may be adapted to maintain sagebrush habitat. This section reviews how shifts in vegetation are likely to affect habitat for wildlife and how interactions with conifer expansion and invasive annual grasses are likely to affect restoration success.
2.3.1. Maintaining Connectivity and Wildlife Habitat
As climate change reshapes the potential distribution of sagebrush, the distribution and abundance of wildlife habitat provided by intact big sagebrush plant communities is also expected to shift (Palmquist and others, 2021; Rigge, Shi, and Postma, 2021; Havrilla and others, 2023). This section focuses on three aspects of wildlife habitat that will likely be affected by the expected shifts in sagebrush vegetation: habitat fragmentation, migration routes, and habitat restoration.
Habitat fragmentation because of climate-driven shifts in vegetation is not limited to sagebrush ecosystems (Mantyka-pringle and others, 2012). In widely distributed vegetation types, changes across elevation gradients may isolate some communities as they move upslope and are replaced by other species at lower elevations (Hsiung and others, 2018). This fragmentation has the potential to alter wildlife movement, disrupt gene flow, and constrain species' adaptive capacity (Andrén, 1994; Fahrig, 2003). For example, the risk of isolating high-elevation big sagebrush habitats raises concerns about diminished connectivity that can impede wildlife movement and hinder their capacity to cope with interannual variability in forage availability (Love and others, 2023). To mitigate these challenges, managers may consider prioritizing conservation and restoration of corridors between areas of high-quality sagebrush habitat (Chambers, Maestas, and others, 2017).
Increasing connectivity between areas of intact sagebrush plant communities will also serve to mitigate the effects of shifts in the timing of soil moisture availability and plant growth because of changes in phenology associated with altered precipitation. These shifts in the timing and productivity of forage may have a negative effect on wildlife species, particularly ungulate game species, reliant on these ecosystems during migration (Merkle and others, 2016; Aikens and others, 2017; Bloom and others, 2022). The temporal shifts in soil moisture availability and plant growth could disrupt wildlife migration seasonal timing and potentially routes. Beyond changes to vegetation, increases in temperature are also likely to affect animal movement, but these animal effects are outside the focus of this synthesis.
In the context of climate-induced changes in wildlife habitat connectivity and shifts in the phenology of plant species, avoiding wildlife population declines may depend on effective habitat restoration (Davies and others, 2011; Chambers, Beck, and others, 2017). As areas of sagebrush are lost because of climate-induced drought and (or) wildfires, resource managers may consider implementing climate resilient restoration strategies. Techniques such as reseeding with locally adapted subspecies and (or) genotypes of big sagebrush, preventing invasive plant establishment, and adopting adaptive land management practices that stimulate sagebrush recovery (such as seeding with perennial grasses across multiple years) are potential options to improve conditions for wildlife (Brabec and others, 2017; Germino and others, 2018; Applestein and others, 2021).
2.3.2. Management to Minimize Invasive Annual Grasses
Invasive annual grasses have already affected large parts of the sagebrush region because of their ability to exploit a broad ecological niche and show resilience to changing climate conditions. To maintain sagebrush habitat for wildlife, managers may use monitoring data that can serve as an early detection system for invasive annual grass species (Bradley, 2009; Chambers, Maestas, and others, 2017). The BLM monitors vegetation through the AIM Strategy across the lands it manages. Managers may want to recognize the value that AIM sampling provides for understanding presence and absence of invasive plant species. As AIM sampling continues, it can provide information about how invasive plant abundance is changing across time. In addition, satellite-based imaging products such as the U.S. Department of Agriculture Rangeland Analysis Platform and the US Geological Survey Rangeland Condition Monitoring Assessment and Projection (RCMAP) may be useful to detect abundance trends for invasive annual grasses (Jones and others, 2018; Rigge and others, 2023). By quickly identifying the presence of these invasive species, swift and effective responses, including targeted herbicide applications, manual removal efforts, or prescribed burns, can be aimed at preventing the establishment and proliferation of these invasive species (Germino and others, 2016; Shinneman and others, 2023).
2.3.3. Managing Conifer Expansion in the Context of Climate Change
Since at least the 1990s, coniferous tree species, such as pinyon pine and juniper, have been expanding into sagebrush ecosystems (Maestas and others, 2021). Although this expansion has negative effects for sagebrush habitat and wildlife populations (Doherty and others, 2022), ongoing and predicted shifts in precipitation patterns may continue to support conifer expansion and potentially reduce the long-term effectiveness of conifer-reduction management strategies (Zimmer and others, 2021).
Proactive vegetation management strategies, such as selective thinning or the targeted removal of expanding conifers, are used to maintain a balance between sagebrush and coniferous trees (Davies and Bates, 2019). This approach preserves habitat for sagebrush-dependent wildlife species while effectively countering the expansion of conifers (Davies and Bates, 2014). However, expected expansion of conifers and the expected increase in precipitation may make proactive efforts to limit conifer expansion more costly as conifers expand at an increased rate (Maestas and others, 2021).
2.4. Potential Climate Change Effects on Recreation in Big Sagebrush Plant Communities
Section 2.4 Highlights
● Managers may consider fire restrictions and (or) seasonal closures to mitigate wildfire risk in the context of growing recreational pressure.
● Modifications to recreation permits may be needed as warmer conditions alter seasonal recreation patterns, particularly in areas where infrastructure that supports recreation is threatened by enhanced wildfire risk.
● Recreation is an opportunity for natural resource managers to connect with the public, providing information on approaches to resource management in a time of rapid change.
Climate change will have ramifications for the recreational uses of big sagebrush plant communities. Direct impacts to recreation from extreme climate events and wildfires were reviewed by Brice and others (2020). This section focuses specifically on the impacts that climate-induced changes to big sagebrush plant communities may have for recreation. These impacts include increased wildfire risks to altered recreational seasons and activities, infrastructure and facilities vulnerabilities, and opportunities for enhanced educational programs.
2.4.1. Increased Wildfire Risks
The increased frequency and severity of wildfires because of climate change poses risks to recreation activities across the sagebrush region, including smoke inhalation and wildfire risk for public recreators. These risks are compounded during periods of high interannual variability that lead to fine fuel buildup and consequently more intense wildfires (Pilliod and others, 2017). In areas that may have cheatgrass expansion, wildfire risks are likely to increase. Activities such as hiking and camping will be particularly affected, as wildfire may temporarily restrict access (Brice and others, 2020). Recreational decisions can proactively mitigate wildfire risks by prioritizing visitor safety, for example, through the use of fire restrictions and seasonal closures.
2.4.2. Altered Recreational Seasons and Activities
Climate-induced shifts in vegetation and altered wildfire regimes are also likely to impact the timing of forage availability and habitat quality for wildlife (Abatzoglou and Kolden, 2011; Coates and others, 2016; Aikens and others, 2017). The shifts in temperature and precipitation patterns because of climate change can affect the timing of natural occurrences and potentially affect recreational activities such as birdwatching, off-road vehicle use, hunting, and shed antler and horn collection (Munson and Long, 2017; Bloom and others, 2022). For example, phenological shifts may alter bird migratory behavior and affect the seasonal timing of birdwatching (Munson and Long, 2017). The timing of mammal migrations may also change because migrating populations alter behavior to match forage availability (Aikens and others, 2017). Thus, hunting and antler collecting seasons may need to be flexible.
2.4.3. Infrastructure and Facilities Vulnerability
The intensification of extreme weather and increased erosion rates pose risks to infrastructure and recreational facilities on BLM lands that can potentially lead to disruptions in recreational access. Infrastructure and facility vulnerability is especially high in areas like the Great Basin where cheatgrass has altered the fire cycle and in regions where expected declines in perennial herbaceous cover will increase the risk of erosion (Bradley and others, 2018; Edwards and others, 2019; Shi and others, 2020). Trails, roads, campsites, and visitor centers may be vulnerable to damage, which emphasizes the need for resilient infrastructure. Climate adaptive fire management practices (manual fine fuel reduction or targeted grazing) can reduce risk to infrastructure, and restoration techniques (reseeding) that increase perennial herbaceous cover can reduce erosion risk (Chambers, Beck, and others, 2017; Shinneman and others, 2023).
2.4.4. Opportunities for Enhanced Educational and Interpretive Programs
Shifts in big sagebrush vegetation because of climate change provide an opportunity to enhance climate change related educational and interpretive programs. Engaging individuals on the topic of climate change by highlighting visible changes to the resources they care about was identified as a positive climate change education strategy (Monroe and others, 2019). Developing informative signs along trails and displays in visitor centers that highlight the ecological changes from climate change can support education efforts.
2.5. Reclamation-Based Uses and Intensive-Use Projects
Section 2.5 Highlights
● The challenge of reclamation following fossil fuel extraction or other intensive land development activities in sagebrush environments is likely to increase as temperatures rise and further limit restoration potential.
● Recognizing geographic patterns of wildlife habitat use may be increasingly helpful for selecting intensive use sites that minimize the consequences of land development for habitat connectivity
● Similarly, recognizing geographic variation in ecological resilience may help identify intensive use sites with maximum potential for successful postdevelopment reclamation success.
2.5.1. Vegetation Changes and Restoration Targets
Ecological resilience of big sagebrush ecosystems is a measure of their ability to recover after disturbances, including development (Chambers, Maestas, and others, 2017). Climate change may reduce ecological resilience in the big sagebrush region (Bradford and others, 2019). Reductions in resistance or resilience may decrease the success of reclamation treatments and suggest that adjustments to expected vegetation composition and cover may be necessary (Chambers, Maestas, and others, 2017). As resilience shifts, the BLM may consider setting targets for reclamation that prevent invasive annual grass establishment, and managers may consider utilizing climate-adapted seed sources to accomplish this (Brabec and others, 2017), applying repeated restoration treatment that enhance the likelihood of successful plant establishment (Shriver and others, 2018), and using assisted migration of plant species to foster the establishment of climatically viable perennial herbaceous species (Havrilla and others, 2023).
2.5.2. Development Site Selection
Shifts in the resistance and resilience of sagebrush communities are also likely to affect decisions about where surface disturbances may occur (Chambers, Beck, and others, 2017). The heightened potential for more frequent and intense wildfires means that assessing shifts in wildfire vulnerability will be important before any land development (Brice and others, 2020). For example, when possible, well-pad placement could also integrate considerations of nearby sagebrush habitats and wildlife populations, particularly near migration corridors and lekking areas (Chambers, Beck, and others, 2017). Site selection may need to avoid intact communities with projected decreases in resistance and resilience caused by increased temperature and lower precipitation (Palmquist and others, 2021; Zimmer and others, 2021; Holdrege and others, 2024a).
2.5.3. Intensive Use Projects
High-intensity use projects, such as solar energy development, are a growing consideration for the BLM. Expected shifts in sagebrush habitat and the changes to resistance and resilience across the region may act as guides for these projects (Chambers, Maestas, and others, 2017; Palmquist and others, 2021). Climate-induced shifts that result in reduced sagebrush cover and density can put a higher value on remaining sagebrush patches where development could be avoided (Chambers, Beck, and others, 2017). The development of intensive projects, such as solar fields, can amplify these effects by fragmenting or removing critical habitats. As such, development may need to avoid important breeding areas for wildlife populations and migration corridors, particularly for species of concern like greater sage grouse and ungulate game species. Finally, the loss and fragmentation of sagebrush habitats because of high-intensity use projects may lower the resilience and connectivity of remaining sagebrush patches.
3. Frameworks, Tools, Datasets, and Analysis Approaches for Understanding Climate Change Impacts on Big Sagebrush Plant Communities
3.1. Climate Adaptation Conceptual Frameworks
Section 3.1 Highlights
● This section briefly introduces conceptual frameworks, tools, datasets, and analytical approaches designed to help natural resource managers identify strategies and tactics for climate adaptation and includes links to learn more about the materials and resources.
As climate change progresses, resource management, including management actions in the sagebrush region, will increasingly need to adapt to the effects of higher temperatures, shifting precipitation patterns, and altered disturbance regimes on plant community composition and structure. Climate adaptation in natural resource management involves adjusting strategies and goals to sustain ecosystems and ecosystem services in the face of changing climate conditions (Stein and others, 2014). This shift from basing natural resource management entirely on historical conditions, variability, and knowledge to explicitly recognizing long-term directional change and anticipating future conditions is a substantial paradigm shift and a critical contemporary challenge (Kemp and others, 2015; Bradford and others, 2018). To help clarify how to approach the paradigm shift represented by climate adaptation, conceptual frameworks for understanding and applying climate adaptation in natural resource management have emerged after roughly 2015 (for example, Peterson St-Laurent and others, 2021; Schuurman and others, 2022).
The Resist-Accept-Direct (RAD) framework is a conceptual tool for helping decisionmakers recognize possible strategies and approaches for climate adaptation (Schuurman and others, 2020, 2022; Lynch and others, 2022). The RAD framework was developed by an interagency effort led by the National Park Service starting in 2015 and is now one of the most well-recognized climate adaptation frameworks. By recognizing the growing effects of climate change on ecosystems, the RAD framework provides guidance and tools for making decisions about resisting climate effects (maintaining or restoring historical ecosystem conditions), accepting climate effects (allowing an ecosystem to change without intervention), or directing climate effects (actively shaping ecosystems toward a preferred new condition). Additional information about the RAD framework, including links to several publications developing the RAD conceptual foundations and a growing array of decision support tools, is available at https://www.nps.gov/subjects/climatechange/resistacceptdirect.htm.
The Climate Change Response Framework (CCRF) is another information source for helping natural resource managers approach climate adaptation. The CCRF is led by the U.S. Department of Agriculture Forest Service’s Northern Institute for Applied Climate Science and has developed a variety of decision support tools for guiding decisionmakers through the process of integrating climate change into natural resource management. More information, including links to multiple tools, is available at https://forestadaptation.org.
Although these conceptual frameworks can be helpful for integrating climate adaptation into vegetation management, their applications specific to management of big sagebrush plant communities have, to date, been relatively limited. Within the RAD framework, accepting or directing change was suggested in some parts of the sagebrush region dominated by invasive annuals (Davies and others, 2021) and may be the appropriate strategy for handling conifer expansion (Van Lanen and others, 2023). Where resistance may be more feasible, research indicates that restoration efforts that target adaptive seeding are most successful for re-establishing sagebrush (Brabec and others, 2017). Although specific approaches to climate adaptation will vary across the sagebrush region, conceptual frameworks such as the RAD framework and the CCRF can help managers make informed decisions when facing uncertainty because of climate change. For example, Holdrege and others (2024a) identified core sagebrush areas where resisting climate-driven degradation is more likely to be successful and contrasted to those other areas where accepting or directing ecological change may be necessary, because they already have lower ecological integrity and are projected to become less climatically suitable in the future.
3.2. Tools for Interactive Visualization of Projected Future Climate Conditions
Users can access and visualize climate data on several websites for their particular region of interest. The following are some example websites and applications:
-
● Climate Engine—The application includes past weather data and near-term weather forecasts. Users can create and download maps or time series of datasets with climate and hydrological variables, remotely sensed data products, hazards, and near-term meteorological and hydrological forecasts for points, polygons, or the conterminous United States (https://app.climateengine.org/).
-
● The Climate Toolbox—The website includes past data, near-term forecasts, and long-term future climate projections. Users can create and download maps, time series, and other dataset visualizations using a large set of tools for historical data, near-term and seasonal forecasts, and future projections for points, polygons, or the conterminous United States (https://climatetoolbox.org/).
-
● IPCC Working Group I Interactive Atlas—The website includes historical data and future projections. Users can create maps and visualizations of past trends, projections, and uncertainty for climate impact drivers for the globe or specific geographic areas (https://interactive-atlas.ipcc.ch/).
3.3. Analytical Approaches to Assess Potential Long-Term Climate Change Effects on Plant Communities
To help resource managers recognize the potential value and appropriateness of datasets for their decisions, this report contrasts the strengths and weaknesses among research approaches to estimate potential climate change effects on big sagebrush plant communities. Two approaches have been used most often to understand the climate change effects on big sagebrush ecosystems: statistical models and process-based models.
Studies that rely on a correlation-based approach, through the use of various statistical modeling methods represent current or historical relationships between climate and vegetation based on either field or remotely sensed data. These empirical studies use relationships constructed from current observations and extrapolate them across parts or all the sagebrush region and into the future. A common approach is to use a model to estimate the relationships between climate and vegetation from a particular dataset and then estimates of potential future vegetation can be made by inputting future climate data from global climate model projections (for example, Rigge, Shi, and Postma, 2021). As a hypothetical example, a model based on spatial correlations could represent a positive relationship between mean annual precipitation and total plant cover, if in the dataset used, wetter areas tend to have more plant cover. This model would then predict increases in future plant cover in areas where climate projections indicate mean annual precipitation will increase. A limitation with these types of statistical models is that they may capture correlations between variables that are not causal. This limitation can be problematic when using the model to make predictions for novel future climate conditions because correlations among climate variables may be different than they were historically.
Process-based models represent physical (for example, soil moisture) and biological (for example, competition between neighboring plants) processes that govern plant establishment, growth, and mortality. They use projections from global climate models and fundamental relationships to project plant and ecosystem responses to climate change. For example, a process-based model may estimate soil moisture across time based on precipitation, temperature, and principles of soil physics. The model can use that information to estimate the amount of plant functional type biomass that the estimated amount of soil water can support across a growing season or years (Palmquist and others, 2021).
An important limitation to consider when using results from either a statistical or a process-based model is that direct validation of future projections are impossible. In the absence of direct model validation, a good approach is to compare or aggregate predictions from multiple models. Renwick and others (2018) found that when estimating sagebrush responses to climate, results were fairly consistent among independent modeling approaches. However, uncertainty among models was still greater than uncertainty from different climate change scenarios. Therefore, a good way to increase confidence in projections and better understand the uncertainty in potential climate effects on plant communities is to assess agreement among studies that use different modeling approaches and rely on independent data (Zimmer and others, 2021).
3.4. Available Datasets Relevant to Climate Change Impacts on Big Sagebrush Plant Communities
This section summarizes several currently (2024) available datasets describing potential climate change impacts on vegetation in the sagebrush region. This section focuses on research that has resulted in peer-reviewed, published, and publicly available spatial datasets that cover parts of the sagebrush region. Experimental or observational datasets that have limited spatial coverage or that do not provide estimates of change in response to climate change were not included. What follows are brief descriptions of spatial datasets that may be useful for individuals seeking estimates of the climate change effects on plant communities across large parts of the sagebrush region. When possible, it may be preferable to use results from multiple datasets instead of one dataset to increase robustness of analyses and subsequent decision making.
3.4.1. Holdrege and others (2024)—Remote Sensing Estimates of Ecological Integrity Combined with Process-Based Modelling Projections of Change
Overview.—Holdrege and others (2024a) used estimates of sagebrush ecological integrity (SEI; from Doherty and others, 2022), and combined these estimates with projections of vegetation change to create projections of SEI under climate change. STEPWAT2, a process-based simulation model, was used to estimate changes in sagebrush, perennial grasses and forbs, and annual grasses and forbs, in response to climate change. These projected changes were then used to calculate a modified SEI that was a projection of potential future SEI under climate change. This dataset can help users assess whether areas that had high ecological integrity from 2017 to 2020 are projected to experience conditions that will be favorable for maintaining that ecological integrity, or whether climatic suitability is likely to decline in the future. Doherty and others (2022) created layer of projected future SEI classes, using similar methods as Holdrege and others (2024a). Individuals wishing to use climate change projections of SEI may want to use the layers from Holdrege and others (2024a), because they represent an update of the work started by Doherty and others (2022) and are different in that they (1) used a slightly updated version of the STEPWAT2, (2) ran simulations for multiple climate scenarios, and (3) ran simulations for multiple ecological assumptions.
Pros.—This dataset directly provides information on which areas had high SEI historically and are likely to be climatically suitable to maintain high SEI in the future, which may be useful for prioritizing management efforts. Many climate scenarios were used, allowing for a fairly robust accounting of climate uncertainty.
Cons.—The greatest source of uncertainty in SEI projections is likely the uncertainty in STEPWAT2 simulation output, therefore these results have similar limitations as Palmquist and others (2021). Additionally, projections of SEI assume cover of conifers, and the amount of human modification on the landscape, will remain fixed because projections of these factors were not available. Remotely sensed estimates of vegetation cover used for calculating historical SEI also contain errors.
Article citation.—Holdrege, M.C., Palmquist, K.A., Schlaepfer, D.R., Lauenroth, W.K., Boyd, C.S., Creutzburg, M.K., Crist, M.R., Doherty, K.E., Remington, T.E., Tull, J.C., Wiechman, L.A., and Bradford, J.B., 2024, Climate change amplifies ongoing declines in sagebrush ecological integrity: Rangeland Ecology & Management, v. 97, p. 25–40, https://doi.org/10.1016/j.rama.2024.08.003.
Dataset citation.—Holdrege, M.C., Schlaepfer, D.R., Palmquist, K.A., Theobald, D.M., and Bradford, J.B., 2024, Current and projected sagebrush ecological integrity across the Western U.S., 2017–2100: U.S. Geological Survey data release, https://doi.org/10.5066/P13RXYZJ.
Format.—Rasters (GeoTIFF).
Resolution.—90 meter (m).
Extent.—The sagebrush region.
Climate Scenarios.—RCP4.5 and RCP8.5 from 2031 to 2060 and from 2071 to 2100 (median across 13 GCMs, as well as high and low estimates provided).
Variables.—SEI (continuous variable, ranging between 0 and 1), SEI classes (core sagebrush areas, growth opportunity areas, other rangeland areas), and Q (“quality”) scores (continuous variable, ranging between 0 and 1) for sagebrush, perennial forbs and grasses, and annual forbs and grasses.
3.4.2. Tredennick and others (2023)—Statistical Models Relating Sagebrush Cover and Climate
Overview.—Tredennick and others (2023a) created correlation-based models (largely relying on time-series) relating sagebrush cover (from the RCMAP products) to climate. They used two climate variables as predictors (spring and summer precipitation, and spring and summer temperature), while also accounting for sagebrush cover in the previous year and surrounding grid-cells. Their study area was restricted to sage-grouse core areas in Wyoming, and they fit separate models to each core area. For a given sage-grouse core area, climate and sagebrush cover data from each year from 1985 to 2018 in 100-m grid-cells were used to fit a regression model. Future climate conditions were then input into the model to predict future sagebrush cover. The datasets include spatial projections of sagebrush cover for individual sage-grouse core areas for multiple periods and emissions scenarios.
Pros.—The authors chose only two climate metrics, and fit simple relationships (log-linear) in the model. Although they may be missing some more complex relationships (for example, other climate variables and their interactions), their approach is less likely to turn up spurious relationships, and the results have a high resolution.
Cons.—Because separate models were fit to each sagebrush core area in Wyoming, each model is based on data from a small area, and as such only includes a small amount of spatial climate variation. Most of the climate variability included in the model is from year-to-year variation. Therefore, sagebrush cover predictions under future climate conditions are likely extrapolating beyond climate conditions used to fit the model. Additionally, because separate models were fit to small areas, any given model may be partially capturing artifacts of a given location instead of broad underlying trends for this part of the sagebrush region. The evidence for this possible limitation is that models for different sage-grouse core areas show a range of responses including negative and positive temperature and precipitation responses.
Article citation.—Tredennick, A.T., Monroe, A.P., Prebyl, T., Lombardi, J., and Aldridge, C.L., 2023, Dynamic spatiotemporal modeling of a habitat‐defining plant species to support wildlife management at regional scales: Ecosphere, v. 14, no. 6, 20 p., https://doi.org/10.1002/ecs2.4534.
Dataset citation.—Tredennick, A.T., Monroe, A.P., Prebyl, T., Lombardi, J., and Aldridge, C.L., 2023, Sagebrush projections for greater sage-grouse core areas in Wyoming, USA, 2018–2100: U.S. Geological Survey data release, https://doi.org/10.5066/P9G58V48.
Format.—Network Common Data Form (netCDF) files.
Resolution.—100 m.
Extent.—Parts of Wyoming (data used was from greater sage-grouse core areas, which are spread across the State). Refer to the article for a map of the core areas.
Climate Scenarios.—Emissions pathways were SSP126, SSP245, and SSP585 and periods were from 2018 to 2045, from 2046 to 2070, and from 2071 to 2100. The results provided are weighted averages of 18 GCMs.
Variables.—Sagebrush cover.
3.4.3. Doherty and others (2022)—Remote Sensing Estimates of Ecological Integrity Combined with Process-Based Modelling Projections of Change
Overview.—Doherty and others (2022) used remotely sensed data to calculate SEI. This calculated SEI was used to create a dataset of SEI classes where pixels across the sagebrush region were categorized as “belonging to core sagebrush areas,” “growth opportunity areas,” or “other rangeland areas.” High sagebrush ecological integrity (designated as “core sagebrush areas”) was defined as areas with an adequate cover of sagebrush and perennial grasses and forbs and no or limited annual grasses and forbs, trees, and human modification of the landscape. Data on projected relative changes in sagebrush, perennial grasses and forbs, and annual grasses and forbs in response to climate change (from Palmquist and others, 2021), were used to calculate a modified SEI that was a projection of potential future SEI under climate change. This projected SEI was used to classify areas in the future (under a single climate scenario) as “core sagebrush areas,” “growth opportunity areas,” or “other rangeland areas.” This dataset can help users assess whether areas that had high ecological integrity from 2017 to 2020 are projected to experience conditions that will be favorable for maintaining that ecological integrity, or whether climatic suitability is likely to decline under future climate conditions. Holdrege and others (2024a) also created projections of future SEI using the newer (projected) dataset that may be a more appropriate representation of SEI because they represent an extensive update of the projections made by Doherty and others (2024).
Pros.—This dataset combines remotely sensed estimates of ecological integrity with estimates of potential changes in sagebrush plant communities in response to climate change. Thereby, this dataset directly provides information on how those core sagebrush areas might respond to climate change.
Cons.—The data layer providing projections of SEI classes incorporates STEPWAT2 simulation results, and therefore it has similar limitations as Palmquist and others (2021). Additionally, the analysis was only done using the median vegetation response (across GCMs) under a single climate scenario and period, so uncertainty in climate change is not considered. Remotely sensed estimates of vegetation cover used for calculating historical SEI also contain errors.
Article citation.—Doherty, K., Theobald, D.M., Bradford, J.B., Wiechman, L.A., Bedrosian, G., Boyd, C.S., Cahill, M., Coates, P.S., Creutzburg, M.K., Crist, M.R., Finn, S.P., Kumar, A.V., Littlefield, C.E., Maestas, J.D., Prentice, K.L., Prochazka, B.G., Remington, T.E., Sparklin, W.D., Tull, J.C., Wurtzebach, Z., and Zeller, K.A., 2022, A sagebrush conservation design to proactively restore America’s sagebrush biome: U.S. Geological Survey Open-File Report 2022–1081, 38 p., https://doi.org/10.3133/ofr20221081.
Dataset citation.—Doherty, K., Theobald, D.M., Holdrege, M.C., Wiechman, L.A., and Bradford, J.B., 2022, Biome-wide sagebrush core habitat and growth areas estimated from a threat-based conservation design: U.S. Geological Survey data release, https://doi.org/10.5066/P94Y5CDV.
Format.—Rasters (GeoTIFF).
Resolution.—30 m.
Extent.—The sagebrush region.
Climate Scenarios.—RCP8.5 from 2030 to 2060 (median across 13 GCMs provided).
Variables.—Layers provide SEI classes (pixels fall in one of three categories: core sagebrush areas, growth opportunity areas, and other rangeland areas). Five layers provide SEI classes for one of five historical periods (between 1998 and 2020), and one layer provides projected future SEI classes under climate change.
3.4.4.Palmquist and others (2021)—Process-Based Modeling of Plant Functional Group Responses to Climate Change
Overview.—Palmquist and others (2021) used an individual plant-based simulation model (STEPWAT2) to simulate growth of plant functional types in sagebrush ecosystems, including big sagebrush, C3 and C4 perennial grasses, cheatgrass, and perennial forbs. The STEPWAT2 model simulates plant growth based on the amount of available water that is estimated in the soil (which is based on temperature, precipitation, and soil type). They simulated sagebrush plant communities at 200 sites, and these results were then extrapolated across the sagebrush region. Simulations were conducted under current climate conditions, for multiple emissions scenarios, periods, and GCMs to ensure that uncertainty induced by different climate scenarios was well represented.
Pros.—The model was designed for dryland ecosystems and specifically for use in sagebrush ecosystems. Because it is process-based, results may be more robust under future (potentially novel) climate conditions. These results represent one of the most extensive process-based modeling efforts of plant functional type specific responses to climate change across the sagebrush region.
Cons.—STEPWAT2 does not incorporate the land-use history of a given location, or the actual vegetation currently observed there. Therefore, output is most appropriately viewed as a rough estimate of the potential amount of biomass of each plant functional type that the climate of a given location can support, and how that might change with a changing climate. In addition, STEPWAT2 results have been validated only in limited context, primarily with spatial comparisons. Thus, the confidence in STEPWAT2 estimates of impacts from long-term climatic shifts remains difficult to assess.
Article citation.—Palmquist, K.A., Schlaepfer, D.R., Renne, R.R., Torbit, S.C., Doherty, K.E., Remington, T.E., Watson, G., Bradford, J.B., and Lauenroth, W.K., 2021, Divergent climate change effects on widespread dryland plant communities driven by climatic and ecohydrological gradients: Global Change Biology, v. 27, no. 20, p. 5169–5185, https://doi.org/10.1111/gcb.15776.
Dataset citation.—Palmquist, K.A., Renne, R.R., Schlaepfer, D.R., Lauenroth, W.K., and Bradford, J.B., 2022, High-resolution maps of projected big sagebrush plant community biomass for 52 future climate scenarios using multivariate matching algorithms: U.S. Geological Survey data release, https://doi.org/10.5066/P9DR9G1Y.
Format.—Rasters (GeoTIFF).
Resolution.—30 arcsecond (less than 1 kilometer [km]).
Extent.—The sagebrush region.
Climate Scenarios.—RCP4.5 and RCP8.5 from 2030 to 2060, and from 2070 to 2010 (13 GCMs per RCP and period).
Variables.—Aboveground biomass (grams per square meter) of big sagebrush, C3 and C4 perennial grasses, cheatgrass, and perennials forbs.
3.4.5. Rigge, Shi, and Postma (2021)—Statistical Modeling Approach Used to Relate Plant Cover to Climate and Other Factors
Overview.—Rigge, Shi, and Postma (2021) used a correlation approach to model vegetation cover as a function of soil, topography, and climate variables. They used generalized additive models, which allowed for fitting flexible (nonlinear) relations. Subsequently, Rigge and others (2023) used the same approach except they used a deep neural network model. They separately modeled shrub, sagebrush, herbaceous, and annual herbaceous cover using data from a random subset of pixels across the sagebrush region each year from 1985 to 2018. Therefore, the relationships captured by their models represent a combination of spatial and temporal patterns found in the data. The vegetation cover values used as the response variables in the models are estimates based on Landsat satellite data. They used the models to predict future cover by inputting future climate conditions from two emissions scenarios (RCP4.5 and RCP8.5) and three periods. Note that the older version of this dataset, which corresponds to the Rigge, Shi, and Postma (2021) manuscript are available at https://doi.org/10.5066/P9EC2094, however, they recommend using the newer version (https://doi.org/10.5066/P9J490BH).
Pros.—The models Rigge, Shi, and Postma(2021) fit represented cover under current conditions at a high resolution. This dataset provides some of the most complete projections (with regard to spatial coverage and plant functional types modeled) available from a correlation-based model in the sagebrush region.
Cons.—Correlational models may not extrapolate well to novel conditions because the relations represented in the model are not necessarily causal. However, Rigge and others (2023) contend that because of the large climate envelope covered by the sagebrush region, most future climate conditions they assessed included few truly novel conditions. Rigge, Shi, and Postma (2021) noted that in the southern Great Basin and southern Colorado Plateau the model projected increases in sagebrush cover in response to climate change, which they suspect is model error. However, Rigge, Shi, and Postma (2023) state that the new estimates have fewer errors.
Article citation.—Rigge, M., Shi, H., and Postma, K., 2021, Projected change in rangeland fractional component cover across the sagebrush biome under climate change through 2085: Ecosphere, v. 12, no. 6, art. e03538, 25 p., https://doi.org/10.1002/ecs2.3538.
Dataset citation.—Rigge, M., Postma, K., Bunde, B., and Shi, H., 2023, Projections of rangeland fractional component cover across Western US rangelands for RCP4.5 and RCP8.5 scenarios for the 2020s, 2050s, and 2080s time-periods: U.S. Geological Survey data release, https://doi.org/10.5066/P9J490BH.
Format.—Rasters (GeoTIFF).
Resolution.—30 m.
Extent.—Rangelands across the Western United States.
Climate Scenarios.—RCP4.5 and RCP8.5 for 2020s, 2050s, and 2080s (results are based on the median output of 15 GCMs).
Variables.—Cover of shrubs, sagebrush, herbaceous plants, annual herbaceous plants, litter, and bare ground.
3.4.6. Schlaepfer and others (2021)—Process-Based and Statistical Models Used to Estimate Future Sagebrush Regeneration
Overview.—Schlaepfer and others (2021) used two different models to estimate future regeneration probability of sagebrush. They used a process-based germination and individual seedling survival model (GISSM), which relies on daily weather and ecohydrological variables (including various soil moisture metrics), to estimate the potential regeneration probability of big sagebrush under natural conditions (in other words, under conditions with no disturbance, no invasion by annual grasses, and so forth). They used a second, correlation-based (regression) model (Shriver and others, 2018) to estimate the probability that restoration succeeds following fire and seeding (based on measurements made across multiple wildfire sites). The Shriver and others (2018) regression model estimates this probability based on two variables (mean temperature and spring soil moisture). Projections of regeneration probability (GISSM) and probability of restoration success (Shriver and others, 2018) were made under multiple climate scenarios and time periods.
Pros.—These results may present the most complete, spatially explicit, modeling results of big sagebrush regeneration responses to climate change.
Cons.—Both models are subject to the limitations that all models of the future share—there is not a way to directly test their predictions. Additionally, they assume the availability of big sagebrush seeds, which may or may not be reasonable depending on the history of the particular location.
Article citation.—Schlaepfer, D.R., Bradford, J.B., Lauenroth, W.K., and Shriver, R.K., 2021, Understanding the future of big sagebrush regeneration—Challenges of projecting complex ecological processes: Ecosphere, v. 12, no. 8, art. e03695, 26 p., https://doi.org/10.1002/ecs2.3695.
Dataset citation.—Schlaepfer, D.R., and Bradford, J.B., 2021, Simulated rangewide big sagebrush regeneration estimates and relationships with abiotic variables as function of soils under historical and future climate projections: U.S. Geological Survey data release, https://doi.org/10.5066/P9MB2QB8.
Format.—netCDF.
Resolution.—10 km.
Extent.—The projections from the Schriver2018 model are restricted to the central and northern Great Basin and Snake River Plain (because data used to fit that model were only from that area). The projections from the GISSM model cover the entire sagebrush region.
Climate Scenarios.—Projections were made for two scenarios (RCP4.5 and RCP8.5) and periods (from 2020 to 2050 and from 2070 to 2099). Median projected values across GCMs were provided.
Variables.—Regeneration probability (GISSM) and probability of restoration success (Schriver2018) of big sagebrush.
3.4.7. Zimmer and others (2021)—Multi-Model Comparison of Vegetation Responses to Climate Change
Overview.—Zimmer and others (2021) combined results from studies that were available before 2020 that created spatial estimates (through correlation or process-based modeling) of vegetation responses to climate change. They compiled results for studies done within the sagebrush region and estimated responses of cheatgrass (3 studies), forage production (3 studies), pinyon-juniper (5 studies), or sagebrush (3 studies) to climate change. They digitized results from figures in published articles and therefore did not have to rely only on studies that had also published their data. By using this approach, they were able to incorporate results from some studies that are not individually listed in the “Available Datasets Relevant to Climate Change Impacts on Big Sagebrush Plant Communities” section of this report. The final dataset Zimmer and others (2021) created includes rasters that count the number of studies projecting positive or negative climate responses in a given pixel for each of the vegetation components they considered. Since Zimmer and others (2021) completed their work, some additional datasets have become available (for example, Palmquist and others, 2021; Rigge, Shi, and Postma, 2021); therefore, their study is not a complete account of all available datasets projecting vegetation responses to climate change in the sagebrush region.
Pros.—Zimmer and others (2021) compared the results of multiple studies that used different methods and approaches. This method may be the best way to assess the level of confidence in the direction of projected change because there are many sources of uncertainty in estimates of sagebrush ecosystem responses to climate change.
Cons.—Zimmer and others (2021) combined results from studies that modeled different response variables that have different units (for example, cover, presence, and net primary productivity). Therefore, the magnitude of responses could not readily be combined, and the data product they created is only able to provide information on confidence in the direction of climate responses (in other words, the number of studies projecting positive or negative responses). As such, this dataset would not be directly useful if an analyst wants to have a continuous variable (for example, change in cover) that estimates magnitude of climate change response. The resolution may also be too coarse for applications requiring spatial resolution greater than about 15 square kilometers (km2).
Article citation.—Zimmer, S.N., Grosklos, G.J., Belmont, P., and Adler, P.B., 2021, Agreement and uncertainty among climate change impact models—A synthesis of sagebrush steppe vegetation projections: Rangeland Ecology & Management, v. 75, p. 119–129, https://doi.org/10.1016/j.rama.2020.12.006.
Dataset citation.—Zimmer, S., Grosklos, G., Belmont, P., and Adler, P., 2020 Agreement and uncertainty among climate change impact models—A synthesis of sagebrush steppe vegetation predictions: HydroShare data release, https://doi.org/10.4211/hs.e6b15828d20843eab4e2babd89787f41.
Format.—Rasters (GeoTIFF).
Resolution.—About 15 km2.
Extent.—Their study area included BLM land in the Intermountain West, specifically focusing on the Northern Basin and Range, Central Basin and Range, Wyoming Basin, and Colorado Plateaus U.S. Environmental Protection Agency Level III Ecoregions (https://gaftp.epa.gov/EPADataCommons/ORD/Ecoregions).
Climate Scenarios.—When available, Zimmer and others (2021) separately compiled results for low (for example, RCP4.5) and high (for example, RCP8.5) emissions scenarios, and they focused on the latest periods available (generally the end of 21st century).
Variables.—Count rasters are provided for cheatgrass, forage production, pinyon-juniper, and sagebrush and provide the number of studies with positive or negative projected change at each grid-cell.
3.4.8. Comer and others (2019)—Habitat Climate Change Vulnerability Index
Overview.—Comer and others (2019) created the Habitat Climate Change Vulnerability Index (HCCVI) for different vegetation types, including sagebrush dominated vegetation types. The HCCVI estimates how vulnerable the vegetation in a given location is to climate change. It applies to a vegetation type instead of a specific species or plant functional group. The HCCVI is a mean of multiple scores or subindices including climate exposure, sensitivity, and adaptive capacity. Climate exposure incorporates how projected future climate in a location will compare to the historical spatial variability in climate for the vegetation type and how different future climate will be from historical climate for that location. The sensitivity score attempts to quantify ecosystem stressors that are likely to affect ecological responses to climate change by including estimates of factors such as insect disease risk, invasive plants, and the amount of human modification of the landscape. The ecosystem adaptive capacity score was calculated as a fixed metric (in other words, does not vary spatially) for the given vegetation type, and incorporates factors such as diversity within functional species groups, and the amount of topographic and climate variability across the area occupied by the vegetation type.
Pros.—Spatial layers of the HCCVI were created for multiple sagebrush dominated vegetation types such as “inter-mountain basin big sagebrush steppe,” which makes it relevant for the sagebrush region. The HCCVI provides a different independent perspective on climate vulnerability relative to the other datasets listed in this report.
Cons.—The HCCVI applies to a given vegetation type, and therefore it cannot directly account for potentially different responses by different plant functional groups (for example, sagebrush may respond differently to changes in precipitation seasonality than C4 grasses will). Sometimes, it may be challenging to interpret HCCVI because it is a mean of many different scores; all scores are measures of different factors and likely have varying degrees of uncertainty and error.
Article citation.—Comer P.J., Hak, J.C., Reid, M.S., Auer, S.L., Schulz, K.A., Hamilton, H.H., Smyth, R.L., Kling, M.M., 2019, Habitat Climate Change Vulnerability Index applied to major vegetation types of the Western Interior United States: Land, v. 8, no. 7, art. 108, 27 p., https://doi.org/10.3390/land8070108.
Dataset citation.—Not applicable.
Where to Download.—The main landing page for the data and supplementary material is available at https://databasin.org/galleries/6704179ca499490bafd2e9080df1908a. The geodatabase (folder ending in “.gdb”) that has the spatially explicit HCCVI data for each vegetation type is available at https://tranxfer.natureserve.org/download/Longterm/Ecology/BLM/.
Format.—Geodatabase.
Resolution.—Data are provided for 100 km2 hexagon shaped polygons.
Extent.—Spatial datasets are available for 52 different vegetation types. Each vegetation type has a different extent, but collectively they cover the interior Western United States.
Climate Scenarios.—RCP4.5 (from 2040 to 2069).
Variables.—The HCCVI (a unitless index that ranges from 0 to 1) and scores or indices used for calculating HCCVI including adaptive capacity, resilience, and exposure.
3.4.9. Renwick and others (2018)—Multi-Model Comparison of Sagebrush Responses to Climate Change
Overview.—Renwick and others (2018) used four different models to estimate sagebrush responses to climate change. Although they did not create a raster layer of projected responses, they estimated projected sagebrush responses at 714 sites across the sagebrush region. For each location, they projected responses using 1 time-series correlation-based model, 1 spatial correlation-based model, and 2 process based models. This study is one of only two multimodel comparisons that we are aware of in the sagebrush region (the other is Zimmer and others [2021]). This study helps assess the confidence in the direction of the responses of big sagebrush to climate change across its range.
Pros.—Because this study used multiple independently developed correlation and process-based models, greater confidence is possible in projections in areas where there is agreement among models.
Cons.—Because model projections were made for point locations this dataset will not be usable as is for individuals who want a raster layer of projected sagebrush responses.
Article citation.—Renwick, K.M., Curtis, C., Kleinhesselink, A.R., Schlaepfer, D., Bradley, B.A., Aldridge, C.L., Poulter, B., and Adler, P.B., 2018, Multi-model comparison highlights consistency in predicted effect of warming on a semi-arid shrub: Global Change Biology, v. 24, no. 1, p. 424–438, https://doi.org/10.1111/gcb.13900.
Dataset citation.—Same as the article citation.
Where to Download.—The file ”merged_data_GCM.csv” includes the projections for all climate scenarios and models and is available at https://github.com/krenwick/sageseer/tree/master/Products.
Format.—Comma-separated values (.CSV) file
Resolution.—Not a raster dataset (data for point locations).
Extent.—Projections were made for 714 sites with documented sagebrush presence scattered evenly across the sagebrush region.
Climate Scenarios.—RCP4.5 and RCP8.5 from 2070 to 2099.
Variables.—Because the models did not all have the same response variables, the following data for multiple response variables are provided: maximum potential percentage of sagebrush cover (spatial correlation model), annual change in percentage of sagebrush cover (temporal correlation model), percentage of sagebrush cover (dynamic global vegetation model), and percentage of years with sagebrush regeneration (seedling survival model).
Methods For Developing This Science Synthesis
A search of peer-reviewed journal articles and government reports relevant to the response of vegetation to climate change was performed. The initial search was primarily a narrative literature review, which relied on author expertise and knowledge of the field to gather sources and synthesize information. From this set of articles, the citations for each article were reviewed to identify other relevant climate change articles focused on sagebrush ecosystems. This set of citations formed the basis of the synthesis. The literature was supplemented with a structured search using terms relevant to each section of the report.
The literature review was developed with the goal to inform Federal land management decisions, actions, and analyses related to native plant communities in upland environments within the big sagebrush region. While creating this report, regular input from staff at the BLM and other Federal land management agencies and the U.S. Geological Survey scientists who work closely with land managers in the sagebrush region was received. This input was used to inform the structure and content of the synthesis to maximize information needs relevant to land managers that want to consider climate change effects on big sagebrush plant communities in their management actions, decisions, and associated NEPA analyses.
References Cited
Abatzoglou, J.T., 2013, Development of gridded surface meteorological data for ecological applications and modelling: International Journal of Climatology, v. 33, no. 1, p. 121–131, accessed January 2024 at https://doi.org/10.1002/joc.3413.
Abatzoglou, J.T., and Brown, T.J., 2012, A comparison of statistical downscaling methods suited for wildfire applications: International Journal of Climatology, v. 32, no. 5, p. 772–780, accessed January 2024 at https://doi.org/10.1002/joc.2312.
Abatzoglou, J.T., and Kolden, C.A., 2011, Climate change in Western US deserts—Potential for increased wildfire and invasive annual grasses: Rangeland Ecology & Management, v. 64, no. 5, p. 471–478, accessed January 2024 at https://doi.org/10.2111/REM-D-09-00151.1.
Adler, P.B., HilleRisLambers, J., and Levine, J.M., 2009, Weak effect of climate variability on coexistence in a sagebrush steppe community: Ecology, v. 90, no. 12, p. 3303–3312, accessed January 2024 at https://doi.org/10.1890/08-2241.1.
Adler, P., Renwick, K., Kachergis, E., Manning, M., Remington, T., Thacker, E., Aldridge, C., Bradley, B., Kleinhesselink, A., Curtis, C., Schlaepfer, D., and Poulter, B., 2018, Managing big sagebrush in a changing climate: Utah State University Extension Paper 1882, 5 p., accessed January 2024 at https://digitalcommons.usu.edu/extension_curall/1882/.
Aikens, E.O., Kauffman, M.J., Merkle, J.A., Dwinnell, S.P.H., Fralick, G.L., and Monteith, K.L., 2017, The greenscape shapes surfing of resource waves in a large migratory herbivore: Ecology Letters, v. 20, no. 6, p. 741–750, accessed January 2024 at https://doi.org/10.1111/ele.12772.
Ainsworth, E.A., and Long, S.P., 2005, What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2: New Phytologist, v. 165, no. 2, p. 351–372, accessed January 2024 at https://doi.org/10.1111/j.1469-8137.2004.01224.x.
Andrén, H., 1994, Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat—A review: Oikos, v. 71, no. 3, p. 355–366, accessed January 2024 at https://doi.org/10.2307/3545823.
Applestein, C., Caughlin, T.T., and Germino, M.J., 2021, Weather affects post-fire recovery of sagebrush-steppe communities and model transferability among sites: Ecosphere, v. 12, no. 4, art. e03446, 21 p., accessed January 2024 at https://doi.org/10.1002/ecs2.3446.
Armal, S., Devineni, N., and Khanbilvardi, R., 2018, Trends in extreme rainfall frequency in the contiguous United States—Attribution to climate change and climate variability modes: Journal of Climate, v. 31, no. 1, p. 369–385, accessed January 2024 at https://doi.org/10.1175/JCLI-D-17-0106.1.
Baker, W.L., 2013, Is wildland fire increasing in sagebrush landscapes of the Western United States?: Annals of the Association of American Geographers, v. 103, no. 1, p. 5–19, accessed January 2024 at https://doi.org/10.1080/00045608.2012.732483.
Balch, J.K., Bradley, B.A., D’Antonio, C.M., and Gómez-Dans, J., 2013, Introduced annual grass increases regional fire activity across the arid Western USA (1980–2009): Global Change Biology, v. 19, no. 1, p. 173–183, accessed January 2024 at https://doi.org/10.1111/gcb.12046.
Barga, SC., Dilts, T.E., and Leger, E.A., 2018, Contrasting climate niches among co-occurring subdominant forbs of the sagebrush steppe: Diversity and Distributions, v. 24, no. 9, p. 1291–1307, accessed January 2024 at https://doi.org/10.1111/ddi.12764.
Barker, B.S., Pilliod, D.S., Rigge, M., and Homer, C.G., 2019, Pre‐fire vegetation drives post‐fire outcomes in sagebrush ecosystems—Evidence from field and remote sensing data: Ecosphere, v. 10, no. 11, art. e02929, 29 p., accessed January 2024 at https://doi.org/10.1002/ecs2.2929.
Bates, J.D., Svejcar, T., Miller, R.F., and Angell, R.A., 2006, The effects of precipitation timing on sagebrush steppe vegetation: Journal of Arid Environments, v. 64, no. 4, p. 670–697, accessed January 2024 at https://doi.org/10.1016/j.jaridenv.2005.06.026.
Beever, E.A., Tausch, R.J., and Thogmartin, W.E., 2008, Multi-scale responses of vegetation to removal of horse grazing from Great Basin (USA) mountain ranges: Plant Ecology, v. 196, p. 163–184, accessed January 2024 at https://doi.org/10.1007/s11258-007-9342-5.
Bishop, T.B.B., Nusink, B.C., Lee Molinari, R., Taylor, J.B., and St. Clair, S.B., 2020, Earlier fall precipitation and low severity fire impacts on cheatgrass and sagebrush establishment: Ecosphere, v. 11, no. 1, art. e03019, 13 p., accessed January 2024 at https://doi.org/10.1002/ecs2.3019.
Bloom, T.D.S., O’Leary, D.S., and Riginos, C., 2022, Flowering time advances since the 1970s in a sagebrush steppe community—Implications for management and restoration: Ecological Applications, v. 32, no. 6, art. e2583, 17 p., accessed January 2024 at https://doi.org/10.1002/eap.2583.
Blumenthal, D.M., Kray, J.A., Ortmans, W., Ziska, L.H., and Pendall, E., 2016, Cheatgrass is favored by warming but not CO2 enrichment in a semi-arid grassland: Global Change Biology, v. 22, no. 9, p. 3026–3038, accessed January 2024 at https://doi.org/10.1111/gcb.13278.
Booth, D., and Bai, Y., 2000, Seeds and seedling establishment of Wyoming big sagebrush, in Schuman, G.E., Richmond, T.C., and Neuman, D.R., eds., Sagebrush establishment on mined lands—Ecology and research—Billings Land Reclamation Symposium, Billings, Mont., March 20–24, 2000, [Proceedings]: Billings, Mont., [U.S. Department of Agriculture Forest Service], p. 24–31. [Also available at https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=b4db7061b630956c335b9445d634daba99ec5e95#page=28.]
Bowman, D.M.J.S., Kolden, C.A., Abatzoglou, J.T., Johnston, F.H., van der Werf, G.R., and Flannigan, M., 2020, Vegetation fires in the Anthropocene: Nature Reviews Earth & Environment, v. 1, p. 500–515, accessed January 2024 at https://doi.org/10.1038/s43017-020-0085-3.
Boyte, S.P., Wylie, B.K., and Major, D.J., 2016, Cheatgrass percent cover change—Comparing recent estimates to climate change−driven [sic] predictions in the Northern Great Basin: Rangeland Ecology & Management, v. 69, no. 4, p. 265–279, accessed January 2024 at https://doi.org/10.1016/j.rama.2016.03.002.
Brabec, M.M., Germino, M.J., and Richardson, B.A., 2017, Climate adaption and post-fire restoration of a foundational perennial in cold desert—Insights from intraspecific variation in response to weather: Journal of Applied Ecology, v. 54, no. 1, p. 293–302, accessed January 2024 at https://doi.org/10.1111/1365-2664.12679.
Bradford, J.B., Betancourt, J.L., Butterfield, B.J., Munson, S.M., and Wood, T.E., 2018, Anticipatory natural resource science and management for a changing future: Frontiers in Ecology and the Environment, v. 16, no. 6, p. 295–303, accessed January 2024 at https://doi.org/10.1002/fee.1806.
Bradford, J.B., Schlaepfer, D.R., and Lauenroth, W.K., 2014, Ecohydrology of adjacent sagebrush and lodgepole pine ecosystems—The consequences of climate change and disturbance: Ecosystems, v. 17, p. 590–605, accessed January 2024 at https://doi.org/10.1007/s10021-013-9745-1.
Bradford, J.B., Schlaepfer, D.R., Lauenroth, W.K., and Palmquist, K.A., 2020, Robust ecological drought projections for drylands in the 21st century: Global Change Biology, v. 26, no. 7, p. 3906–3919, accessed January 2024 at https://doi.org/10.1111/gcb.15075.
Bradford, J.B., Schlaepfer, D.R., Lauenroth, W.K., Palmquist, K.A., Chambers, J.C., Maestas, J.D., and Campbell, S.B., 2019, Climate-driven shifts in soil temperature and moisture regimes suggest opportunities to enhance assessments of dryland resilience and resistance: Frontiers in Ecology and Evolution, v. 7, art. 358, 16 p., accessed January 2024 at https://doi.org/10.3389/fevo.2019.00358.
Bradley, B.A., 2009, Regional analysis of the impacts of climate change on cheatgrass invasion shows potential risk and opportunity: Global Change Biology, v. 15, no. 1, p. 196–208, accessed January 2024 at https://doi.org/10.1111/j.1365-2486.2008.01709.x.
Bradley, B.A., 2010, Assessing ecosystem threats from global and regional change—Hierarchical modeling of risk to sagebrush ecosystems from climate change, land use and invasive species in Nevada, USA: Ecography, v. 33, no. 1, p. 198–208, accessed January 2024 at https://doi.org/10.1111/j.1600-0587.2009.05684.x.
Bradley, B.A., Curtis, C.A., Fusco, E.J., Abatzoglou, J.T., Balch, J.K., Dadashi, S., and Tuanmu, M.-N., 2018, Cheatgrass (Bromus tectorum) distribution in the intermountain Western United States and its relationship to fire frequency, seasonality, and ignitions: Biological Invasions, v. 20, p. 1493–1506, accessed January 2024 at https://doi.org/10.1007/s10530-017-1641-8.
Bradley, B.A., Curtis, C.A., and Chambers, J.C., 2016, Bromus response to climate and projected changes with climate change, chap. 9 of Germino, M.J., Chambers, J.C., and Brown, C.S., eds., Exotic brome-grasses in arid and semiarid ecosystems of the Western US—Causes, consequences, and management implications: Cham, Switzerland, Springer International Publishing, p. 257–274.
Breshears, D.D., Knapp, A.K., Law, D.J., Smith, M.D., Twidwell, D., and Wonkka, C.L., 2016, Rangeland responses to predicted increases in drought extremity: Rangelands, v. 38, no. 4, p. 191–196, accessed January 2024 at https://doi.org/10.1016/j.rala.2016.06.009.
Brice, E.M., Miller, B.A., Zhang, H., Goldstein, K., Zimmer, S.N., Grosklos, G.J., Belmont, P., Flint, C.G., Givens, J.E., Adler, P.B., Brunson, M.W., and Smith, J.W., 2020, Impacts of climate change on multiple use management of Bureau of Land Management land in the Intermountain West, USA: Ecosphere, v. 11, no. 11, art. e03286, 29 p., accessed January 2024 at https://doi.org/10.1002/ecs2.3286.
Brunson, M.W., and Shindler, B.A., 2004, Geographic variation in social acceptability of wildland fuels management in the Western United States: Society and Natural Resources, v. 17, no. 8, p. 661–678, accessed January 2024 at https://doi.org/10.1080/08941920490480688.
Bushey, J.A., Hoffman, A.M., Gleason, S.M., Smith, M.D., and Ocheltree, T.W., 2023, Water limitation reveals local adaptation and plasticity in the drought tolerance strategies of Bouteloua gracilis: Ecosphere, v. 14, no. 1, art. e4335, 12 p., accessed January 2024 at https://doi.org/10.1002/ecs2.4335.
Cabral, A.C., De Miguel, J.M., Rescia, A.J., Schmitz, M.F., and Pineda, F.D., 2003, Shrub encroachment in Argentinean savannas: Journal of Vegetation Science, v. 14, no. 2, p. 145–152, accessed January 2024 at https://doi.org/10.1111/j.1654-1103.2003.tb02139.x.
Chambers, J.C., Beck, J.L., Bradford, J.B., Bybee, J., Campbell, S., Carlson, J., Christiansen, T.J., Clause, K.J., Collins, G., Crist, M.R., Dinkins, J.B., Doherty, K.E., Edwards, F., Espinosa, S., Griffin, K.A., Griffin, P., Haas, J.R., Hanser, S.E., Havlina, D.W., Henke, K.F., Hennig, J.D., Joyce, L.A., Kilkenny, F.M., Kulpa, S.M., Kurth, L.L., Maestas, J.D., Manning, M., Mayer, K.E., Mealor, B.A., McCarthy, C., Pellant, M., Perea, M.A., Prentice, K.L., Pyke, D.A., Wiechman, L.A., and Wuenschel, A., 2017, Science framework for conservation and restoration of the sagebrush biome—Linking the Department of the Interior’s Integrated Rangeland Fire Management Strategy to long-term strategic conservation actions: Fort Collins, Colo., U.S. Department of Agriculture Forest Service Rocky Mountain Research Station General Technical Report RMRS-GTR-360, 213 p., accessed October 29, 2024, at https://research.fs.usda.gov/treesearch/53983.
Chambers, J.C., Brooks, M.L., Germino, M.J., Maestas, J.D., Board, D.I., Jones, M.O., and Allred, B.W., 2019, Operationalizing resilience and resistance concepts to address invasive grass–fire cycles: Frontiers in Ecology and Evolution, v. 7, art. 185, 25 p., accessed January 2024 at https://doi.org/10.3389/fevo.2019.00185.
Chambers, J.C., Brown, J.L., Bradford, J.B., Board, D.I., Campbell, S.B., Clause, K.J., Hanberry, B., Schlaepfer, D.R., and Urza, A.K., 2023, New indicators of ecological resilience and invasion resistance to support prioritization and management in the sagebrush biome, United States: Frontiers in Ecology and Evolution, v. 10, 17 p., accessed January 2024 at https://doi.org/10.3389/fevo.2022.1009268.
Chambers, J.C., Maestas, J.D., Pyke, D.A., Boyd, C.S., Pellant, M., and Wuenschel, A., 2017, Using resilience and resistance concepts to manage persistent threats to sagebrush ecosystems and greater sage-grouse: Rangeland Ecology & Management, v. 70, no. 2, p. 149–164, accessed January 2024 at https://doi.org/10.1016/j.rama.2016.08.005.
Chambers, J.C., Miller, R.F., Board, D.I., Pyke, D.A., Roundy, B.A., Grace, J.B., Schupp, E.W., and Tausch, R.J., 2014, Resilience and resistance of sagebrush ecosystems—Implications for state and transition models and management treatments: Rangeland Ecology & Management, v. 67, no. 5, p. 440–454, accessed January 2024 at https://doi.org/10.2111/REM-D-13-00074.1.
Coates, P.S., Ricca, M.A., Prochazka, B.G., Brooks, M.L., Doherty, K.E., Kroger, T., Blomberg, E.J., Hagen, C.A., and Casazza, M.L., 2016, Wildfire, climate, and invasive grass interactions negatively impact an indicator species by reshaping sagebrush ecosystems: Proceedings of the National Academy of Sciences, v. 113, no. 45, p. 12745–12750, accessed January 2024 at https://doi.org/10.1073/pnas.1606898113.
Compagnoni, A., and Adler, P.B. 2014a, Warming, competition, and Bromus tectorum population growth across an elevation gradient: Ecosphere, v. 5, no. 9, art. 121, p. 1–34, accessed January 2024 at https://doi.org/10.1890/ES14-00047.1.
Compagnoni, A., and Adler, P.B., 2014b, Warming, soil moisture, and loss of snow increase Bromus tectorum’s population growth rate: Elementa—Science of the Anthropocene, v. 2, 10 p., accessed January 2024 at https://doi.org/10.12952/journal.elementa.000020.
Comer P.J., Hak, J.C., Reid, M.S., Auer, S.L., Schulz, K.A., Hamilton, H.H., Smyth, R.L., Kling, M.M., 2019, Habitat Climate Change Vulnerability Index applied to major vegetation types of the Western Interior United States: Land, v. 8, no. 7, art. 108, 27 p., accessed January 2024 https://doi.org/10.3390/land8070108.
Coop, J.D., Parks, S.A., Stevens-Rumann, C.S., Ritter, S.M., and Hoffman, C.M., 2022, Extreme fire spread events and area burned under recent and future climate in the Western USA: Global Ecology and Biogeography, v. 31, no. 10, p. 1949–1959, accessed January 2024 at https://doi.org/10.1111/geb.13496.
Copeland, S.M., Munson, S.M., Bradford, J.B., Butterfield, B.J., and Gunnell, K.L., 2019, Long-term plant community trajectories suggest divergent responses of native and non-native perennials and annuals to vegetation removal and seeding treatments: Restoration Ecology, v. 27, no. 4, p. 821–831, accessed January 2024 at https://doi.org/10.1111/rec.12928.
Cross, M.S., and Harte, J., 2007, Compensatory responses to loss of warming-sensitive plant species: Ecology, v. 88, no. 3, p. 740–748, accessed January 2024 at https://doi.org/10.1890/06-1029.
Crous, K.Y., Reich, P.B., Hunter, M.D., and Ellsworth, D.S., 2010, Maintenance of leaf N controls the photosynthetic CO2 response of grassland species exposed to 9 years of free-air CO2 enrichment: Global Change Biology, v. 16, no. 7, p. 2076–2088, accessed January 2024 at https://doi.org/10.1111/j.1365-2486.2009.02058.x.
Curtis, P.S., and Wang, X., 1998, A meta-analysis of elevated CO2 effects on woody plant mass, form, and physiology: Oecologia, v. 113, p. 299–313, accessed January 2024 at https://doi.org/10.1007/s004420050381.
D’Antonio, C.M., and Vitousek, P.M. 1992, Biological invasions by exotic grasses, the grass/fire cycle, and global change: Annual Review of Ecology, Evolution, and Systematics, v. 23, p. 63–87, accessed January 2024 at https://www.jstor.org/stable/2097282. https://doi.org/10.1146/annurev.es.23.110192.000431.
Davies, K.W., and Bates, J.D., 2014, Attempting to restore herbaceous understories in Wyoming big sagebrush communities with mowing and seeding: Restoration Ecology, v. 22, no. 5, p. 608–615, accessed January 2024 at https://doi.org/10.1111/rec.12110.
Davies, K.W., and Bates, J.D., 2019, Longer-term evaluation of sagebrush restoration after juniper control and herbaceous vegetation trade-offs: Rangeland Ecology & Management, v. 72, no. 2, p. 260–265, accessed January 2024 at https://doi.org/10.1016/j.rama.2018.10.006.
Davies, K.W., Boyd, C.S., Beck, J.L., Bates, J.D., Svejcar, T.J., and Gregg, M.A., 2011, Saving the sagebrush sea—An ecosystem conservation plan for big sagebrush plant communities: Biological Conservation, v. 144, no. 11, p. 2573–2584, accessed January 2024 at https://doi.org/10.1016/j.biocon.2011.07.016.
Davies, K.W., Leger, E.A., Boyd, C.S., and Hallett, L.M., 2021, Living with exotic annual grasses in the sagebrush ecosystem: Journal of Environmental Management, v. 288, art. 112417, 11 p., accessed January 2024 at https://doi.org/10.1016/j.jenvman.2021.112417.
Davies, K.W., and Svejcar, T.J., 2008, Comparison of medusahead-invaded and noninvaded Wyoming big sagebrush steppe in Southeastern Oregon: Rangeland Ecology & Management, v. 61, no. 6, p. 623–629, accessed January 2024 at https://doi.org/10.2111/08-005.1.
Davies, K.W., Wollstein, K., Dragt, B., and O’Connor, C., 2022, Grazing management to reduce wildfire risk in invasive annual grass prone sagebrush communities: Rangelands, v. 44, no. 3, p. 194–199, accessed January 2024 at https://doi.org/10.1016/j.rala.2022.02.001.
Díaz, S., and Cabido, M., 1997, Plant functional types and ecosystem function in relation to global change: Journal of Vegetation Science, v. 8, no. 4, p. 463–474, accessed January 2024 at https://doi.org/10.2307/3237198.
Doherty, K., Theobald, D.M., Bradford, J.B., Wiechman, L.A., Bedrosian, G., Boyd, C.S., Cahill, M., Coates, P.S., Creutzburg, M.K., Crist, M.R., Finn, S.P., Kumar, A.V., Littlefield, C.E., Maestas, J.D., Prentice, K.L., Prochazka, B.G., Remington, T.E., Sparklin, W.D., Tull, J.C., Wurtzebach, Z., and Zeller, K.A., 2022, A sagebrush conservation design to proactively restore America’s sagebrush biome: U.S. Geological Survey Open-File Report 2022–1081, 38 p., accessed January 2024 at https://doi.org/10.3133/ofr20221081.
Doherty, K., Theobald, D.M., Holdrege, M.C., Wiechman, L.A., and Bradford, J.B., 2022, Biome-wide sagebrush core habitat and growth areas estimated from a threat-based conservation design: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P94Y5CDV.
Dolezal, J., Jandova, V., Macek, M., Mudrak, O., Altman, J., Schweingruber, F.H., and Liancourt, P., 2021, Climate warming drives Himalayan alpine plant growth and recruitment dynamics: Journal of Ecology, v. 109, no. 1, p. 179–190, accessed January 2024 at https://doi.org/10.1111/1365-2745.13459.
Du, H., Wang, Z., Yu, W., Liu, Y., and Huang, B., 2011, Differential metabolic responses of perennial grass Cynodon transvaalensis×Cynodon dactylon (C4) and Poa Pratensis (C3) to heat stress: Physiologia Plantarum, v. 141, no. 3, p. 251–264, accessed January 2024 at https://doi.org/10.1111/j.1399-3054.2010.01432.x.
Edwards, B.L., Webb, N.P., Brown, D.P., Elias, E., Peck, D.E., Pierson, F.B., Williams, C.J., and Herrick, J.E., 2019, Climate change impacts on wind and water erosion on US rangelands: Journal of Soil and Water Conservation, v. 74, no. 4, p. 405–418, accessed January 2024 at https://doi.org/10.2489/jswc.74.4.405.
Ellsworth, L.M., Newingham, B.A., Shaff, S.E., Williams, C.L., Strand, E.K., Reeves, M., Pyke, D.A., Schupp, E.W., and Chambers, J.C., 2022, Fuel reduction treatments reduce modeled fire intensity in the sagebrush steppe: Ecosphere, v. 13, no. 3, art. e4064, 20 p., accessed January 2024 at https://doi.org/10.1002/ecs2.4064.
Fahrig, L., 2003, Effects of habitat fragmentation on biodiversity: Annual Review of Ecology, Evolution, and Systematics, v. 34, p. 487–515, accessed January 2024 at https://doi.org/10.1146/annurev.ecolsys.34.011802.132419.
Felton, A.J., Knapp, A.K., and Smith, M.D., 2021, Precipitation–productivity relationships and the duration of precipitation anomalies—An underappreciated dimension of climate change: Global Change Biology, v. 27, no. 6, p. 1127–1140, accessed January 2024 at https://doi.org/10.1111/gcb.15480.
Felton, A.J., Shriver, R.K., Stemkovski, M., Bradford, J.B., Suding, K.N., and Adler, P.B., 2022, Climate disequilibrium dominates uncertainty in long-term projections of primary productivity: Ecology Letters, v. 25, no. 12, p. 2688–2698, accessed January 2024 at https://doi.org/10.1111/ele.14132.
Flannigan, M.D., Wotton, B. M., Marshall, G.A., de Groot, W.J., Johnston, J., Jurko, N., and Cantin, A.S., 2016, Fuel moisture sensitivity to temperature and precipitation—Climate change implications: Climatic Change, v. 134, p. 59–71, accessed January 2024 at https://doi.org/10.1007/s10584-015-1521-0.
Germaine, S.S., Assal, T., Freeman, A., and Carter, S.K., 2020, Distance effects of gas field infrastructure on pygmy rabbits in southwestern Wyoming: Ecosphere, v. 11, no. 8, art. e03230, 16 p., accessed January 2024 at https://doi.org/10.1002/ecs2.3230.
Germino, M.J., Barnard, D.M., Davidson, B.E., Arkle, R.S., Pilliod, D.S., Fisk, M.R., and Applestein, C., 2018, Thresholds and hotspots for shrub restoration following a heterogeneous megafire: Landscape Ecology, v. 33, p. 1177–1194, accessed January 2024 at https://doi.org/10.1007/s10980-018-0662-8.
Germino, M.J., Belnap, J., Stark, J.M., Allen, E.B., and Rau, B.M., 2016, Ecosystem impacts of exotic annual invaders in the Genus Bromus, in Germino, M.J., Chambers, J.C., and Brown, C.S., eds., Exotic brome-grasses in arid and semiarid ecosystems of the Western US—Causes, consequences, and management implications: Cham, Switzerland, Springer International Publishing, p. 61–95, accessed January 2024 at https://doi.org/10.1007/978-3-319-24930-8_3.
Gherardi, L.A., and Sala, O.E., 2015, Enhanced precipitation variability decreases grass- and increases shrub-productivity: Proceedings of the National Academy of Sciences, v. 112, no. 41, p. 12735–12740, accessed January 2024 at https://doi.org/10.1073/pnas.1506433112.
Gherardi, L.A., and Sala, O.E., 2019, Effect of interannual precipitation variability on dryland productivity—A global synthesis: Global Change Biology, v. 25, no. 1, p. 269–276, accessed January 2024 at https://doi.org/10.1111/gcb.14480.
Gordon, R., Brunson, M.W., and Shindler, B., 2014, Acceptance, acceptability, and trust for sagebrush restoration options in the Great Basin—A longitudinal perspective: Rangeland Ecology & Management, v. 67, no. 5, p. 573–583, accessed January 2024 at https://doi.org/10.2111/REM-D-13-00016.1.
Gornish, E.S., Guo, J.S., Porensky, L.M., Perryman, B.L., and Leger, E.A., 2023, Pre‐fire grazing and herbicide treatments can affect post‐fire vegetation in a Great Basin rangeland: Ecological Solutions and Evidence, v. 4, no. 1, art. e12215, 12 p., accessed January 2024 at https://doi.org/10.1002/2688-8319.12215.
Hamerlynck, E.P., Huxman, T.E., Loik, M.E., and Smith, S.D., 2000, Effects of extreme high temperature, drought and elevated CO2 on photosynthesis of the Mojave Desert evergreen shrub, Larrea tridentata: Plant Ecology, v. 148, p. 183–193, accessed January 2024 at https://doi.org/10.1023/A:1009896111405.
Harris, G.A., 1967, Some competitive relationships between Agropyron spicatum and Bromus tectorum: Ecological Monographs, v. 37, no. 2, p. 89–111, accessed January 2024 at https://doi.org/10.2307/2937337.
Harte, J., and Shaw, R., 1995, Shifting dominance within a montane vegetation community—Results of a climate-warming experiment: Science, v. 267, no. 5199, p. 876–880, accessed January 2024 at https://doi.org/10.1126/science.267.5199.876.
Hartman, J.C., and Nippert, J.B., 2013, Physiological and growth responses of switchgrass (Panicum virgatum L.) in native stands under passive air temperature manipulation: Global Change Biology Bioenergy, v. 5, no. 6, p. 683–692, accessed January 2024 at https://doi.org/10.1111/j.1757-1707.2012.01204.x.
Havrilla, C.A., Bradford, J.B., Yackulic, C.B., and Munson, S.M., 2023, Divergent climate impacts on C3 versus C4 grasses imply widespread 21st century shifts in grassland functional composition: Diversity and Distributions, v. 29, no. 3, p. 379–394, accessed January 2024 at https://doi.org/10.1111/ddi.13669.
Holdrege, M.C., Beard, K.H., and Kulmatiski, A., 2021, Woody plant growth increases with precipitation intensity in a cold semiarid system: Ecology, v. 102, no. 1, art. e03212, 11 p., accessed January 2024 at https://doi.org/10.1002/ecy.3212.
Holdrege, M.C., Kulmatiski, A., Beard, K.H., and Palmquist, K.A., 2023, Precipitation intensification increases shrub dominance in arid, not mesic, ecosystems: Ecosystems, v. 26, p. 568–584, accessed January 2024 at https://doi.org/10.1007/s10021-022-00778-1.
Holdrege, M.C., Palmquist, K.A., Schlaepfer, D.R., Lauenroth, W.K., Boyd, C.S., Creutzburg, M.K., Crist, M.R., Doherty, K.E., Remington, T.E., Tull, J.C., Wiechman, L.A., and Bradford, J.B., 2024a, Climate change amplifies ongoing declines in sagebrush ecological integrity: Rangeland Ecology & Management, v. 97, p. 25–40, accessed October 18, 2024, at https://doi.org/10.1016/j.rama.2024.08.003.
Holdrege, M.C., Schlaepfer, D.R., Palmquist, K.A., Crist, M., Doherty, K.E., Lauenroth, W.K., Remington, T.E., Riley, K., Short, K.C., Tull, J.C., Wiechman, L.A., and Bradford, J.B., 2024b, Wildfire probability estimated from recent climate and fine fuels across the big sagebrush region: Fire Ecology, v. 20, art. 22, 20 p., accessed January 2024 at https://doi.org/10.1186/s42408-024-00252-4.
Holdrege, M.C., Schlaepfer, D.R., Palmquist, K.A., Theobald, D.M., and Bradford, J.B., 2024, Current and projected sagebrush ecological integrity across the Western U.S., 2017–2100: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P13RXYZJ.
Homer, C.G., Aldridge, C.L., Meyer, D.K., and Schell, S.J. 2012, Multi-scale remote sensing sagebrush characterization with regression trees over Wyoming, USA—Laying a foundation for monitoring: International Journal of Applied Earth Observation and Geoinformation, v. 14, no. 1, p. 233–244, accessed January 2024 at https://doi.org/10.1016/j.jag.2011.09.012.
Homer, C.G., Xian, G., Aldridge, C.L., Meyer, D.K., Loveland, T.R., and O’Donnell, M.S., 2015, Forecasting sagebrush ecosystem components and greater sage-grouse habitat for 2050—Learning from past climate patterns and Landsat imagery to predict the future: Ecological Indicators, v. 55, p. 131–145, accessed January 2024 at https://doi.org/10.1016/j.ecolind.2015.03.002.
Hoover, D.L., Duniway, M.C., and Belnap, J., 2015, Pulse-drought atop press-drought—unexpected plant responses and implications for dryland ecosystems: Oecologia, v. 179, p. 1211–1221, accessed January 2024 at https://doi.org/10.1007/s00442-015-3414-3.
Hoover, D.L., Knapp, A.K., and Smith, M.D., 2014, Resistance and resilience of a grassland ecosystem to climate extremes: Ecology, v. 95, no. 9, p. 2646–2656, accessed January 2024 at https://doi.org/10.1890/13-2186.1.
Hou, E., Litvak, M.E., Rudgers, J.A., Jiang, L., Collins, S.L., Pockman, W.T., Hui, D., Niu, S., and Luo, Y., 2021, Divergent responses of primary production to increasing precipitation variability in global drylands: Global Change Biology, v. 27, no. 20, p. 5225–5237, accessed January 2024 at https://doi.org/10.1111/gcb.15801.
Howell, A., Winkler, D.E., Phillips, M.L., McNellis, B., and Reed, S.C., 2020, Experimental warming changes phenology and shortens growing season of the dominant invasive plant Bromus tectorum (cheatgrass): Frontiers in Plant Science, v. 11, 15 p., accessed January 2024 at https://doi.org/10.3389/fpls.2020.570001.
Hsiung, A.C., Boyle, W.A., Cooper, R.J., and Chandler, R.B., 2018, Altitudinal migration—Ecological drivers, knowledge gaps, and conservation implications: Biological Reviews, v. 93, no. 4, p. 2049–2070, accessed January 2024 at https://doi.org/10.1111/brv.12435.
Hsu, J.S., Powell, J., and Adler, P.B., 2012, Sensitivity of mean annual primary production to precipitation: Global Change Biology, v. 18, no. 7, p. 2246–2255, accessed January 2024 at https://doi.org/10.1111/j.1365-2486.2012.02687.x.
Huang, J., Yu, H., Guan, X., Wang, G., and Guo, R., 2016, Accelerated dryland expansion under climate change: Nature Climate Change, v. 6, p. 166–171, accessed January 2024 at https://doi.org/10.1038/nclimate2837.
Hunt, H.W., Elliott, E.T., Detling, J.K., Morgan, J.A., and Chen, D.-X., 1996, Responses of a C3 and a C4 perennial grass to elevated CO2 and temperature under different water regimes: Global Change Biology, v. 2, no. 1, p. 35–47, accessed January 2024 at https://doi.org/10.1111/j.1365-2486.1996.tb00047.x.
Idso, S.B., and Idso, K.E., 2001, Effects of atmospheric CO2 enrichment on plant constituents related to animal and human health: Environmental and Experimental Botany, v. 45, no. 2, p. 179–199, accessed January 2024 at https://doi.org/10.1016/S0098-8472(00)00091-5.
Intergovernmental Panel on Climate Change [IPCC], 2014, Climate change 2014—Synthesis report, in Core Writing Team, Pachauri, R.K, and Meyer, L., eds., Contribution of working groups I, II, and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change: Geneva, Switzerland, Intergovernmental Panel on Climate Change, 151 p., accessed June 2022 at https://www.ipcc.ch/report/ar5/syr/.
Intergovernmental Panel on Climate Change [IPCC], 2022, Climate change 2022—Impacts, adaptation and vulnerability—Working group II contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change: Cambridge University Press, 3,068 p., accessed January 2024 at https://www.ipcc.ch/report/ar6/wg2/.
Jeffries, M.I., and Finn, S.P., 2019, The sagebrush biome range extent, as derived from classified Landsat imagery: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P950H8HS.
Jones, M.O., Allred, B.W., Naugle, D.E., Maestas, J.D., Donnelly, P., Metz, L.J., Karl, J., Smith, R., Bestelmeyer, B., Boyd, C., Kerby, J.D., and McIver, J.D., 2018, Innovation in rangeland monitoring—Annual, 30 m, plant functional type percent cover maps for U.S. rangelands, 1984–2017: Ecosphere, v. 9, no. 9, art. e02430, 19 p., accessed January 2024 at https://doi.org/10.1002/ecs2.2430.
Jordan, S.E., Palmquist, K.A., Bradford, J.B., and Lauenroth, W.K., 2020, Soil water availability shapes species richness in mid-latitude shrub steppe plant communities: Journal of Vegetation Science, v. 31, no. 4, p. 646–657, accessed January 2024 at https://doi.org/10.1111/jvs.12874.
Karban, R., and Pezzola, E., 2017, Effects of a multi-year drought on a drought-adapted shrub, Artemisia tridentata: Plant Ecology, v. 218, p. 547–554, accessed January 2024 at https://doi.org/10.1007/s11258-017-0710-5.
Kemp, K., Blades, J., Klos, P.Z., Hall, T., Force, J.E., Morgan, P., and Tinkham, W., 2015, Managing for climate change on Federal lands of the Western United States—Perceived usefulness of climate science, effectiveness of adaptation strategies, and barriers to implementation: Ecology & Society, v. 20, no. 2, 14 p., accessed January 2024 at https://doi.org/10.5751/ES-07522-200217.
King, S.R.B., Schoenecker, K.A., and Manier, D J., 2019, Potential spread of cheatgrass and other invasive species by feral horses in Western Colorado: Rangeland Ecology & Management, v. 72, no. 4, p. 706–710, accessed January 2024 at https://doi.org/10.1016/j.rama.2019.02.006.
Kleinhesselink, A.R., and Adler, P.B., 2018, The response of big sagebrush (Artemisia tridentata) to interannual climate variation changes across its range: Ecology, v. 99, no. 5, p. 1139–1149, accessed January 2024 at https://doi.org/10.1002/ecy.2191.
Klemm, T., Briske, D.D., and Reeves, M.C., 2020, Vulnerability of rangeland beef cattle production to climate-induced NPP fluctuations in the US Great Plains: Global Change Biology, v. 26, no. 9, p. 4841–4853, accessed January 2024 at https://doi.org/10.1111/gcb.15202.
Kopp, C.W., and Cleland, E.E., 2015, A range-expanding shrub species alters plant phenological response to experimental warming: PLoS ONE, v. 10, art. e0139029, 9 p., accessed January 2024 at https://doi.org/10.1371/journal.pone.0139029.
Larson, C.D., Lehnhoff, E.A., and Rew, L.J., 2017, A warmer and drier climate in the northern sagebrush biome does not promote cheatgrass invasion or change its response to fire: Oecologia, v. 185, p. 763–774, accessed January 2024 at https://doi.org/10.1007/s00442-017-3976-3.
Lauenroth, W.K., Schlaepfer, D.R., and Bradford, J.B., 2014, Ecohydrology of dry regions—Storage versus pulse soil water dynamics: Ecosystems, v. 17, p. 1469–1479, accessed January 2024 at https://doi.org/10.1007/s10021-014-9808-y.
Lee, E., Kumar, P., Barron-Gafford, G.A., Hendryx, S.M., Sanchez-Cañete, E.P., Minor, R.L., Colella, T., and Scott, R.L., 2018, Impact of hydraulic redistribution on multispecies vegetation water use in a semiarid savanna ecosystem—An experimental and modeling synthesis: Water Resources Research, v. 54, no. 6, p. 4009–4027, accessed January 2024 at https://doi.org/10.1029/2017WR021006.
Lee, T.D., Barrott, S.H., and Reich, P.B., 2011, Photosynthetic responses of 13 grassland species across 11 years of free-air CO2 enrichment is modest, consistent and independent of N supply: Global Change Biology, v. 17, no. 9, p. 2893–2904, accessed January 2024 at https://doi.org/10.1111/j.1365-2486.2011.02435.x.
Levin, S.A., 1992, The problem of pattern and scale in ecology—The Robert H. MacArthur award lecture: Ecology, v. 73, no. 6, p. 1943–1967, accessed January 2024 at https://doi.org/10.2307/1941447.
Liang, M., Feng, X., and Gornish, E.S., 2021, Rainfall pulses mediate long-term plant community compositional dynamics in a semi-arid rangeland: Journal of Applied Ecology, v. 58, no. 4, p. 708–717, accessed January 2024 at https://doi.org/10.1111/1365-2664.13780.
Loik, M.E., and Harte, J., 1996, High-temperature tolerance of Artemisia tridentata and Potentilla gracilis under a climate change manipulation: Oecologia, v. 108, p. 224–231, accessed January 2024 at https://doi.org/10.1007/BF00334645.
Love, S.J., Schweitzer, J.A., Woolbright, S.A., and Bailey, J.K., 2023, Sky islands are a global tool for predicting the ecological and evolutionary consequences of climate change: Annual Review of Ecology, Evolution, and Systematics, v. 54, p. 219–236, accessed January 2024 at https://doi.org/10.1146/annurev-ecolsys-102221-050029.
Lozano-Parra, J., Schnabel, S., Pulido, M., Gómez-Gutiérrez, Á., and Lavado-Contador, F., 2018, Effects of soil moisture and vegetation cover on biomass growth in water-limited environments: Land Degradation & Development, v. 29, no. 12, p. 4405–4414, accessed January 2024 at https://doi.org/10.1002/ldr.3193.
Lucash, M.S., Farnsworth, B., and Winner, W.E., 2005, Response of sagebrush steppe species to elevated CO2 and soil temperature: Western North American Naturalist, v. 65, no. 1, p. 80–86, accessed January 2024 at https://www.jstor.org/stable/pdf/41717425.pdf.
Lynch, A.J., Thompson, L.M., Morton, J.M., Beever, E.A., Clifford, M., Limpinsel, D., Magill, R.T., Magness, D.R., Melvin, T.A., Newman, R.A., Porath, M.T., Rahel, F.J., Reynolds, J.H., Schuurman, G.W., Sethi, S.A., and Wilkening, J.L., 2022, RAD adaptive management for transforming ecosystems: BioScience, v. 72, no. 1, p. 45–56, accessed January 2024 at https://doi.org/10.1093/biosci/biab091.
Maestas, J.D., Naugle, D.E., Chambers, J.C., Tack, J.D., Boyd, C.S., and Tague, J.M., 2021, Conifer expansion, chap. M of Sagebrush conservation strategy—Challenges to sagebrush conservation: U.S. Geological Survey Open-File Report 2020–1125, p. 139–152, accessed January 2024 at https://doi.org/10.3133/ofr20201125.
Maestre, F.T., Quero, J.L., Gotelli, N.J., Escudero, A., Ochoa, V., Delgado-Baquerizo, M., García-Gómez, M., Bowker, M.A., Soliveres, S., Escolar, C., García-Palacios, P., Berdugo, M., Valencia, E., Gozalo, B., Gallardo, A., Aguilera, L., Arredondo, T., Blones, J., Boeken, B., Bran, D., Conceição, A.A., Cabrera, O., Chaieb, M., Derak, M., Eldridge, D.J., Espinosa, C.I., Florentino, A., Gaitán, J., Gatica, M.G., Ghiloufi, W., Gómez-González, S., Gutiérrez, J.R., Hernández, R.M., Huang, X., Huber-Sannwald, E., Jankju, M., Miriti, M., Monerris, J., Mau, R.L., Morici, E., Naseri, K., Ospina, A., Polo, V., Prina, A., Pucheta, E., Ramírez-Collantes, D.A., Romão, R., Tighe, M., Torres-Díaz, C., Val, J., Veiga, J.P., Wang, D., and Zaady, E., 2012, Plant species richness and ecosystem multifunctionality in global drylands: Science, v. 35, no. 6065, p. 214–218, accessed January 2024 at https://doi.org/10.1126/science.1215442.
Maestre, F.T., Salguero-Gómez, R., and Quero, J.L., 2012, It is getting hotter in here—Determining and projecting the impacts of global environmental change on drylands: Philosophical Transactions of the Royal Society B—Biological Sciences, v. 367, p. 3062–3075, accessed January 2024 at https://doi.org/10.1098/rstb.2011.0323.
Maher, A.T., Tanaka, J.A., and Rimbey, N., 2013, Economic risks of cheatgrass invasion on a simulated Eastern Oregon ranch: Rangeland Ecology & Management, v. 66, no. 3, p. 356–363, accessed January 2024 at https://doi.org/10.2111/REM-D-10-00126.1.
Maier, A.M., Perryman, B.L., Olson, R.A., and Hild, A.L., 2001, Climatic influences on recruitment of 3 subspecies of Artemisia tridentata: Journal of Range Management, v. 54, no. 6, p. 699–703, accessed January 2024 at https://doi.org/10.2307/4003674.
Mantyka-pringle, C.S., Martin, T.G., and Rhodes, J.R., 2012, Interactions between climate and habitat loss effects on biodiversity—A systematic review and meta-analysis: Global Change Biology, v. 18, no. 4, p. 1239–1252, accessed January 2024 at https://doi.org/10.1111/j.1365-2486.2011.02593.x.
Martel, J.-L., Brissette, F., Troin, M., Arsenault, R., Chen, J., Su, T., and Lucas-Picher, P., 2022, CMIP5 and CMIP6 model projection comparison for hydrological impacts over North America: Geophysical Research Letters, v. 49, no. 15, art. e2022GL098364, 11 p., accessed January 2024 at https://doi.org/10.1029/2022GL098364.
Maurer, G.E., Hallmark, A.J., Brown, R.F., Sala, O.E., and Collins, S.L., 2020, Sensitivity of primary production to precipitation across the United States: Ecology Letters, v. 23, no. 3, p. 527–536, accessed January 2024 at https://doi.org/10.1111/ele.13455.
Maxwell, S.L., Butt, N., Maron, M., McAlpine, C.A., Chapman, S., Ullmann, A., Segan, D.B., and Watson, J.E M., 2019, Conservation implications of ecological responses to extreme weather and climate events: Diversity and Distributions, v. 25, no. 4, p. 613–625, accessed January 2024 at https://doi.org/10.1111/ddi.12878.
McNaughton, S.J., Oesterheld, M., Frank, D.A., and Williams, K.J., 1989, Ecosystem-level patterns of primary productivity and herbivory in terrestrial habitats: Nature, v. 341, p. 142–144, accessed January 2024 at https://doi.org/10.1038/341142a0.
Merkle, J.A., Monteith, K.L., Aikens, E.O., Hayes, M.M., Hersey, K.R., Middleton, A.D., Oates, B.A., Sawyer, H., Scurlock, B.M., and Kauffman, M.J., 2016, Large herbivores surf waves of green-up during spring: Proceedings of the Royal Society B—Biological Sciences, v. 283, no. 1833, 8 p., accessed January 2024 at https://doi.org/10.1098/rspb.2016.0456.
Miller, R., Svejcar, T., West, N., 1994, Implications of livestock grazing in the intermountain sagebrush region: plant composition, in Vavra, M., Laycock, W.A., and Pieper, R.D., eds., Ecological implications of herbivory in the West—Society for Range Management, Denver, Colo., December 22, 1995: Denver, Colo., Society for Range Management, p. 101–146, accessed January 2024 at https://www.cabidigitallibrary.org/doi/full/10.5555/19950713826.
Molvar, E.M., Rosentreter, R., Mansfield, D., and Anderson, G.M., 2024, Cheatgrass invasions—History, causes, consequences, and solutions: Hailey, Idaho, Western Watersheds Project, 128 p., accessed January 2024 at https://www.westernwatersheds.org/wp-content/uploads/2024/09/Cheatgrass-Report.pdf.
Monroe, M.C., Plate, R.R., Oxarart, A., Bowers, A., and Chaves, W.A., 2019, Identifying effective climate change education strategies—A systematic review of the research: Environmental Education Research, v. 25, p. 791–812, accessed January 2024 at https://doi.org/10.1080/13504622.2017.1360842.
Morgan, J.A., LeCain, D.R., Pendall, E., Blumenthal, D.M., Kimball, B.A., Carrillo, Y., Williams, D.G., Heisler-White, J., Dijkstra, F.A., and West, M., 2011, C4 grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland: Nature, v. 476, p. 202–205, accessed January 2024 at https://doi.org/10.1038/nature10274.
Munson, S.M., Belnap, J., Schelz, C.D., Moran, M., and Carolin, T.W., 2011, On the brink of change—Plant responses to climate on the Colorado Plateau: Ecosphere, v. 2, no. 6, art. 68, 15 p., accessed January 2024 at https://doi.org/10.1890/ES11-00059.1.
Munson, S.M., Muldavin, E.H., Belnap, J., Peters, D.P.C., Anderson, J.P., Reiser, M.H., Gallo, K., Melgoza-Castillo, A., Herrick, J.E., and Christiansen, T.A., 2013, Regional signatures of plant response to drought and elevated temperature across a desert ecosystem: Ecology, v. 94, no. 9, p. 2030–2041, accessed January 2024 at https://doi.org/10.1890/12-1586.1.
Munson, S.M., and Long, A L., 2017, Climate drives shifts in grass reproductive phenology across the Western USA: New Phytologist, v. 213, no. 4, p. 1945–1955, accessed January 2024 at https://doi.org/10.1111/nph.14327.
Muscha, J.M., and Hild, A.L., 2006, Biological soil crusts in grazed and ungrazed Wyoming sagebrush steppe: Journal of Arid Environments, v. 67, no. 2, p. 195–207, accessed January 2024 at https://doi.org/10.1016/j.jaridenv.2006.02.010.
O’Neill, B.C., Tebaldi, C., van Vuuren, D.P., Eyring, V., Friedlingstein, P., Hurtt, G., Knutti, R., Kriegler, E., Lamarque, J.-F., Lowe, J., Meehl, G.A., Moss, R., Riahi, K., and Sanderson, B.M., 2016, The Scenario Model Intercomparison Project (ScenarioMIP) for CMIP6: Geoscientific Model Development, v. 9, no. 9, p. 3461–3482, accessed January 2024 at https://doi.org/10.5194/gmd-9-3461-2016.
Palmquist, K.A., Schlaepfer, D.R., Bradford, J.B., and Lauenroth, W.K., 2016, Mid-latitude shrub steppe plant communities—Climate change consequences for soil water resources: Ecology, v. 97, no. 9, p. 2342–2354, accessed January 2024 at https://doi.org/10.1002/ecy.1457.
Palmquist, K.A., Renne, R.R., Schlaepfer, D.R., Lauenroth, W.K., and Bradford, J.B., 2022, High-resolution maps of projected big sagebrush plant community biomass for 52 future climate scenarios using multivariate matching algorithms: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P9DR9G1Y.
Palmquist, K.A., Schlaepfer, D.R., Renne, R.R., Torbit, S.C., Doherty, K.E., Remington, T.E., Watson, G., Bradford, J.B., and Lauenroth, W.K., 2021, Divergent climate change effects on widespread dryland plant communities driven by climatic and ecohydrological gradients: Global Change Biology, v. 27, no. 9, p. 5169–5185, accessed January 2024 at https://doi.org/10.1111/gcb.15776.
Parmesan, C., Root, T.L., and Willig, M.R., 2000, Impacts of extreme weather and climate on terrestrial biota: Bulletin of the American Meteorological Society, v. 81, no. 3, p. 443–450, accessed January 2024 at hhttps://journals.ametsoc.org/view/journals/bams/81/3/1520-0477_2000_081_0443_ioewac_2_3_co_2.xml.
Pastick, N.J., Wylie, B.K., Rigge, M.B., Dahal, D., Boyte, S.P., Jones, M.O., Allred, B.W., Parajuli, S., and Wu, Z., 2021, Rapid monitoring of the abundance and spread of exotic annual grasses in the Western United States using remote sensing and machine learning: AGU Advances, v. 2, no. 2, art. e2020AV000298, 22 p., accessed January 2024 at https://doi.org/10.1029/2020AV000298.
Pennington, V.E., Bradford, J.B., Palmquist, K.A., Renne, R.R., and Lauenroth, W.K., 2019, Patterns of big sagebrush plant community composition and stand structure in the Western United States: Rangeland Ecology & Management, v. 72, no. 3, p. 505–514, accessed January 2024 at https://doi.org/10.1016/j.rama.2018.11.013.
Pennington, V.E., Schlaepfer, D.R., Beck, J.L., Bradford, J.B., Palmquist, K.A., and Lauenroth, W.K., 2016, Sagebrush, greater sage-grouse, and the occurrence and importance of forbs: Western North American Naturalist, v. 76, p. 298–312, accessed January 2024 at https://doi.org/10.3398/064.076.0307.
Perfors, T., Harte, J., and Alter, S.E., 2003, Enhanced growth of sagebrush (Artemisia tridentata) in response to manipulated ecosystem warming: Global Change Biology, v. 9, no. 5, p. 736–742, accessed January 2024 at https://doi.org/10.1046/j.1365-2486.2003.00559.x.
Peterson St-Laurent, G., Oakes, L.E., Cross, M., and Hagerman, S., 2021, R–R–T (resistance–resilience–transformation) typology reveals differential conservation approaches across ecosystems and time: Communications Biology, v. 4, p. 1–9, accessed January 2024 at https://doi.org/10.1038/s42003-020-01556-2.
Pilliod, D.S., Welty, J.L., and Arkle, R.S., 2017, Refining the cheatgrass–fire cycle in the Great Basin: Precipitation timing and fine fuel composition predict wildfire trends: Ecology and Evolution, v. 7, no. 19, p. 8126–8151, accessed January 2024 at https://doi.org/10.1002/ece3.3414.
Reich, P.B., Hobbie, S.E., Lee, T.D., and Pastore, M.A., 2018, Unexpected reversal of C3 versus C4 grass response to elevated CO2 during a 20-year field experiment: Science, v. 360, no. 6386, p. 317–320, accessed January 2024 at https://doi.org/10.1126/science.aas9313.
Remington, T.E., Deibert, P.A., Hanser, S.E., Davis, D.M., Robb, L.A., and Welty, J.L., 2021, Sagebrush conservation strategy—Challenges to sagebrush conservation: U.S. Geological Survey Open-File Report 2020–1125, 327 p., accessed January 2024 at https://doi.org/10.3133/ofr20201125.
Renne, R.R., Bradford, J.B., Burke, I.C., and Lauenroth, W.K., 2019, Soil texture and precipitation seasonality influence plant community structure in North American temperate shrub steppe: Ecology, v. 100, no. 11, art. e02824, 12 p., accessed January 2024 at https://doi.org/10.1002/ecy.2824.
Renne, R.R., Schlaepfer, D.R., Palmquist, K.A., Bradford, J.B., Burke, I.C., and Lauenroth, W.K., 2019, Soil and stand structure explain shrub mortality patterns following global change–type drought and extreme precipitation: Ecology, 100, no. 12, art. e02889, 17 p., accessed January 2024 at https://doi.org/10.1002/ecy.2889.
Renwick, K.M., Curtis, C., Kleinhesselink, A.R., Schlaepfer, D., Bradley, B.A., Aldridge, C.L., Poulter, B., and Adler, P.B., 2018, Multi-model comparison highlights consistency in predicted effect of warming on a semi-arid shrub: Global Change Biology, v. 24, no. 1, p. 424–438, accessed January 2024 at https://doi.org/10.1111/gcb.13900.
Richards, J.H., and Caldwell, M.M., 1987, Hydraulic lift: Substantial nocturnal water transport between soil layers by Artemisia tridentata roots: Oecologia, v. 73, p. 486–489, accessed January 2024 at https://doi.org/10.1007/BF00379405.
Rigge, M.B., Bunde, B., Shi, H., Postma, K., 2021, Rangeland Condition Monitoring Assessment and Projection (RCMAP) fractional component time-series across the Western U.S. 1985–2020 (ver. 2.0, October 2021): U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P95IQ4B.
Rigge, M., Postma, K., Bunde, B., and Shi, H., 2023, Projections of rangeland fractional component cover across Western US rangelands for RCP4.5 and RCP8.5 scenarios for the 2020s, 2050s, and 2080s time-periods: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P9J490BH.
Rigge, M., Shi, H., and Postma, K., 2021, Projected change in rangeland fractional component cover across the sagebrush biome under climate change through 2085: Ecosphere, v. 12, no. 6, art. e03538, 25 p., accessed January 2024 at https://doi.org/10.1002/ecs2.3538.
Roundy, B.A., Chambers, J.C., Pyke, D.A., Miller, R.F., Tausch, R.J., Schupp, E.W., Rau, B., and Gruell, T., 2018, Resilience and resistance in sagebrush ecosystems are associated with seasonal soil temperature and water availability: Ecosphere, v. 9, no. 9, art. e02417, 27 p., accessed January 2024 at https://doi.org/10.1002/ecs2.2417.
Sala, O.E., and Lauenroth, W.K., 1982, Small rainfall events—An ecological role in semiarid regions: Oecologia, v. 53, p. 301–304, accessed January 2024 at https://doi.org/10.1007/BF00389004.
Schlaepfer, D.R., and Bradford, J.B., 2021, Simulated rangewide big sagebrush regeneration estimates and relationships with abiotic variables as function of soils under historical and future climate projections: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P9MB2QB8.
Schlaepfer, D.R., Bradford, J.B., Lauenroth, W.K., and Shriver, R.K., 2021, Understanding the future of big sagebrush regeneration—Challenges of projecting complex ecological processes: Ecosphere, v. 12, no. 8, art. e03695, 26 p., accessed January 2024 at https://doi.org/10.1002/ecs2.3695.
Schlaepfer, D.R., Lauenroth, W.K., and Bradford, J.B., 2012a, Consequences of declining snow accumulation for water balance of mid-latitude dry regions: Global Change Biology, v. 18, no. 6, p. 1988–1997, accessed January 2024 at https://doi.org/10.1111/j.1365-2486.2012.02642.x.
Schlaepfer, D.R., Lauenroth, W.K., and Bradford, J.B., 2012b, Ecohydrological niche of sagebrush ecosystems: Ecohydrology, v. 5, no. 4, p. 453–466, accessed January 2024 at https://doi.org/10.1002/eco.238.
Schlaepfer, D.R., Taylor, K.A., Pennington, V.E., Nelson, K.N., Martyn, T.E., Rottler, C.M., Lauenroth, W.K., and Bradford, J.B., 2015, Simulated big sagebrush regeneration supports predicted changes at the trailing and leading edges of distribution shifts: Ecosphere, v. 6, no. 1, art. 3, 31 p., accessed January 2024 at https://doi.org/10.1890/ES14-00208.1.
Schlaepfer, D.R., Chambers, J.C., Urza, A.K., Hanberry, B.B., Brown, J.L., Board, D.I., Campbell, S.B., Clause, K.J., Crist, M.R., and Bradford, J.B., 2024, Declining ecological resilience and invasion resistance under climate change in the sagebrush region, United States: Ecological Applications, art. e3065, 22 p., accessed January 7, 2025, at https://doi.org/10.1002/eap.3065.
Schuurman G.W., Hawkins-Hoffman, C., Cole, D.N., Lawrence, D.J., Morton, J.M., Magness, D.R., Cravens, A.E., Covington, S., O'Malley, R., and Fisichelli, N.A., 2020, Resist-accept-direct (RAD)—A framework for the 21st-century natural resource manager: Fort Collins, Colo., National Park Service Natural Resource Report NPS/NRSS/CCRP/NRR—2020/2213, 20 p., accessed January 2024 at https://doi.org/10.36967/nrr-2283597.
Schuurman, G.W., Cole, D.N., Cravens, A.E., Covington, S., Crausbay, S.D., Hoffman, C.H., Lawrence, D.J., Magness, D.R., Morton, J.M., Nelson, E.A., and O’Malley, R., 2022, Navigating ecological transformation—Resist–accept–direct as a path to a new resource management paradigm: BioScience, v. 72, no. 1, p. 16–29, accessed January 2024 at https://doi.org/10.1093/biosci/biab067.
Seleiman, M.F., Al-Suhaibani, N., Ali, N., Akmal, M., Alotaibi, M., Refay, Y., Dindaroglu, T., Abdul-Wajid, H.H., and Battaglia, M.L., 2021, Drought stress impacts on plants and different approaches to alleviate its adverse effects: Plants, v. 10, no. 2, art. 259, 25 p., accessed January 2024 at https://doi.org/10.3390/plants10020259.
Shafer, S.L., Bartlein, P.J., and Thompson, R.S., 2001, Potential changes in the distributions of Western North America tree and shrub taxa under future climate scenarios: Ecosystems, v. 4, p. 200–215, accessed January 2024 at https://doi.org/10.1007/s10021-001-0004-5.
Shi, H., Homer, C., Rigge, M., Postma, K., and Xian, G., 2020, Analyzing vegetation change in a sagebrush ecosystem using long-term field observations and Landsat imagery in Wyoming: Ecosphere, v. 11, no. 12, art. e03311, 20 p., accessed January 2024 at https://doi.org/10.1002/ecs2.3311.
Shi, H., Rigge, M., Homer, C.G., Xian, G., Meyer, D.K., and Bunde, B., 2018, Historical cover trends in a sagebrush steppe ecosystem from 1985 to 2013—Links with climate, disturbance, and management: Ecosystems, v. 21, p. 913–929, accessed January 2024 at https://doi.org/10.1007/s10021-017-0191-3.
Shinneman, D.J., Baker, W.L., Shinneman, D.J., and Baker, W.L., 2009, Environmental and climatic variables as potential drivers of post-fire cover of cheatgrass (Bromus tectorum) in seeded and unseeded semiarid ecosystems: International Journal of Wildland Fire, v. 18, no. 2, p. 191–202, accessed January 2024 at https://doi.org/10.1071/WF07043.
Shinneman, D.J., Strand, E.K., Pellant, M., Abatzoglou, J.T., Brunson, M.W., Glenn, N.F., Heinrichs, J.A., Sadegh, M., and Vaillant, N.M., 2023, Future direction of fuels management in sagebrush rangelands: Rangeland Ecology & Management, v. 86, p. 50–63, accessed January 2024 at https://doi.org/10.1016/j.rama.2022.10.009.
Shriver, R.K., Andrews, C.M., Pilliod, D.S., Arkle, R.S., Welty, J.L., Germino, M.J., Duniway, M.C., Pyke, D.A., and Bradford, J.B., 2018, Adapting management to a changing world—Warm temperatures, dry soil, and interannual variability limit restoration success of a dominant woody shrub in temperate drylands: Global Change Biology, v. 24, no. 10, p. 4972–4982, accessed January 2024 at https://doi.org/10.1111/gcb.14374.
Shyvers, J.E., Tarbox, B.C., Van Schmidt, N.D., Saher, D.J., Heinrichs, J.A., and Aldridge, C.L., 2022, Database of invasive annual grass spatial products for the Western United States January 2010 to February 2021: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P9VW97AO.
Simler-Williamson, A.B., Applestein, C., and Germino, M.J., 2022, Interannual variation in climate contributes to contingency in post-fire restoration outcomes in seeded sagebrush steppe: Conservation Science and Practice, v. 4, no. 7, art. e12737, 15 p., accessed January 2024 at https://doi.org/10.1111/csp2.12737.
Skinner, R.H., Hanson, J.D., Hutchinson, G.L., and Schuman, G.E., 2002, Response of C3 and C4 grasses to supplemental summer precipitation: Journal of Range Management, v. 55, no. 5, p. 517–522, accessed January 2024 at https://doi.org/10.2307/4003232.
Smith, J.T., Allred, B.W., Boyd, C.S., Davies, K.W., Kleinhesselink, A.R., Morford, S.L., and Naugle, D.E., 2023, Fire needs annual grasses more than annual grasses need fire: Biological Conservation, v. 286, art. 110299, 9 p., accessed January 2024 at https://doi.org/10.1016/j.biocon.2023.110299.
Smith, M.D., 2011, An ecological perspective on extreme climatic events—A synthetic definition and framework to guide future research: Journal of Ecology, v. 99, no. 3, p. 656–663, accessed January 2024 at https://doi.org/10.1111/j.1365-2745.2011.01798.x.
Smull, D.M., Pendleton, N., Kleinhesselink, A.R., and Adler, P.B., 2019, Climate change, snow mold and the Bromus tectorum invasion—Mixed evidence for release from cold weather pathogens: AoB PLANTS, v. 11, no. 5, art. plz043, 9 p., accessed January 2024 at https://doi.org/10.1093/aobpla/plz043.
Stein, B., Glick, P., Edelson, N.A., and Staudt, A., 2014, Climate-smart conservation: putting adaption principles into practice: Washington D.C., National Wildlife Federation, 262 p., accessed January 2024 at https://pubs.usgs.gov/publication/70093621.
Still, S.M., and Richardson, B.A., 2015, Projections of contemporary and future climate niche for Wyoming big sagebrush (Artemisia tridentata subsp. wyomingensis)—A guide for restoration: Natural Areas Journal, v. 35, no. 1, p. 30–43, accessed January 2024 at https://doi.org/10.3375/043.035.0106.
Tarbox, B.C., Van Schmidt, N.D., Shyvers, J.E., Saher, D.J., Heinrichs, J.A., and Aldridge, C.L., 2022, Bridging the gap between spatial modeling and management of invasive annual grasses in the imperiled sagebrush biome: Rangeland Ecology & Management, v. 82, p. 104–115, accessed January 2024 at https://doi.org/10.1016/j.rama.2022.01.006.
Thorarinsdottir, T.L., Sillmann, J., Haugen, M.,Gissibl, N., and Sandstad, M., 2020, Evaluation of CMIP5 and CMIP6 simulations of historical surface air temperature extremes using proper evaluation methods: Environmental Research Letters, v. 15, art. 124041, 13 p., accessed January 2024 at https://doi.org/10.1088/1748-9326/abc778.
Tredennick, A.T., Hooten, M.B., Aldridge, C.L., Homer, C.G., Kleinhesselink, A.R., and Adler, P.B., 2016, Forecasting climate change impacts on plant populations over large spatial extents: Ecosphere, v. 7, no. 10, 16 p., accessed January 2024 at https://doi.org/10.1002/ecs2.1525.
Tredennick, A.T., Kleinhesselink, A.R., Taylor, J.B., and Adler, P.B., 2018, Ecosystem functional response across precipitation extremes in a sagebrush steppe: PeerJ, v. 6, art. e4485, 19 p., accessed January 2024 at https://doi.org/10.7717/peerj.4485.
Tredennick, A.T., Monroe, A.P., Prebyl, T., Lombardi, J., and Aldridge, C.L., 2023a, Dynamic spatiotemporal modeling of a habitat-defining plant species to support wildlife management at regional scales: Ecosphere, v. 14, no. 6, art. e4534, 20 p., accessed January 2024 at https://doi.org/10.1002/ecs2.4534.
Tredennick, A.T., Monroe, A.P., Prebyl, T., Lombardi, J., and Aldridge, C.L., 2023b, Sagebrush projections for greater sage-grouse core areas in Wyoming, USA, 2018–2100: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P9G58V48.
Turco, M., von Hardenberg, J., AghaKouchak, A., Llasat, M.C., Provenzale, A., and Trigo, R.M., 2017, On the key role of droughts in the dynamics of summer fires in Mediterranean Europe: Scientific Reports, v. 7, art. 81, 10 p., accessed January 2024 at https://doi.org/10.1038/s41598-017-00116-9.
U.S. Department of the Interior, 2008, Adaptive management implementation policy: U.S. Department of the Interior Manual, chap. 1, accessed October 2024 at https://www.doi.gov/sites/doi.gov/files/elips/documents/522-dm-1.pdf.
U.S. Department of the Interior, 2023, Climate change policy: U.S. Department of the Interior Manual, chap. 1, accessed October 2024 at https://www.doi.gov/sites/doi.gov/files/elips/documents/523-dm-1.pdf.
U.S. Global Change Research Program, 2017, Climate science special report, v. 4 of Wuebbles, D.J., Fahey, D.W., Hibbard, K.A., Dokken, D.J., Stewart, B.C., and Maycock, T.K., eds., Fourth national climate assessment: Washington D.C., U.S. Global Change Research Program, 477 p., accessed January 2024 at https://www.globalchange.gov/reports/climate-science-special-report-fourth-national-climate-assessment-nca4-volume-i.
Valpine, P., de, and Harte, J., 2001, Plant responses to experimental warming in a montane meadow: Ecology, v. 82, no. 3, p. 637–648, accessed January 2024 at https://doi.org/10.1890/0012-9658(2001)082[0637:PRTEWI]2.0.CO;2.
Van Lanen, N.J., Monroe, A.P., and Aldridge, C.L., 2023, A hidden cost of single species management—Habitat-relationships reveal potential negative effects of conifer removal on a non-target species: Biological Conservation, v. 280, art. 109959, 10 p., accessed January 2024 at https://doi.org/10.1016/j.biocon.2023.109959.
Volder, A., Briske, D.D., and Tjoelker, M.G., 2013, Climate warming and precipitation redistribution modify tree–grass interactions and tree species establishment in a warm-temperate savanna: Global Change Biology, v. 19, no. 3, p. 843–857, accessed January 2024 at https://doi.org/10.1111/gcb.12068.
Weisberg, P.J., Lingua, E., and Pillai, R.B., 2007, Spatial patterns of pinyon–juniper woodland expansion in Central Nevada: Rangeland Ecology & Management, v. 60, no. 3, p. 115–124, accessed January 2024 at https://doi.org/10.2111/05-224R2.1.
Wiegand, K., Saltz, D., and Ward, D., 2006, A patch-dynamics approach to savanna dynamics and woody plant encroachment—Insights from an arid savanna: Perspectives in Plant Ecology, Evolution and Systematics, v. 7, no. 4, p. 229–242, accessed January 2024 at https://doi.org/10.1016/j.ppees.2005.10.001.
Williams, A.L., Wills, K.E., Janes, J.K., Vander Schoor, J.K., Newton, P.C.D., and Hovenden, M.J., 2007, Warming and free-air CO2 enrichment alter demographics in four co-occurring grassland species: New Phytologist, v. 176, no. 2, p. 365–374, accessed January 2024 at https://doi.org/10.1111/j.1469-8137.2007.02170.x.
Williamson, M.A., Fleishman, E., Mac Nally, R.C., Chambers, J.C., Bradley, B.A., Dobkin, D.S., Board, D.I., Fogarty, F.A., Horning, N., Leu, M., and Wohlfeil Zillig, M., 2020, Fire, livestock grazing, topography, and precipitation affect occurrence and prevalence of cheatgrass (Bromus tectorum) in the Central Great Basin, USA: Biological Invasions, v. 22, p. 663–680, accessed January 2024 at https://doi.org/10.1007/s10530-019-02120-8.
Winkler, D.E., Belnap, J., Hoover, D., Reed, S.C., and Duniway, M.C., 2019, Shrub persistence and increased grass mortality in response to drought in dryland systems: Global Change Biology, v. 25, no. 9, p. 3121–3135, accessed January 2024 at https://doi.org/10.1111/gcb.14667.
Xian, G., Homer, C.G., and Aldridge, C.L., 2012, Effects of land cover and regional climate variations on long-term spatiotemporal changes in sagebrush ecosystems: GIScience & Remote Sensing, v. 49, no. 3, p. 378–396, accessed January 2024 at https://doi.org/10.2747/1548-1603.49.3.378.
Zavaleta, E.S., Shaw, M.R., Chiariello, N.R., Thomas, B.D., Cleland, E.E., Field, C.B., and Mooney, H.A., 2003, Grassland responses to three years of elevated temperature, CO2, precipitation, and N deposition: Ecological Monographs, v. 73, no. 4, p. 585–604, accessed January 2024 at https://doi.org/10.1890/02-4053.
Zhang, F., Biederman, J.A., Dannenberg, M.P., Yan, D., Reed, S.C., and Smith, W.K., 2021, Five decades of observed daily precipitation reveal longer and more variable drought events across much of the Western United States: Geophysical Research Letters, v. 48, no. 7, art. e2020GL092293, 11 p., accessed January 2024 at https://doi.org/10.1029/2020GL092293.
Zimmer, S., Grosklos, G., Belmont, P., and Adler, P., 2020, Agreement and uncertainty among climate change impact models—A synthesis of sagebrush steppe vegetation predictions: HydroShare data release, accessed January 2024 at https://doi.org/10.4211/hs.e6b15828d20843eab4e2babd89787f41.
Zimmer, S.N., Grosklos, G.J., Belmont, P., and Adler, P.B., 2021, Agreement and uncertainty among climate change impact models—A synthesis of sagebrush steppe vegetation projections: Rangeland Ecology & Management, v. 75, p. 119–129, accessed January 2024 at https://doi.org/10.1016/j.rama.2020.12.006.
Appendix 1. Maps of Projected 21st Century Climate and Drought Conditions
-
• Mean climate
-
o Mean annual temperature (MAT; fig. 1.1A–I)
-
o Mean annual precipitation (PPT; fig. 1.2A–I)
-
-
• Interannual variability
-
o Standard deviation (SD) of annual temperature (MAT; fig. 1.3A–I)
-
o Coefficient of variation (CV) of annual precipitation (PPT; fig. 1.4A–I)
-
-
• Growing season
-
o Mean day of year (DOY) with last spring frost (the DOY during which the last exposure to frost occurs before the warm season; fig. 1.5A–I)
-
o Mean DOY with first fall frost (the DOY during which the first exposure to frost occurs after the warm season; fig. 1.6A–I)
-
o Mean growing season length (duration of the growing season; fig. 1.7A–I)
-
-
• Moisture seasonality
-
o Mean seasonal timing of PPT (precipitation seasonality, the correlation between monthly precipitation and air temperature; fig. 1.8A–I)
-
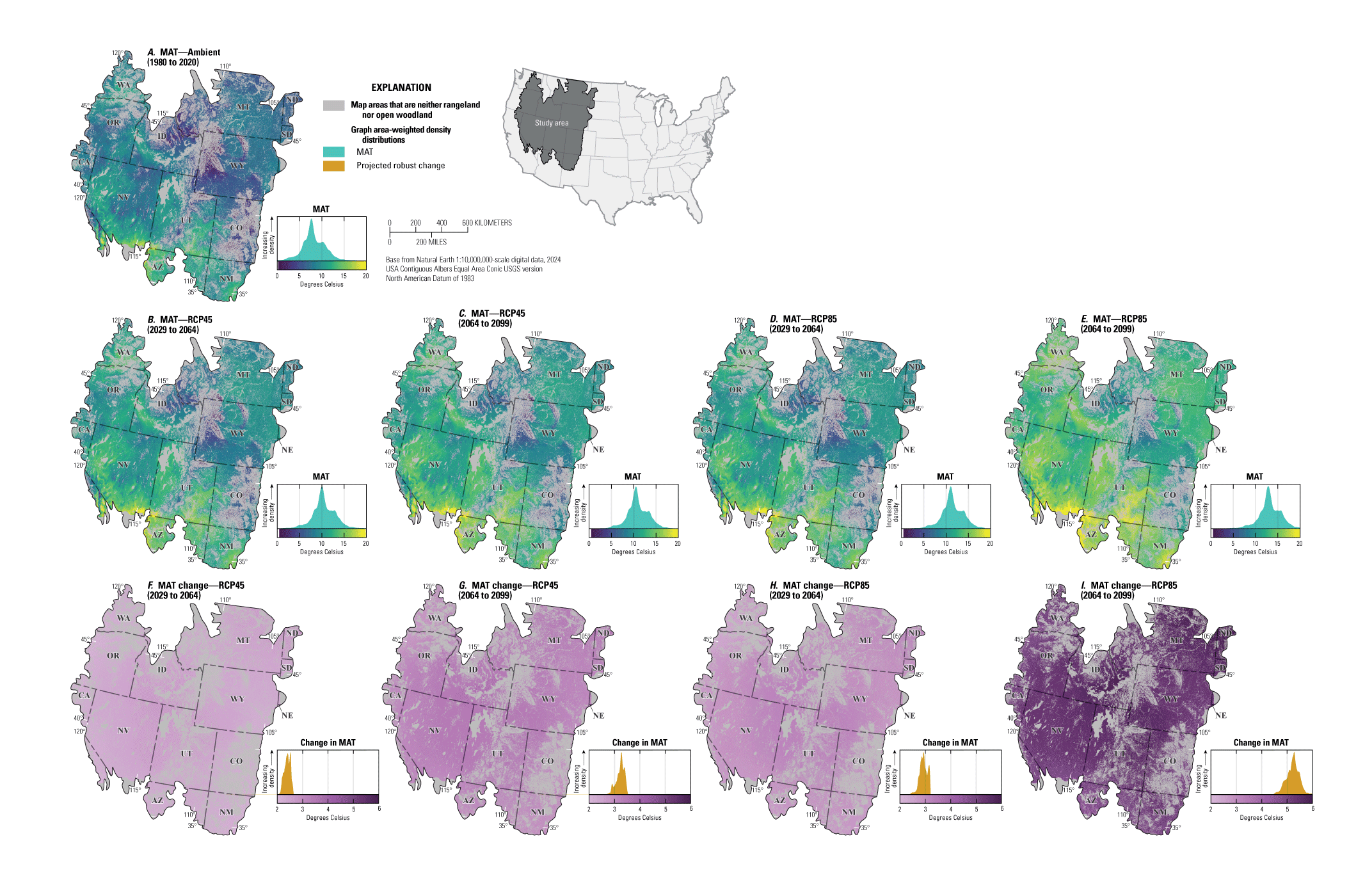
A–I, Maps showing the ambient, projected future, and projected changes in mean annual temperature (MAT; in degrees Celsius). Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019).
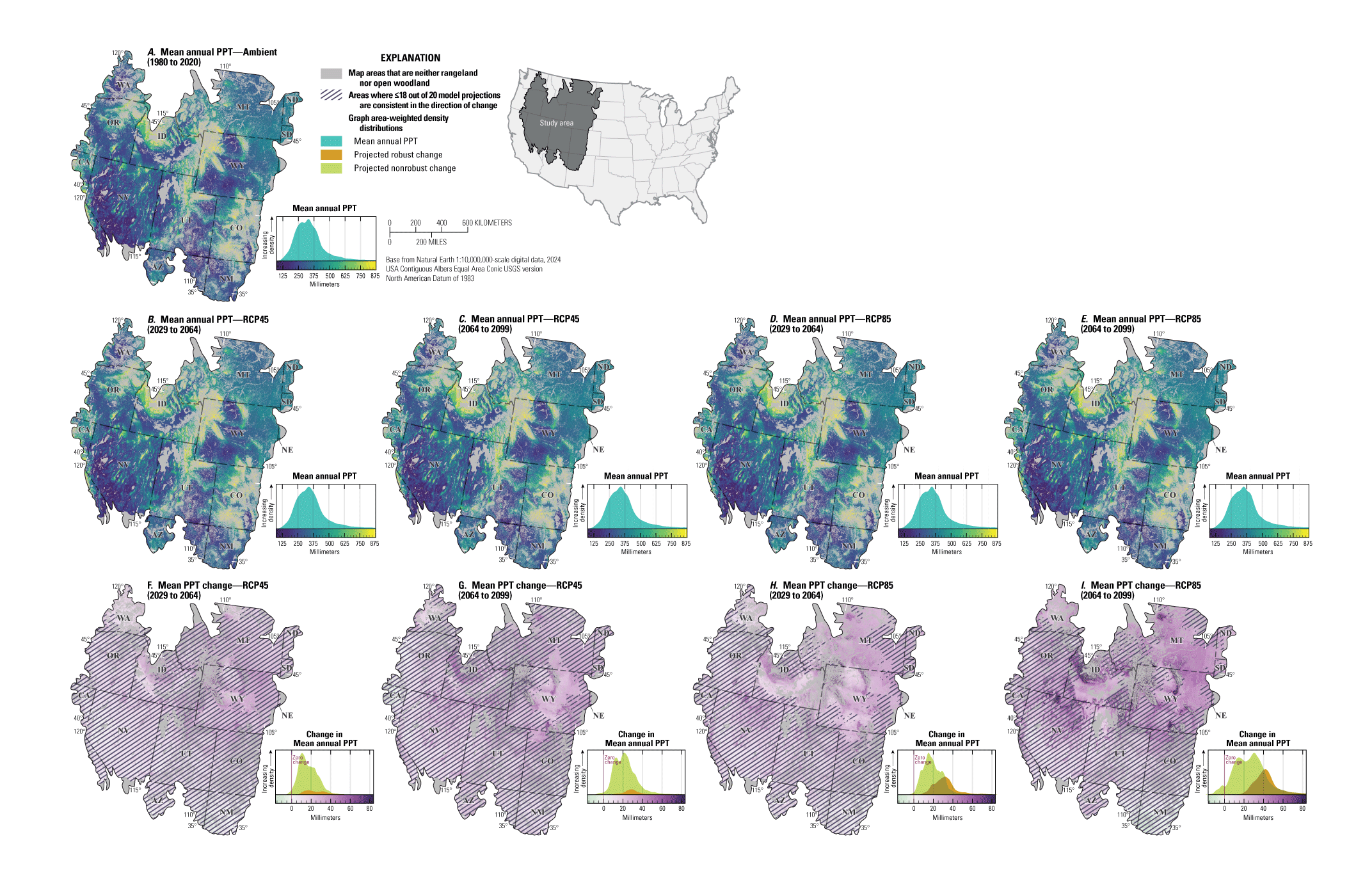
A–I, Maps showing historical, future and change in mean annual precipitation (PPT) amount (in millimeters). Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013) Sagebrush biome polygon from Jeffries and Finn (2019). ≤, less than or equal to
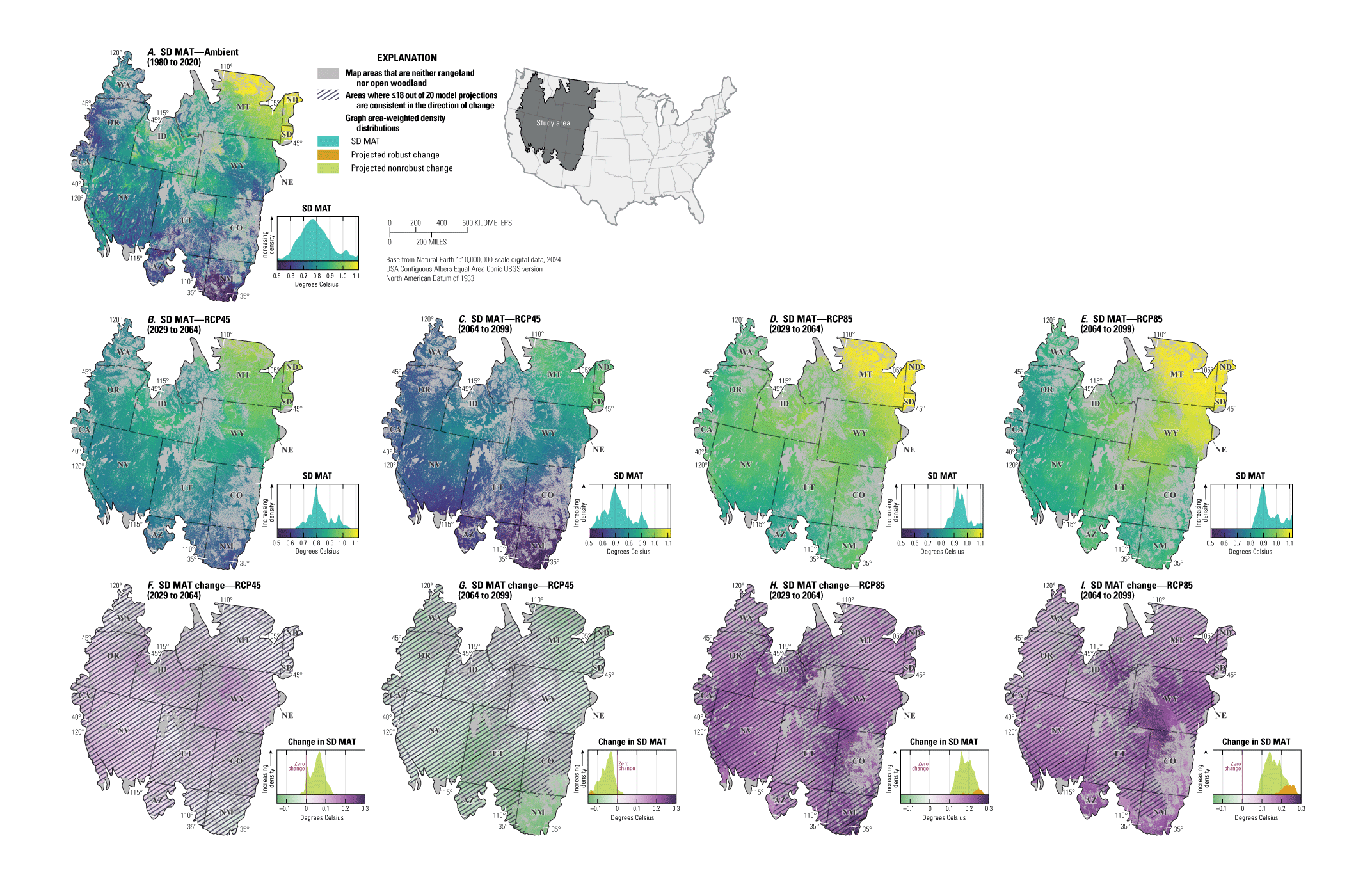
A–I, Maps showing historical, future and change in standard deviation (SD) of mean annual temperature (MAT; in degrees Celsuis). Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019). ≤, less than or equal to
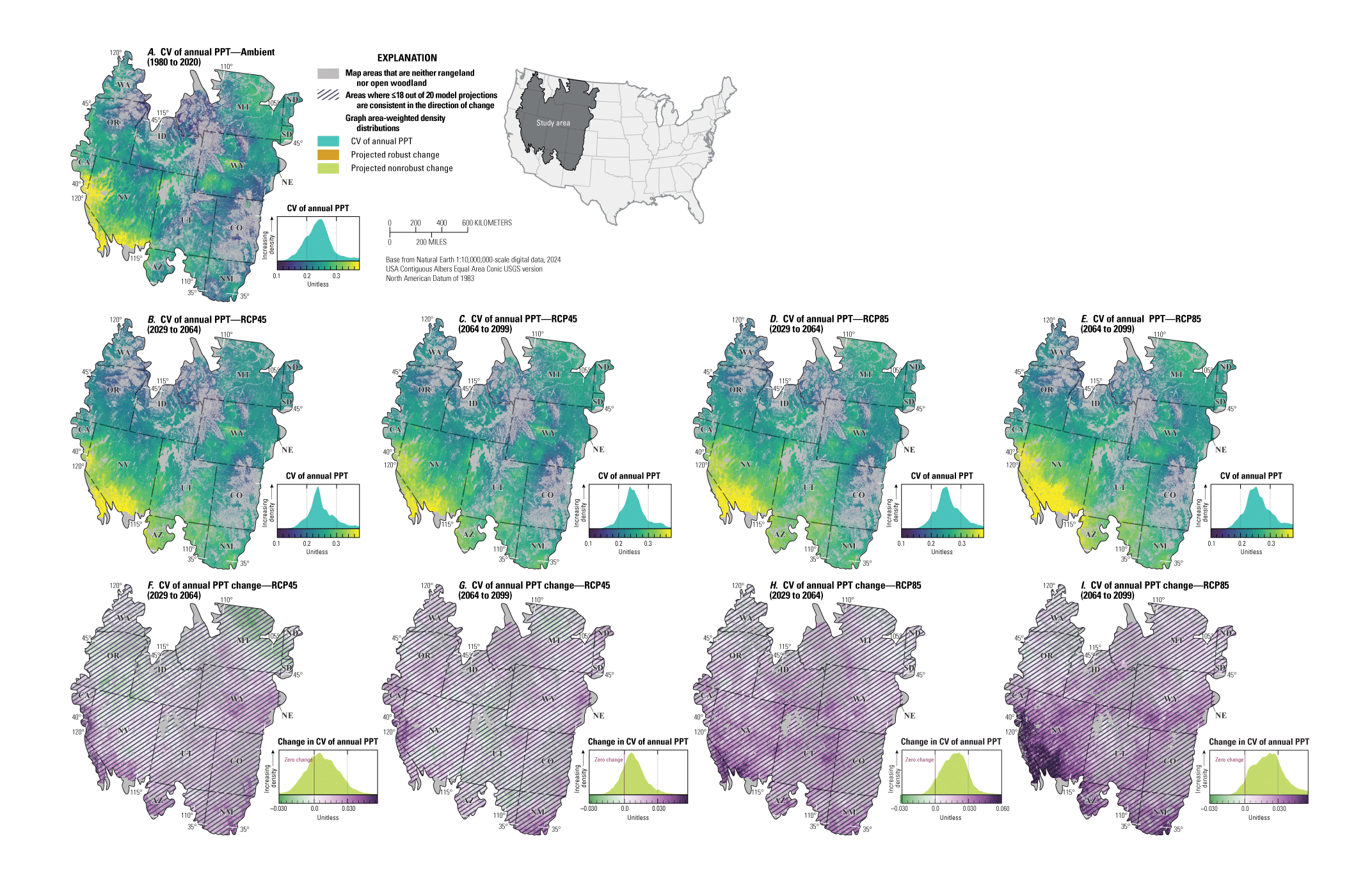
A–I, Maps showing historical, future and change in coefficient of variation (CV) of annual precipitation (PPT). Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019). ≤, less than or equal to
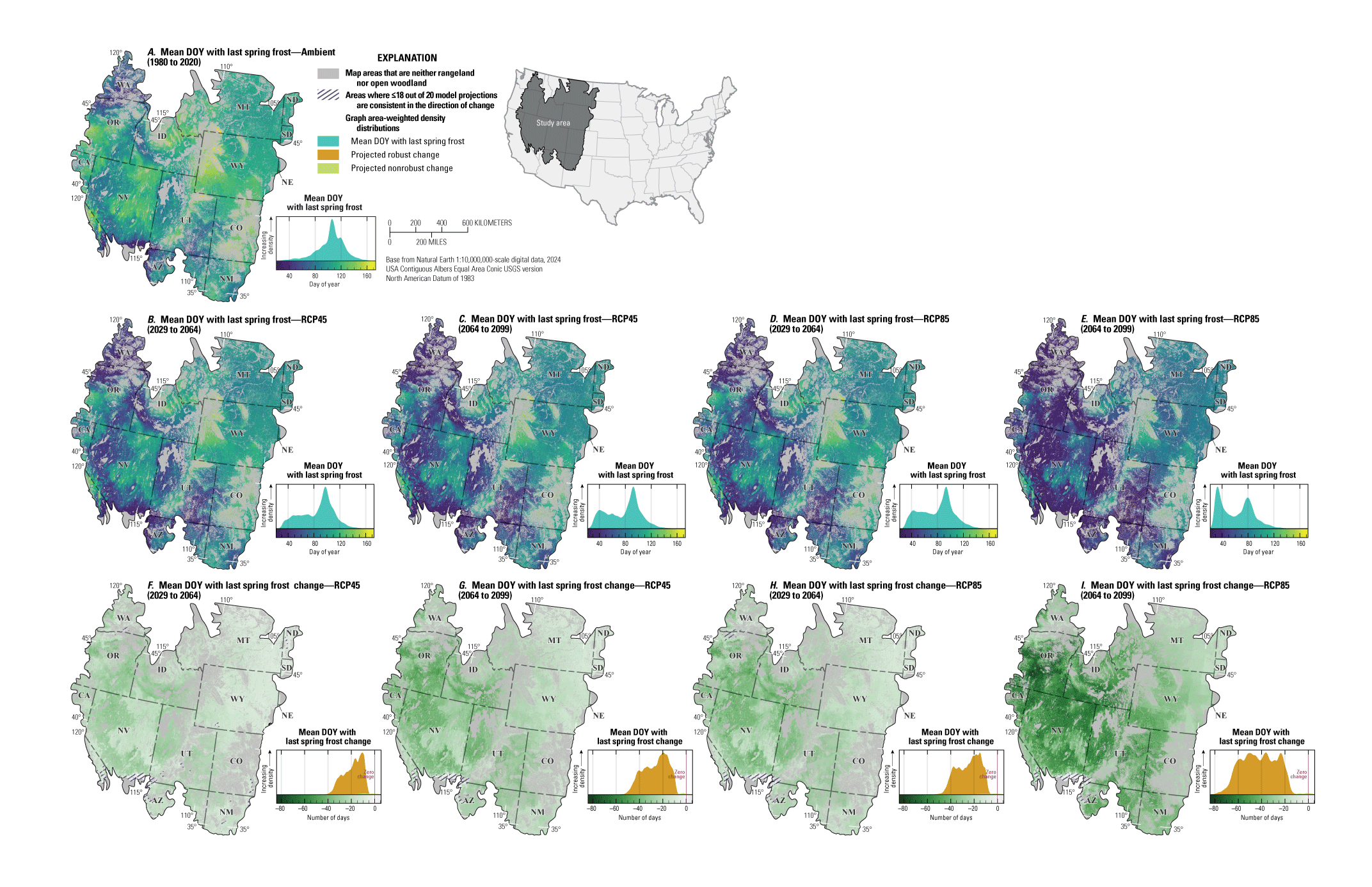
A–I, Maps showing historical, future and change in day of year (DOY) with the last spring frost. Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019). ≤, less than or equal to
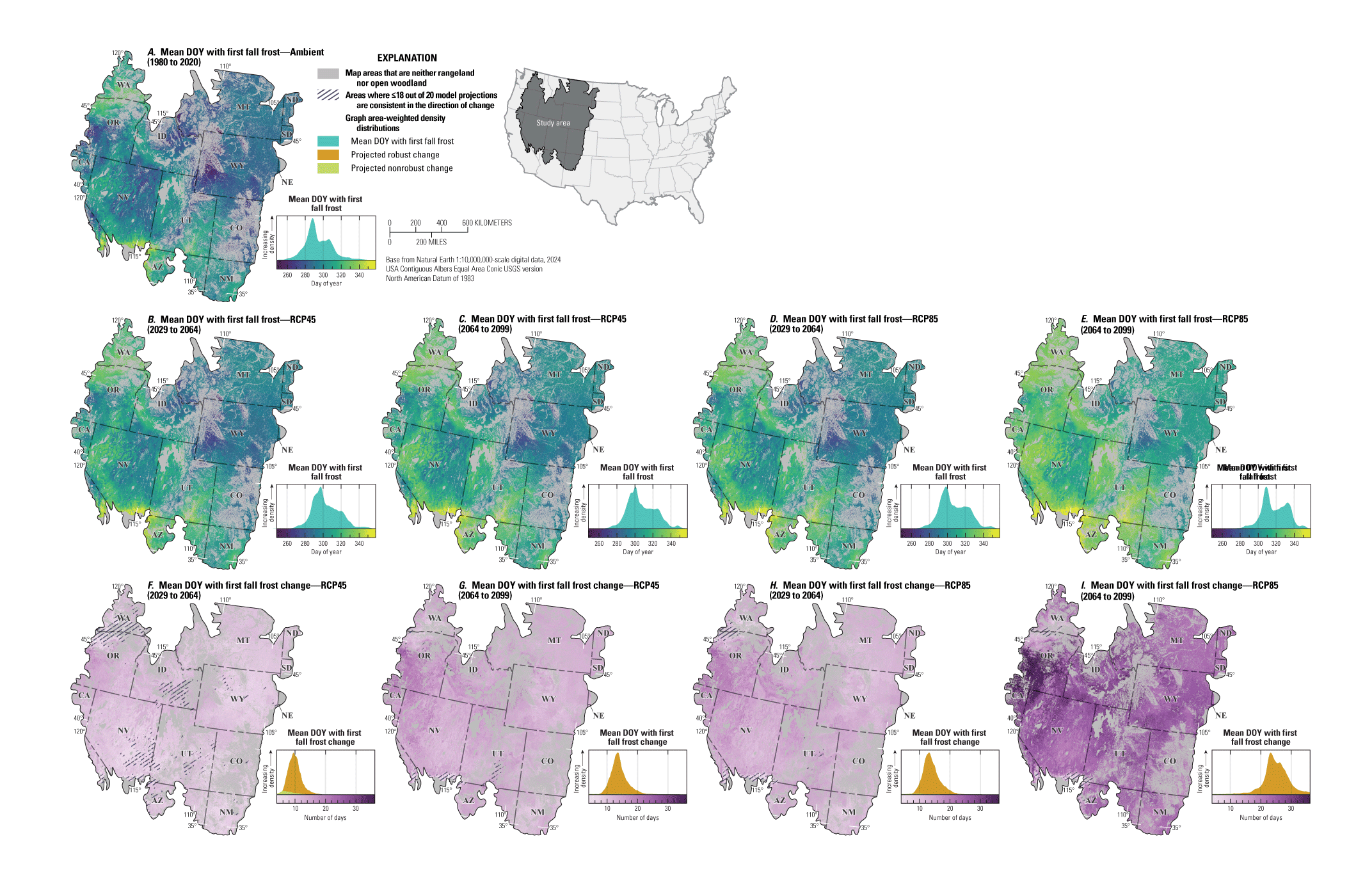
A–I, Maps showing historical, future and change in day of year (DOY) with the first fall frost. Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019). ≤, less than or equal to

A–I, Maps showing historical, future, and change in growing season length (in days). Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019).
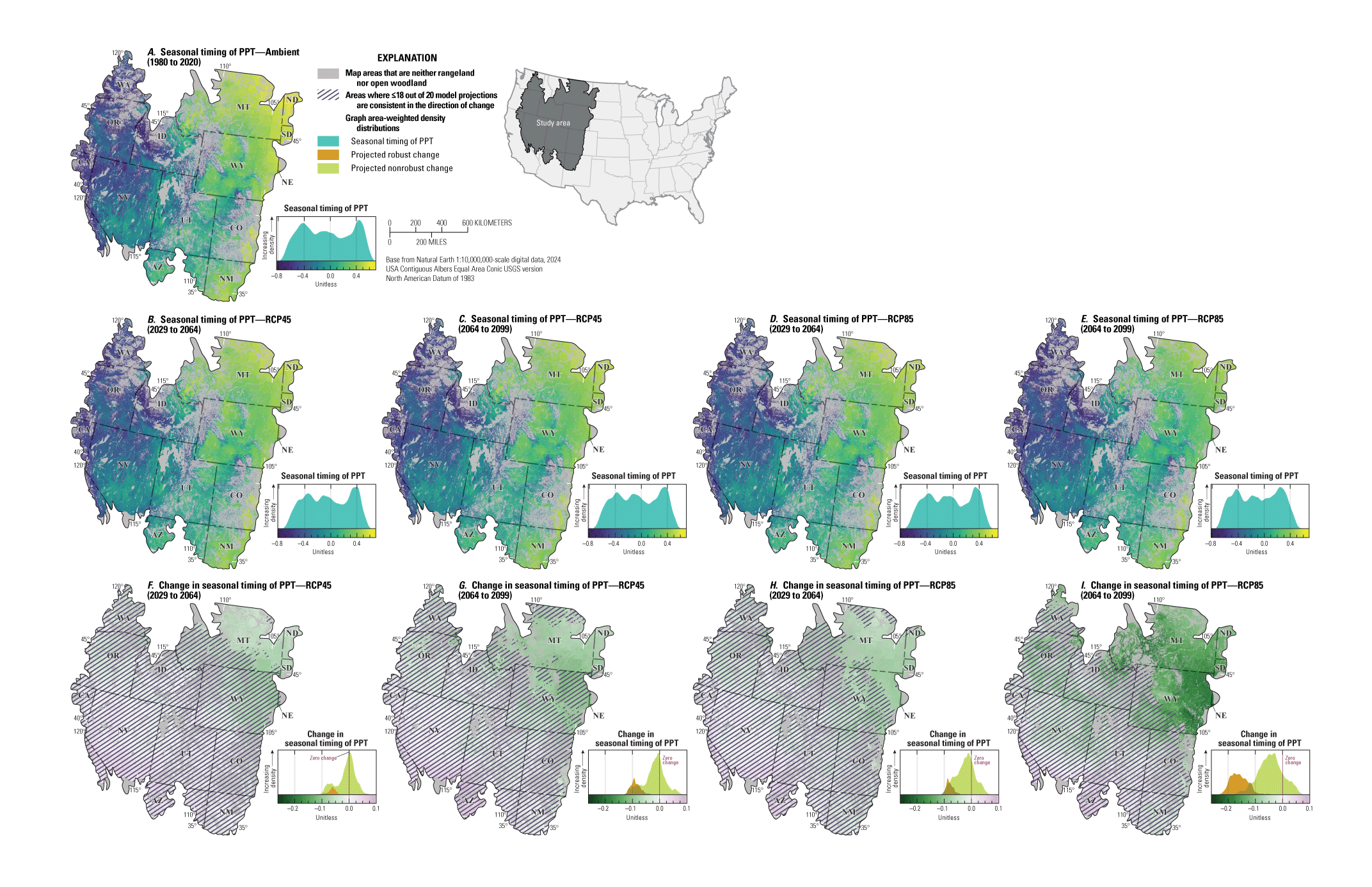
A–I, Maps showing historical, future and change in seasonal timing of precipitation, quantified as the correlation coefficient between mean monthly precipitation and mean monthly temperature. Ambient conditions include the years from 1980 to 2020 and future projections show median values and median change values across 20 global circulation models [GCM]s from the Coupled Model Intercomparison Project Phase 5 [CMIP5] representative concentration pathway [RCP] RCP4.5 and RCP8.5 for the years from 2029 to 2064 and from 2064 to 2099. Color scales show values between the minimum and the outlier-removed maximum (outliers are condensed, in other words, the maximum color represents values at the maximum and larger). Maps show the areas defined as “rangeland,” “open woodland,” and “other areas” (Chambers and others, 2023). Graphs on the right of each map show area-weighted density distributions of values across the entire region. Data modified from Abatzoglou and Brown (2012) and Abatzoglou (2013). Sagebrush biome polygon from Jeffries and Finn (2019). PPT, precipitation; ≤, less than or equal to
References Cited
Abatzoglou, J.T., 2013, Development of gridded surface meteorological data for ecological applications and modelling: International Journal of Climatology, v. 33, no. 1, p. 121–131, accessed January 2024 at https://doi.org/10.1002/joc.3413.
Abatzoglou, J.T., and Brown, T.J., 2012, A comparison of statistical downscaling methods suited for wildfire applications: International Journal of Climatology, v. 32, no. 5, p. 772–780, accessed January 2024 at https://doi.org/10.1002/joc.2312.
Chambers, J.C., Brown, J.L., Bradford, J.B., Board, D.I., Campbell, S.B., Clause, K.J., Hanberry, B., Schlaepfer, D.R., and Urza, A.K., 2023, New indicators of ecological resilience and invasion resistance to support prioritization and management in the sagebrush biome, United States: Frontiers in Ecology and Evolution, v. 10, 17 p., accessed January 2024 at https://doi.org/10.3389/fevo.2022.1009268.
Jeffries, M.I., and Finn, S.P., 2019, The sagebrush biome range extent, as derived from classified Landsat imagery: U.S. Geological Survey data release, accessed January 2024 at https://doi.org/10.5066/P950H8HS.
Abbreviations
>
greater than
AIM
Assessment, Inventory and Monitoring
C3
cool season
C4
warm season
CCRF
Climate Change Response Framework
CFR
Code of Federal Regulations
CO2
carbon dioxide
CMIP5
Coupled Model Intercomparison Project Phase 5
CSV
comma-separated values
CV
coefficient of variation
CWD
climate water deficit
DOY
day of year
GCM
global circulation models
GHG
greenhouse gas
GISSM
Germination and Individual Seedling Survival Model
HCCVI
Habitat Climate Change Vulnerability Index
IPCC
Intergovernmental Panel on Climate Change
MAT
mean annual temperature
NEPA
National Environmental Policy Act
netCDF
Network Common Data Form
PPT
precipitation
RAD
Resist-Accept-Direct
RCMAP
Rangeland Condition Monitoring Assessment and Projection
RCP
representative concentration pathway
SD
standard deviation
SDM
species distribution models
SEI
sagebrush ecological integrity
SSP
shared socioeconomic pathway
ssp.
subspecies
spp.
several species of
U.S.C.
United States Code
Publishing support provided by the Science Publishing Network, Denver and Lafayette Publishing Service Centers
For more information about this report, contact the
Director, USGS Southwest Biological Science Center
2255 N. Gemini Drive
Flagstaff, AZ 86001
https://www.usgs.gov/centers/southwest-biological-science-center
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
Carpenter, S.M., Holdrege, M.C., Schlaepfer, D.R., Phillips, J., Griffin, P., Lauenroth, W.K., and Bradford, J.B., 2025, Climate change impacts on plant communities in the sagebrush region—A science synthesis to inform Bureau of Land Management resource management: U.S. Geological Survey Scientific Investigations Report 2025–5045, 60 p., https://doi.org/10.3133/sir20255045.
ISSN: 2328-0328 (online)
Study Area
| Publication type | Report |
|---|---|
| Publication Subtype | USGS Numbered Series |
| Title | Climate change impacts on plant communities in the sagebrush region—A science synthesis to inform Bureau of Land Management resource management |
| Series title | Scientific Investigations Report |
| Series number | 2025-5045 |
| DOI | 10.3133/sir20255045 |
| Publication Date | July 15, 2025 |
| Year Published | 2025 |
| Language | English |
| Publisher | U.S. Geological Survey |
| Publisher location | Reston VA |
| Contributing office(s) | Southwest Biological Science Center |
| Description | x, 60 p. |
| Country | United States |
| State | Arizona, California, Colorado, Idaho, Montana, Nevada, New Mexico, North Dakota, Oregon, South Dakota, Utah, Washington, Wyoming |
| Online Only (Y/N) | Y |