

Potential Risks of Vegetation Treatments to Introduce and Increase Invasive Annual Grasses in Rangelands of Montana, North Dakota, and South Dakota—A Science Synthesis to Inform National Environmental Policy Act Analyses
Links
- Document: Report (4.69 MB pdf) , HTML , XML
- Related Works:
- Fact Sheet 2024-3028 Structured Science Syntheses to Inform Decision Making on Federal Public Lands
- Scientific Investigations Report 2024-5087 Effects of Noise from Oil and Gas Development on Raptors and Songbirds—A Science Synthesis to Inform National Environmental Policy Act Analyses
- Scientific Investigations Report 2023-5114 Effects of Noise from Oil and Gas Development on Ungulates and Small Mammals—A Science Synthesis to Inform National Environmental Policy Act Analyses
- Scientific Investigations Report 2023-5132 Effects of Culverts on Habitat Connectivity in Streams—A Science Synthesis to Inform National Environmental Policy Act Analyses
- Download citation as: RIS | Dublin Core
Acknowledgments
Funding for this work was provided by the Bureau of Land Management (BLM) Montana/Dakotas State Office. We thank the broader project team that advised on the development of this synthesis: Nathan J. Kleist (U.S. Geological Survey [USGS]), Elisabeth C. Teige (contractor to the USGS), Richard J. Lehrter (contractor to the BLM), Megan A. Gilbert (BLM), Claudia Mengelt (USGS), John C. Tull (U.S. Fish and Wildlife Service), and Travis S. Haby (BLM). We also thank Amy Symstad (USGS) and Cara Applestein (USGS) for providing independent peer reviews of this synthesis. Reyer Rens (BLM), Cory Neuharth (BLM), and Jason Sprung (BLM) provided courtesy reviews of the synthesis.
Executive Summary
Background: The U.S. Geological Survey is working with Federal land management agencies to develop a series of planned structured science syntheses to support environmental effects analyses that agencies conduct under the National Environmental Policy Act (NEPA). In this report, we synthesize science information relevant to environmental effects analyses concerned with the potential effects of proposed vegetation treatments to increase the distribution and abundance of invasive annual grasses (IAGs). This report does not focus on vegetation treatments specifically targeting IAGs, rather on understanding potential IAG responses following more general habitat restoration actions. The focal environments for this synthesis are rangelands in the intermontane valleys of Montana and the northern Great Plains of Montana, North Dakota, and South Dakota (hereafter referred to as “the focal region”).
How this report can inform a NEPA analysis: We organized the sections of this synthesis to align with the standard elements of an environmental effects analysis conducted under the NEPA, including characterizing a proposed action and alternatives (sec. 1), characterizing the affected environment (sec. 2), selecting methods for analyzing potential environmental effects (sec. 3), identifying issues and assessing the potential environmental effects for each issue (sec. 4), and identifying management practices that could inform project design and mitigation of potential adverse effects (sec. 5).
Vegetation treatments and assessing IAG distribution: Vegetation treatments in the focal region are typically one or a combination of prescribed fire, mechanical, hand-pulling, and herbicide treatments; alternative actions often considered in NEPA analyses are use of a single treatment type or no action. Alternative actions may include treatments applied at varying spatial scales, dosages, or in combination with other management actions such as grazing and seeding desirable species.
Key determinants to the susceptibility of rangelands to IAGs are climate and the native plant community. In addition, propagule pressure and land use history are important factors that affect how likely a system will be invaded by annual grasses. Data for IAGs are available for much of the focal region, as invasive species are included in standard vegetation sampling protocols and multiple approaches have been developed for predicting IAG presence from remotely sensed imagery. Locally collected, in-house data can supplement remote sensing products to evaluate their reliability and increase the understanding of current presence and abundance of IAGs on the landscape in NEPA analyses. Depending on available data, resources, and expertise, NEPA analyses of potential increases in IAGs from vegetation treatments may range from rapid, qualitative assessments to rigorous modeling of IAG responses in the project area under alternative management and climate scenarios.
Research on whether vegetation treatments increase IAGs and associated mitigation measures: The likelihood of increasing IAGs from vegetation treatments depends on treatment type and environmental context. In sagebrush ecosystems of the focal region, prescribed fire often reduces or does not increase IAGs. However, treatments that cause soil disturbances, such as mechanical removals of sagebrush or firebreak constructions, are more likely to increase IAGs than other treatments. Herbicides applied to reduce sagebrush cover have not increased the proportion of IAGs in the plant community.
In grasslands, prescribed fire often reduces IAGs, but effects depend on the abundance of native species and are often short lived. Mowing can increase or decrease IAGs in grassland ecosystems, whereas some herbicides can increase control of IAGs following fire. Grassland site conditions, such as southeast-facing slopes, sandier or rockier sites, or lower native species cover or richness, affect the likelihood of invasion by annual grasses.
Research on vegetation treatments and IAGs in sagebrush ecosystems specific to the focal region could enhance understanding of the potential effects of treatments from interactions among changing climate, IAGs, and native vegetation. Further research could also reveal the balance between the durability and frequency of prescribed fire needed to effectively control IAGs while avoiding other unintended vegetation community responses. Effectiveness of multiple treatment types and interactions with other management actions (for example, grazing) is also a research area that could be advanced.
Maintaining adequate cover of perennial vegetation creates rangelands that are resistant and resilient to annual grass invasions. Vegetation treatments can be a vector for invasion by IAGs, making it important to focus on treatment type, placement, and seasonal timing to limit invasion potential. Herbicides can provide effective mitigation, especially in combination with other controls such as prescribed fire or grazing.
Conclusion: This science synthesis can facilitate the use of the best available science in decisions about vegetation treatments on public lands in the intermontane valleys of Montana and the Great Plains of Montana, North Dakota, and South Dakota. This report can be incorporated by reference in NEPA documentation, included in a project record, or provide a general reference for understanding and identifying literature about increases in IAGs associated with vegetation treatments in rangelands in the region.
Methods for developing this synthesis: Rutherford and others (2023) introduced a methodology for developing science syntheses to inform analyses conducted under NEPA, and relevant text from that report is reproduced herein. This and other syntheses build on that foundation and methodology and apply it to new topics of management concern on western lands.We conducted a structured literature search of online databases to find science about the increases and mitigation of IAGs associated with vegetation treatments in rangelands of the focal region. Throughout this effort, we worked with staff from the Bureau of Land Management, U.S. Fish and Wildlife Service, and U.S. Geological Survey to coproduce this report.
Purpose of This Report
Federal land management agencies permit and plan for many uses and activities on public lands across the United States. Under the National Environmental Policy Act of 1969 (NEPA; 42 U.S.C. 4321 et seq.), Federal agencies must analyze and disclose the potential environmental effects of major Federal actions that may substantially affect the quality of the human environment. Regulations for implementing the NEPA require the integrated use of the natural and social sciences in agency planning and decision processes (40 CFR § 1501.2). Science is foundational to understanding how proposed Federal actions may affect natural resources, ecosystems, and human communities. Science syntheses can be a useful mechanism for sharing science information with public land managers to inform their decisions (Seavy and Howell, 2010; Ryan and others, 2018). Integrating knowledge and research findings can increase the generality, applicability, and accessibility of that information (Wyborn and others, 2018).
The introduction and establishment of nonnative species can have severe consequences for biodiversity and ecological functions (Mack and others, 2000). In the Western United States, invasive annual grasses (IAG) cause environmental problems in arid and semiarid ecosystems (D’Antonio and Vitousek, 1992; Germino and others, 2016a). For example, invasive grass species, in conjunction with humans, alter fire regimes (Fusco and others, 2019, 202167). Notably, Bromus tectorum L. (cheatgrass) has altered fire cycles in many of the sagebrush ecosystems of the Great Basin, leading to novel, annual grass-dominated communities (Germino, Belnap, and others, 2016; Pilliod and others, 2017). The wide expanse and consequences of IAGs in the Western United States have spurred a large body of literature, including review products covering Bromus spp. (brome; Germino and others, 2016b), guides for managers (Chambers, Pyke, and others, 2014; Chambers, Beck, and others, 2016), annotated bibliographies (Poor and others, 2021), and a multitude of models predicting the occurrences of these species (Tarbox and others, 2022). Although the ecosystem transformation from IAGs is a high risk in parts of the Great Basin, the different vegetation communities and climate to the east of the Rocky Mountains change the potential establishment and response of communities to IAGs (for example, Lavin and others, 2013; Taylor and others, 2014; Brummer and others, 2016).
The purpose of this report is to synthesize scientific information relevant to NEPA analyses for proposed vegetation treatments on Federal lands in rangelands in the intermontane valleys of Montana and the northern Great Plains of Montana, North Dakota, and South Dakota (hereafter referred to as “the focal region”; fig. 1). This synthesis aims to convey regional differences among the Great Basin and areas farther to the east in the Rocky Mountains and northern Great Plains where increased resistance to annual grasses has been recognized. Although increased resistance opens opportunities for different management approaches, namely habitat restoration treatments, IAGs are still a threat. Understanding which situations have a high risk of IAG establishment or expansion is key to making informed management decisions, particularly as new species, like Ventenata dubia (Leers) Coss. (ventenata), to this region expand their distribution. The synthesis also includes studies from northeastern Wyoming within the northern Great Plains that are especially relevant to the focal region. This synthesis focuses specifically on changes in risk, or possible responses, of IAG species in areas undergoing mechanical, hand-pulling, herbicide, and prescribed fire treatments. The primary annual grass species considered in this synthesis are cheatgrass and Bromus japonicus Thunb. ex Murray (Japanese brome, also known as Bromus arvensis L. [field brome]). Although the U.S. Department of Agriculture PLANTS Database (https://plants.usda.gov/home) indicates that Japanese brome is synonymous with field brome, we used Japanese brome throughout this report (Montana Natural Heritage Program, 2024a), consistent with most cited studies, and noted the studies that used the names “field brome” or “Bromus arvensis,” where cited. Information on ventenata, Taeniatherum caput-medusae (L.) Nevski (medusahead), and other species in the genus Bromus is included as available.
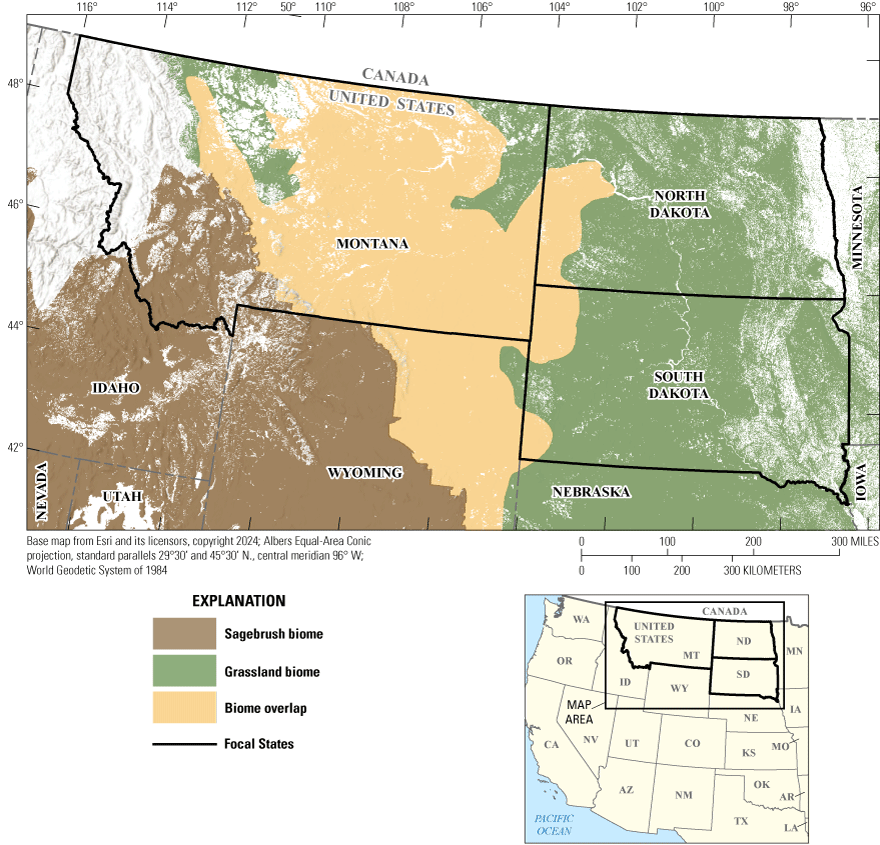
Map showing the focal region of the synthesis on invasive annual grasses within rangelands of Montana, North Dakota, and South Dakota. Rangelands as delineated by Reeves and Mitchell (2011) are color coded according to if they fall within the sagebrush biome (Jeffries and Finn, 2019), the central grassland biome (Heller, 2021), or the overlap between the two.
How to Use This Report
The content, structure, and section numbering of this report are designed to support NEPA analyses and reflect the steps of project planning and NEPA analyses (table 1). This report is meant to be a general reference and can be used, for example, as follows:
-
• Incorporated by reference in NEPA documents or to directly provide language for use in NEPA documents,
-
• Included as supplemental information to a NEPA document, or
-
• Used as a resource to gather literature and identify gaps in available science related to a management decision and context.
When incorporating this report by reference, drawing language, or citing information from this report, please use the Johnston and others (2025) suggested citation on page ii of this report.
Although the information in this report generally is organized according to steps in a NEPA process (table 1), the science synthesized in each section may inform multiple components of an analysis. In addition, the information can be applied iteratively, in a different order than the order presented in this report, or to aspects of project planning outside of the environmental effects analysis. We also provide a glossary of technical terms, and all glossary terms appear in bold at the first mention in the report.
Table 1.
Information in this report is presented to support project planning and specific steps in the National Environmental Policy Act (NEPA) analysis. Information about NEPA analyses is available in the Council on Environmental Quality NEPA regulations at 40 CFR 1500 et seq. and in department- and agency-specific policy and guidance.[Table modified from Rutherford and others (2023). IAG; invasive annual grasses]
Caveats to Use of This Report
This report is a science synthesis that focuses on research and tools within a focal region to support NEPA analyses, rather than a comprehensive literature review of IAGs across a broader geography. In addition, this report does not provide all information necessary to conduct a full environmental impact analysis or make conclusions regarding the magnitude of environmental effects. Resource planners and managers may need to supplement the information contained in this synthesis with local information. Information about specific design elements of the proposed project, local landscape conditions, and potential environmental effects from factors other than IAGs can complement the information contained in this synthesis.
Although the content of this report is focused on IAGs and vegetation treatments, this report may inform other types of actions on public lands. Many actions that occur on public lands involve machinery, traffic, and other human activities that may introduce or increase IAGs through mechanisms like those actions associated with vegetation treatments. However, because of the specificity of the science synthesized in this report, we urge caution when generalizing its content to other types of actions.
Science Synthesis—The Potential for Vegetation Treatments to Introduce and Increase Invasive Annual Grasses
1. Typical Actions and Alternatives—Common Vegetation Treatments and Associated Disturbances
Section 1 Highlights
• Vegetation treatments, used to mimic disturbance or target undesirable species, consist of fire, physical cutting and removal, or chemical approaches.
• Treatments differ in the amount of vegetation and soil disturbance resulting from the actions.
• Treatment types are selected to meet specific objectives and can be used in different combinations to increase effectiveness.
Proposed vegetation treatments in rangelands of the focal region are typically one or a combination of prescribed fire, mechanical, hand-pulling, and herbicide treatments (fig. 2A–F; DiTomaso and others, 2010; Chambers, Strand, and others, 2024). Alternative actions may include treatments applied at varying spatial scales, dosages, or in combination with other management actions such as grazing and seeding of desirable species. Historically, these treatments often were intended to increase herbaceous vegetation for livestock. Managers now increasingly use these techniques to improve wildlife habitat, reduce fuels, restore sagebrush, and increase the resistance and resilience of native communities to invasion by IAGs (Fox and others, 2009; McIver and others, 2014; Chambers, Beck, and others, 2016; Chambers, Germino, and others, 2016; Chambers, Brooks, and others, 2019; Chambers, Strand, and others, 2024). In sagebrush ecosystems, these treatments are applied to reduce or eliminate tree or shrub cover to increase herbaceous vegetation and diversify age classes of shrubs (Beck and others, 2012; Johnston and others, 2018). In grassland ecosystems, these treatments are mostly applied to generate heterogeneity, control woody vegetation, and often specifically to control IAGs, sometimes in combination with prescriptive grazing (Reinhart and others, 2020; Vermeire and others, 2021, 2023; Symstad and others, 2021).

Photographs of tools and techniques used to treat vegetation for fuels and habitat management such as A, skid steers (Photograph by Ann Boucher, Bureau of Land Management), B, herbicides (Photograph by David S. Pilliod, U.S. Geological Survey), C, prescribed fire (Photograph by Colby K. Neal, Bureau of Land Management), and D, hand-pulling (Photograph by Colby K. Neal, Bureau of Land Management). Treatments vary by type and severity of disturbances that can affect the likelihood of invasion by nonnative annual grasses. Programs like the Bureau of Land Management’s Assessment, Inventory, and Monitoring Strategy provide data on vegetation composition and cover through E, vegetation sampling (Photography by Jonathan Moor, Bureau of Land Management) that improves understanding of invasive annual grass distribution, which may increase through mechanisms that include F, unintended transport of seeds (Photograph by Jennifer Strickland, U.S. Fish and Wildlife Service).
Treatments usually involve creating staging areas and onsite visits by managers and researchers using on- and off-road vehicles on existing or new roads. Generally, seeds of IAGs can be introduced or spread by vehicles or clothing during site visits (Ansong and Pickering, 2013a, b). However, prescribed fire, mechanical, hand-pulling, and herbicide treatments differ in several ways, including type and severity of ground disturbance that can affect vulnerability of treated sites to IAGs.
Prescribed fire is commonly applied to rangelands during spring or fall (Ellsworth and Kauffman, 2017). Site preparation for prescribed fire may include construction of (1) fences for livestock exclusion to promote growth of fine fuels and (2) fire lines around burn perimeters to prevent fire spread by removing vegetation using techniques such as mowing, bulldozing, or disking that vary in severity of ground disturbance. Fire can strongly affect soil properties by increasing pyrogenic carbon, inorganic carbon, and soil pH, while decreasing soil organic carbon (Sankey and others, 2012; Santín and others, 2016; Lohse and others, 2022) and altering nutrient availability for plants (Blank and others, 2003; Rau and others, 2007). Lower fuel loads in grasslands relative to shrublands or woodlands result in lower soil heating and transformation in properties (Neary and Leonard, 2020). Severe fires can remove enough aboveground vegetation to reduce competition for colonizing grasses, whereas less severe fires can stimulate root sprout in some woody species (for example, Symphoricarpos albus (L.) S.F. Blake [common snowberry], Prunus virginiana L. [common chokecherry], Cercocarpus ledifolius Nutt. [curl-leaf mountain-mahogany], Rhus spp. L. [sumac]) but kill others (for example, Artemisia tridentata ssp. [big sagebrush], Juniperus spp. L. [juniper], Pinus spp. L. [pine]). Fire severity varies among and within applications because it is determined by many factors, including fuels, weather, moisture, and topography (Bukowski and Baker, 2013; Pausas and others, 2017).
Mechanical treatments to remove conifers or shrubs involve aerators, skid steers, excavators, masticators, rollerchoppers, chains, harrows, or tree saws with silvicultural prescriptions for selective cutting and piling or scattering wood debris (Williams and others, 2020). Piles may be burned, or sites may receive broadcast burns following mechanical treatments. Mowing is a common mechanical treatment in grasslands that can mimic some functions of fire and grazing to diversify vegetation communities and reduce woody encroachment (Vermeire and others, 2020). Mechanical treatments involve driving heavy machinery through sagebrush or grasslands and may create ground disturbances, crush vegetation, and bring seeds of undesired species to the site.
Hand-pulling treatments to remove conifers differ from mechanical treatments in that workers access treatment areas on foot from existing roads (Ross and others, 2012). Workers use chainsaws or other cutting methods to fell trees. Larger trees and branches are commonly chopped and distributed within the treatment area (lop and scatter) but also may be piled and burned. Treatments may involve complete tree removal or simply thinning, depending on the project objectives. In addition, retreatments may need to occur after a few years because this approach and similar approaches do not kill seeds and seedlings as effectively as fire (Williams and others, 2020).
Herbicides are typically broadcast by aircraft or ground-based vehicles. Tebuthiuron (Spike) is commonly applied to reduce woody vegetation like sagebrush (for example, Blumenthal and others, 2006) in favor of herbaceous cover for wildlife. Varying application rates control the level of vegetation reduction (Olson and Whitson, 2002). Alternative herbicides, like 2,4-D ([2,4-dichlorophenoxy] acetic acid), can induce larger reductions in woody and broad-leaf vegetation and affect more species (Sturges, 1993; Bureau of Land Management [BLM], 2014). Herbicides, including glyphosate, imidazolinone, and sulfonylurea, are often applied to control nonnative vegetation, especially IAGs (DiTomaso and others, 2010; Metier and others, 2019).
2. Affected Environment—Information on Possible Drivers of Invasion by Nonnative Annual Grasses and Data Sources to Characterize Their Distribution and Dynamics
2.1. A Brief Description of Rangelands of Montana, North Dakota, and South Dakota
Section 2.1 Highlights
• Sagebrush ecosystems are more common in arid parts of the focal region where deep-rooted shrubs can establish between less frequent disturbances, whereas grasslands are associated with more frequent fire because of higher and more continuous vegetation cover.
• Dominant IAGs vary across the focal region; the most common are cheatgrass, Japanese brome, ventenata, and medusahead.
Vegetation communities are variable across rangelands in the focal region. Sagebrush–grassland communities dominate the more arid parts of the region, and mixed-grass prairies dominate the more mesic sites, typically farther to the east (Montana Natural Heritage Program, 2024b, c). Sagebrush–grassland communities have a mix of sagebrush and bunchgrass species that vary across gradients of elevation and soil moisture. Grasslands of the focal region are typically mixed-grass prairies, comprising a diversity of bunchgrasses and sod forming grasses. Both major community types include a diverse range of forbs. Grasslands have higher vegetation cover and a more continuous fuel bed, which historically led to more frequent fires than sagebrush ecosystems (Guyette and others, 2015; Ashton and others, 2016).
The sagebrush biome reaches its northeastern limits in the focal region, where Artemisia tridentata ssp. wyomingensis Beetle & Young (Wyoming big sagebrush) is widespread at lower elevations with perennial bunchgrasses (for example, Pascopyrum smithii (Rydb.) Á. Löve [western wheatgrass], Achnatherum hymenoides (Roem. & Schult.) Barkworth [Indian ricegrass], Bouteloua gracilis (Kunth) Lag. ex Griffiths [blue grama], Poa secunda J. Presl [Sandberg bluegrass], and Pseudoroegneria spicata (Pursh) Á. Löve [bluebunch wheatgrass]) and forbs (for example, Phlox spp. [phlox], Arenaria spp. [sandwort], Opuntia spp. [cactus], Sphaeralcea coccinea (Nutt.) Rydb. [scarlet globemallow], Dalea purpurea Vent. [purple prairie clover], Liatris punctata Hook. [dotted blazing star], and Astragalus spp. [milkvetch]). Mesic sites on mountain slopes, mesas, and ridgetops in southwest to central Montana are often dominated by Artemisia tridentata ssp. vaseyana (Rydb.) Beetle (mountain big sagebrush). Other common shrubs include Artemisia cana Pursh (silver sagebrush), Artemisia tridentata ssp. spiciformis (Osterh.; subalpine big sagebrush), Artemisia tripartita Rydb. (three-tip sagebrush), Artemisia arbuscula Nutt. (low sagebrush), Sarcobatus vermiculatus (Hook.) Torr. (black greasewood), Atriplex spp. [saltbush], Ericameria nauseosa (Pall. ex Pursh) G.L. Nesom & Baird (rubber rabbitbrush), and Purshia tridentata (Pursh) DC. (antelope bitterbrush). Mesic sites support diverse grass (for example, Festuca campestris Rydb. [rough fescue], Festuca idahoensis Elmer [Idaho fescue], Elymus elymoides (Raf.) Swezey [squirreltail], and Achnatherum spp. [needlegrass]) and forb (for example, Balsamorhiza sagittata (Pursh) Nutt. [arrowleaf balsamroot] Castilleja spp. [Indian paintbrush], Potentilla spp. [cinquefoil], Erigeron spp. [fleabane], phlox, and milkvetch) communities. Herbaceous to shrub cover ratios generally increase with moisture, elevation, and longitude (Montana Natural Heritage Program, 2024b, c). Cheatgrass is most common in warm, dry regions of southwestern Montana, whereas Japanese brome, also known as field brome (Montana Natural Heritage Program, 2024a), is more common in cool, wet environments in eastern parts of the focal region. Conifer encroachment is especially common in southwestern Montana and has led to the application of vegetation treatments to recover sagebrush communities (Williams and others, 2020).
Mixed-grass prairie covers much of the eastern parts of the focal region, where grass cover is usually greater than 50 percent and common species include Festuca spp. (fescue), western wheatgrass, squirreltail, Hesperostipa comata (Trin. & Rupr.) Barkworth (needle-and-thread), Hesperostipa curtiseta (Hitchc.) Barkworth (shortbristle needle and thread), Elymus lanceolatus ssp. lanceolatus (Scribn. & J.G. Sm.) Gould (thickspike wheatgrass), and Nassella viridula (Trin.) Barkworth (green needlegrass). Japanese brome is more common than cheatgrass, and nonnative perennial grasses (for example, Poa pratensis L. [Kentucky bluegrass]) are also problematic invaders. Fire has been an important disturbance in this region with return intervals of approximately 1–35 years in grasslands (Zouhar, 2021) and 20–80 years in sagebrush (Innes and Zouhar, 2018). Grazing by wildlife also disturbs and modifies vegetation communities. Grazing by livestock is common on public lands (Derner and others, 2023), which are mostly administered by the BLM or by the U.S. Department of Agriculture Forest Service as national grasslands. Remnants of native prairie occur on these lands and other lands administered by the U.S. Fish and Wildlife Service and National Park Service.
2.2. Factors Determining Vulnerability of Sites to Invasion by Nonnative Annual Grasses in the Western United States
Section 2.2 Highlights
• Factors affecting invasion resistance and resilience globally include propagule pressure, native species richness, and land use history.
• Precipitation seasonality and changes to disturbance regimes affect the likelihood of nonnative annual grass invasions.
• Annual grasses tend to be the dominant invader of sagebrush ecosystems, whereas grasslands face additional threats from nonnative perennial grasses and forbs and woody species.
2.2.1. Climate, Competition, and Propagules
Establishment of nonnative species can lead to severe impairment and changes to ecosystem processes and functions (D’Antonio and Vitousek, 1992; Davis and others, 2000; Germino, Belnap, and others, 2016). The processes, drivers, and factors governing the establishment and expansion of IAG species in the Western United States are the subject of a large body of literature. Interactions among climate, disturbance, and human factors affect the resistance of habitats to invasion and determine the specific species that can be successful invaders (Brooks and others, 2016; Chambers, Germino, and others, 2016). Annual and perennial grass invasions are particularly distinct in arid and semiarid regions; annual species occur in the drier regions, and perennial species are in more mesic regions (D’Antonio and Vitousek, 1992). The introduction, expansion, and effects of IAG species have been covered in several reviews, annotated bibliographies, and reports (for example, Knapp, 1996; Mack, 2011; Germino and others, 2016b; Poor and others, 2021; Meineke and others, 2024). Gaskin and others (2021) provide a general review of invasive plant issues in the Great Plains, and additional literature is available on responses of perennial nonnative grasses to management and treatments (for example, DeKeyser and others, 2013; Kobiela and others, 2017; Bennett and others, 2019; Dornbusch and others, 2020).
Key determinants to the susceptibility of habitats to the invasion of Bromus species are climate and the native plant community. For example, cheatgrass is a dominant invader in the Great Basin, but more limited in northern Great Plains communities. This dominance is not entirely because poor suitability of climate or soil conditions, but also because cheatgrass is competitively excluded by native perennial grass species (Brooks and others, 2016). Sites more susceptible to cheatgrass tend to have lower summer precipitation (winter or early spring dominated moisture regimes) and warmer summers and winters (Taylor and others, 2014; Brummer and others, 2016). In addition, the seasonality of precipitation has consequences for water availability and hence herbaceous cover and ecosystem resilience (fig. 3A–E). Changes to the disturbance regime can allow nonnatives to establish and effectively outcompete native species (Germino, Belnap, and others, 2016; Brooks and others, 2016). Although widely implicated in the spread of IAGs, fire is not required for invasion (Smith and others, 2023). Once established, however, IAGs can initiate feedback in grass–fire cycles that facilitate their dominance.
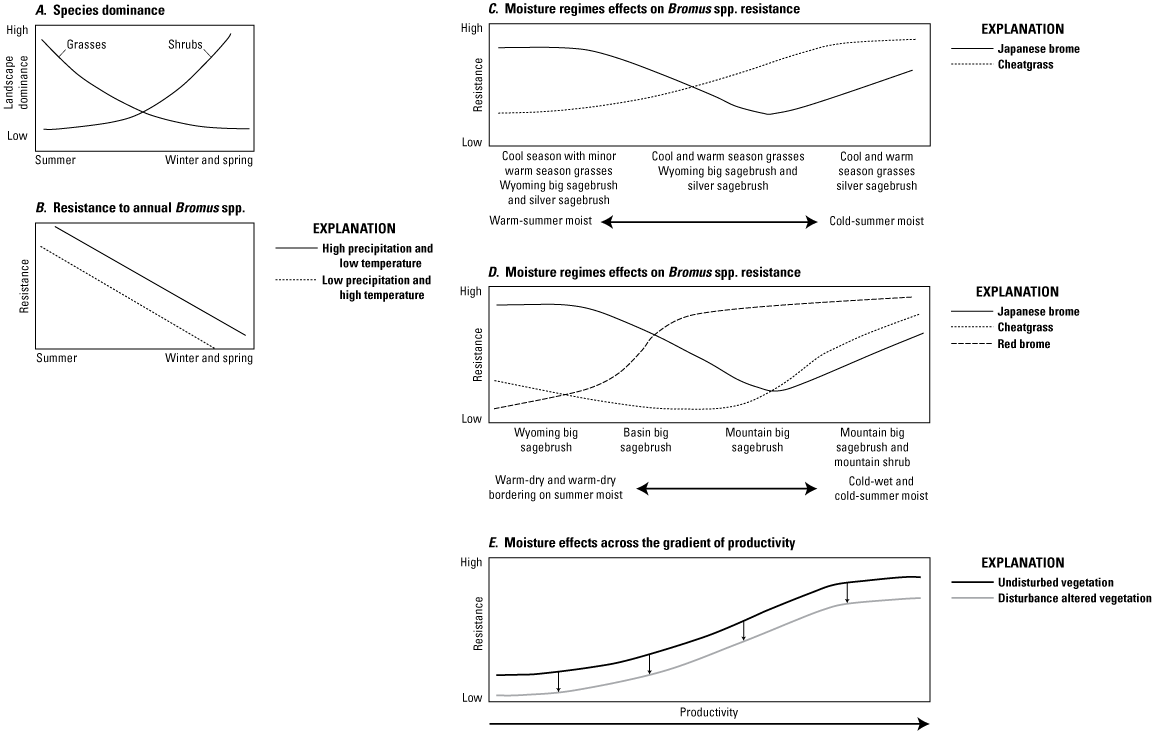
Graphs showing the effects of precipitation seasonality effects on A, shrub–grass ratios and B, resistance to annual Bromus spp. (bromes); the effects of climate on resistance to C, Bromus japonicus Thunb. ex Murray (Japanese brome; also known as Bromus arvensis L. [field brome]) and Bromus tectorum L. (cheatgrass) and D, Japanese brome, cheatgrass, and Bromus rubens (red brome) across sagebrush communities; and E, the effects of disturbance on these relationships across the gradient of productivity. Modified from Chambers, Beck, and others (2016).
Propagule pressure, native species richness, and land use history can affect the likelihood that a system will be invaded by nonnative plant species (Lonsdale, 1999; Davis and others, 2000; Fridley and others, 2007; De Stefano and others, 2024). In many ecosystems, plant communities become more invadable when there are more unused resources including light, water, and nutrients. These pulse events can be because of lags between when resources become available and when plants use them (such as early season soil water recharge not being used by native plants until later in the growing season) or from disturbance events releasing nutrients and causing plant mortality (D’Antonio and Vitousek, 1992; Davis and others, 2000). When these resource pulses occur, invasion factors such as propagule pressure and landscape condition determine the chances of establishment of nonnative species (Davis and others, 2000). Locations with high propagule pressure and disturbed or heterogenous landscape conditions may be most likely to match invasive species with suitable resources. When nonnative grasses invade, there is a risk of changes to successional processes (for example, grass–fire cycles) and increased variability in productivity (Germino, Belnap, and others, 2016; Pilliod and others, 2017).
2.2.2. Climate Suitability for Invasive Annual Grasses in Montana, North Dakota, and South Dakota
As climate suitability for cheatgrass decreases (in other words, areas with more spring–summer precipitation and colder winter temperature), resistance to IAGs and resilience to disturbances like fire increase (Bradley, 2009; Bradley and others, 2016). Rangelands in the focal region typically have climatic conditions that are less suitable for cheatgrass. These areas have evolved with disturbances (for example, drought, fire, grazing) and tend to recover quickly (Brooks and others, 2016). However, changes to climate (increased aridity or temperatures) and disturbances outside of historical regimes (frequency, timing, and so forth) can reduce the competitive advantage of native plant species. As of 2024, areas within these rangelands that have cheatgrass and Japanese brome tend to be more arid (south facing and lower elevation) or exposed to surface disturbance (Brooks and others, 2016).
Less is known about suitability of rangelands in the focal region for ventenata and medusahead (Adhikari and others, 2023). Ventenata can occupy distinct areas and may invade higher elevations more readily than cheatgrass, despite having similar responses to seasonal precipitation (Applestein and Germino, 2022b; Prevéy and others, 2024). Ventenata often co-occurs with medusahead in Idaho and Oregon, occupying areas that have distinct environmental conditions from areas with cheatgrass (Jones, Norton, and Prather, 2018; Tortorelli and others, 2020). Adhikari and others (2023) found that suitability for ventenata in Montana may increase with climate change.
Although IAGs tend to be a larger threat in sagebrush–grassland communities, the grasslands of the northwestern Great Plains also face additional threats. Invasive plant species include invasive annual and perennial grass species (for example, Kentucky bluegrass and Bromus inermis Leyss. [smooth brome]) and woody species (Symstad and Leis, 2017; Gaskin and others, 2021). Therefore, managers may also need to consider broader encroachment and more invasive species in restoration and treatment plans (Symstad and Leis, 2017). Much research on invasive plant species responses to vegetation treatment in grasslands of the northern Great Plains is focused on invasions by perennial and woody species, rather than IAGs. Nevertheless, in sampling from 1998 to 2015 across seven National Park Service units in the northern Great Plains, annual brome cover was found to be highly variable across years but did increase throughout the study period (Ashton and others, 2016).
2.3. Available Datasets of Invasive Annual Grasses Presence and Abundance
Section 2.3 Highlights
• Both in situ and modeled (for example, remote sensing derived) datasets exist to provide contextual and possibly site-specific data to supplement local unit records.
• Guidance is available to help understand which invasive annual grass model may be best suited for a specific question.
Data for IAGs are available for much of the focal region because invasive species are included in vegetation sampling protocols (for example, Herrick and others, 2017) and multiple approaches have been developed for predicting IAG presence based on remotely sensed imagery (Tarbox and others, 2022). Therefore, locally collected, in-house data (for example, historical surveys and Daubenmire plots) often can be supplemented with data from remotely sensed products to provide a broader understanding of current presence and abundance on the landscape for NEPA analyses. In addition, where site-specific data may be limited, predictive models of IAGs can provide information about potential presence in and around project locations.
2.3.1. Sources of Field-Collected Data on Invasive Annual Grasses
Vegetation sampling protocols that characterize species composition as well as specific inventories of IAGs provide useful data about IAG presence. For example, invasive plants are included in protocols such as “Interpreting Indicators of Rangeland Health” (Pellant and others, 2020), terrestrial vegetation monitoring surveys as part of the BLM Assessment, Inventory, and Monitoring (AIM)Strategy program (Toevs and others, 2011; Kachergis and others, 2022), and in the U.S. Department of Agriculture Natural Resources Conservation Service National Rangeland Inventory (Herrick and others, 2017). Additional location data may be available from the National Park Service (Symstad and others, 2012), U.S. Fish and Wildlife Service (Gannon and others, 2013), State Natural Heritage Programs (for example, through the Montana Natural Heritage Program Map Viewer [https://mtnhp.org/mapviewer/]), State wildlife agencies, and online mapping efforts (for example, Early Detection and Distribution Mapping System [https://www.eddmaps.org/] and Global Biodiversity Information Facility [https://www.gbif.org/]).
In situ plot data can provide site-specific data to complement vegetation maps or develop new products to understand IAG distribution in project areas. Species-specific data can be used to derive occurrence or cover estimates, although care should be taken to understand the sampling schema, purpose, and approach when moving from point data to estimates for a project area; metrics from existing data may not represent vegetation characteristics for specific questions and areas of interest. Because data collection in monitoring programs often covers large geographic areas, the availability of few surveys within local areas and lack of site-specific information in some sources (for example, the National Rangeland Inventory) can limit the utility of these datasets as sole sources for analyses of specific sites.
2.3.2. Vegetation Maps from Remote Sensing
The availability of a large amount of in situ data for IAG species, in combination with key spectral, habitat, and phenological characteristics of many invasive species (Bradley, 2014) has led to multiple remote sensing-based products that provide information on the presence, abundance, and cover of IAGs (table 2; fig. 4A–D). Available data include those data mapping fractional cover (for example, tools such as the U.S. Department of Agriculture Agricultural Research Service Rangeland Analysis Platform [RAP; Jones and others, 2018], Rangeland Condition Monitoring Assessment and Projection [RCMAP; Rigge and others, 2021], and Landscape Cover Analysis and Reporting Tools [LandCART; Zhou and others, 2020]; IAGs are mapped as a group in these products), species-specific estimations of cover or presence (Montana Natural Heritage Program 2021a, b, c), risk models, and models estimating future risk based on climate change. Within the models for spatial coverage of IAG species are those models estimating cover annually back in time (Jones and others, 2018; Rigge and others, 2021), cover early in the growing season to help focus treatments (Dahal and others, 2022), or estimating future changes in distribution (Bradley, 2009). Tarbox and others (2022) reviewed 23 vegetation maps that describe IAG distribution across key western regions, the entire Western United States, or the Nation. A web-based tool, INHABIT (https://gis.usgs.gov/inhabit/), also provides species distribution models with downloadable maps of IAG distribution across the continental United States (Engelstad and others, 2022). This tool also shows the modeled relationships of each species with environmental factors, providing information for assessing IAG distribution in project areas.
2.3.3. Maps of Resistance and Resilience to Invasion for Decision Making
Another type of IAG model with broad spatial coverage includes those models examining areas at risk for invasion by annual grasses. Primarily these models are based on the resistance and resilience of a geographic area to fire and IAGs. Mapping is then based on factors such as climate, soils, and other environmental gradients (Chambers, Maestas, and others, 2017; Chambers, Brown, and others, 2023), which may be used to prioritize areas for conservation and restoration. Combined with maps of focal ecosystems (for example, sagebrush), these resistance and resilience maps can enhance identification of areas that are most likely to function as core areas for native species through time. Models that predict resistance and resilience with climate-based variables may be forecasted with alternative climate scenarios to assess potential changes in future IAG distributions. Application of these concepts to management is included in several references (for example, Chambers, Pyke, and others, 2014; Chambers, Brooks, and others, 2019; Chambers, Brown, and others, 2023). Some resources focus on areas to the east of the Rocky Mountains, that is, the eastern part of the Centrocercus spp. (sage-grouse) range (Chambers, Beck, and others, 2016). Maps in these studies partially cover Montana and the western parts of North Dakota and South Dakota.
2.3.4. How to Decide What Invasive Annual Grass Data Products and Sources to Use to Inform Your Decision
The multitude of potential models that use various approaches for different purposes can make it difficult to choose an appropriate data source. Allred and others (2022) recommend four principles to guide the selection of a mapping product: “1) use maps in a decision-making framework, 2) use maps to better understand and embrace landscape variability, 3) keep error in perspective, and 4) think critically about contradictions” (p. 79). Additional references are available to inform selection of vegetation maps for IAGs, including a guide for managers (Van Schmidt and others, 2022), a compendium that summarizes 23 reviewed products (Saher and others, 2022), a database with summaries of these products (Shyvers and others, 2022), and a publication describing the selection and review methodology for these products (Tarbox and others, 2022). These references summarize critical characteristics of data sources, including map extent, content, and spatiotemporal resolution for users to determine data suitability for an analysis.
Table 2.
Data products derived from remote sensing and spatial data available at the time of publication that provide information on the presence or cover of invasive annual grasses for Montana, North Dakota, and South Dakota.[Refer to Saher and others (2022) for more data products and description. spp., several species of; USGS, U.S. Geological Survey; %, percent; IAG, invasive annual grass]
| Name | Location | Description | Citation |
|---|---|---|---|
| Rangeland Analysis Platform (RAP) | https://rangelands.app/ | Landsat (satellite) derived estimates of fractional cover of vegetation groups (including annual herbaceous), covering the contiguous United States. | Jones and others, 2018 |
| Rangeland Condition Monitoring Assessment and Projection (RCMAP) | https://www.mrlc.gov/rangeland-viewer/ | Landsat (satellite) derived estimates of fractional cover of vegetation groups (including annual herbaceous), covering the Artemisia spp. (sagebrush) biome. | Rigge and others, 2021 |
| Fractional Estimates of Multiple Exotic Annual Grass (EAG) Species in the Sagebrush Biome, USA, 2016–2024 (ver. 5.0, July 2025) | https://doi.org/10.5066/P9GC5JVG | Historical (2016–23) estimates of fractional cover for nonnative annual grasses in rangelands, covering Western States, including Montana and the western halves of North Dakota and South Dakota. | Dahal and others, 2022 |
| New Indicators of Ecological Resilience and Invasion Resistance to Support Prioritization and Management in the Sagebrush Biome, United States | https://doi.org/10.5061/dryad.h18931zpb | Indices of relative resistance and resilience based on climate and soil water availability within the sagebrush biome of Western States, including Montana, North Dakota, and South Dakota. | Chambers and others, 2023 |
| Historic and Future Trends in Exotic Annual Grass (%) Cover in the Western United States (from 1985 to 2019 and 2025 to 2040) | https://doi.org/10.5066/P9Z85VET | Modeled historical conditions and future predictions of IAG cover and trend across Western States, including Montana and the western halves of North Dakota and South Dakota. | Pastick and others, 2021a |
| Annual Herbaceous Cover across Rangelands of the Sagebrush Biome | https://doi.org/10.5066/P9VL3LD5 | Combination of RAP, RCMAP, and USGS Harmonized Landsat and Sentinel (Pastick and others, 2020) data products to create weighted mean synthesis across Western States, including Montana and the western halves of North Dakota and South Dakota. | Maestas and others, 2020 |
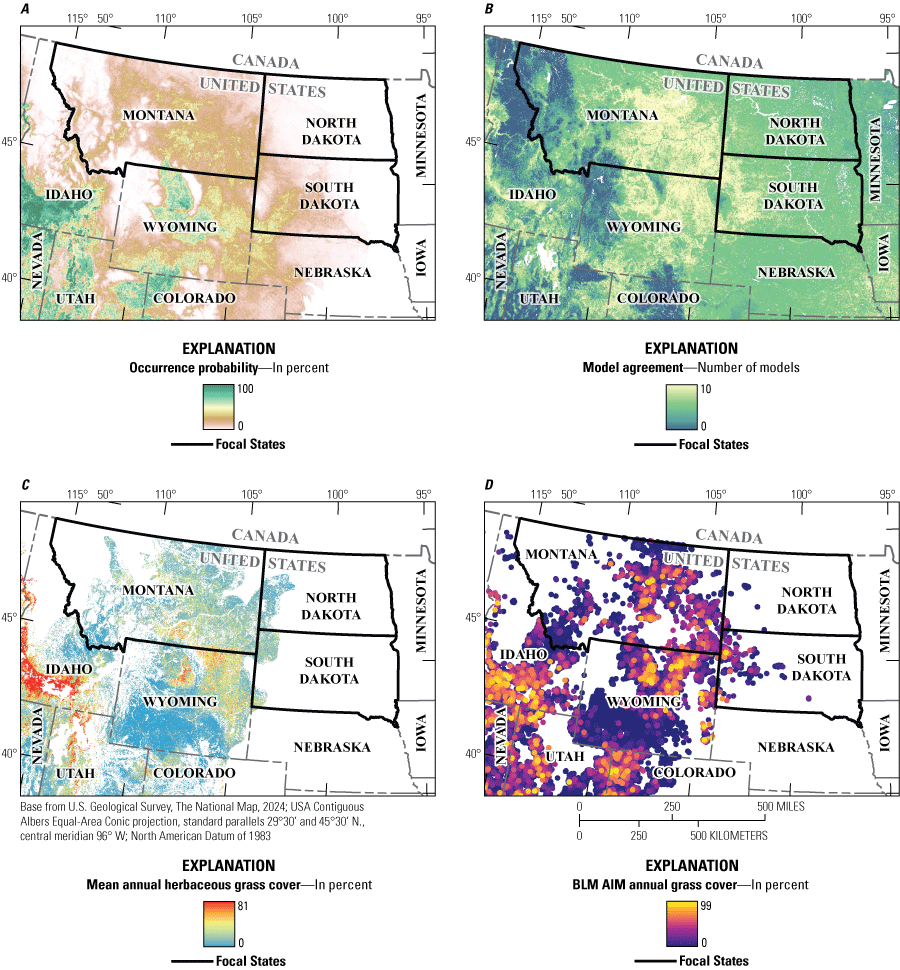
Maps showing the distribution of invasive annual grasses in Montana, North Dakota, and South Dakota based on A, occurrence probability (in percent) for Bromus tectorum L. (cheatgrass), B, occurrence agreement across 10 habitat suitability models for Bromus japonicus Thunb. ex Murray (Japanese brome), C, mean cover (in percent) of annual herbaceous vegetation across satellite-based models, and D, percent cover of annual grasses at Bureau of Land Management (BLM) Assessment, Inventory, and Monitoring (AIM) plots. Data from McMahon and others (2021), Williams and others, (2023), Maestas and others, (2020), and BLM (2021), respectively.
Map utility varies with the types of questions, scales of analysis, accuracy, and magnitude of effects under investigation, but understanding appropriate scales of analysis for fractional cover maps remains an active area of research (Applestein and Germino, 2021, 2022a, 2023). Some map developers have cautioned that fractional cover maps are inappropriate for pixel-scale applications (Savage and Slyder, 2022). Although many of these maps are available at 30-meter resolution, errors in pixel-level cover estimates may lead to misleading results when used to evaluate vegetation occurrence or change within small treatments that encompass few pixels. Errors reported in validation studies for these maps reflect mean values from ecosystem-wide assessments (Rigge and others, 2019, 2020, 2021; Allred and others, 2021; Zhang and others, 2019; Savage and Slyder, 2022); however, error can vary substantially across and within regions by unknown amounts and for different reasons. Accordingly, applications of fractional cover maps, such as estimating propagule pressure and risk of increasing IAGs from vegetation treatments, may be accomplished with the greatest confidence when findings are corroborated from multiple data sources, including ground data.
3. Methods and Considerations for Evaluating Potential Increases of Invasive Annual Grasses from Vegetation Treatments
Section 3 Highlights
• Risk factors, such as current IAG presence, climate, site location, and treatment approach can help managers decide on the appropriate level of analysis.
• Formal modeling approaches at the local level may allow for consideration of additional factors not considered in broader extent models.
• Cumulative effects may include the presence and cover of IAGs in the surrounding landscape as well as additional restoration and vegetation treatments.
Depending on available data, resources, and expertise, NEPA analyses of potential IAG increases from vegetation treatments range from rapid, qualitative assessments to rigorous modeling of invasion probabilities in the project area under alternative management and weather or climate scenarios. Qualitative assessments may be most appropriate and practical when resources for analyses are limited and when the potential for increasing IAGs is clearly high or low. This type of assessment may consist of overlaying the project map with maps of IAG distribution (for example, fractional cover and probability of occurrence), resistance and resilience, and other important factors like aspect. Factors known to affect IAG occurrence may be found in existing models of species distributions. For example, INHABIT includes plots for the modeled relationship of each species and each predictor of its distribution. Where available, data from field surveys within the project area can validate patterns of IAG distribution in vegetation maps.
Additional insights on potential increases in IAGs from vegetation treatments may be found by formally analyzing IAG distribution and important factors within the project area. Spatially explicit analysis of IAG distributions for a project area requires maps of factors that affect IAG occurrence. These factors often include topographic (for example, slope, aspect, and elevation), environmental (for example, soil type, moisture potential, and proximal invasions), or anthropogenic (for example, distance to road, oil and gas development, and previous treatment) features. Using ground survey data or maps of IAG presence (for example, fractional cover derived from satellite imagery), analysts can model and map the probability of IAG occurrence with species distribution models (for example, maximum entropy modeling [MaxEnt]; Jarnevich and Young, 2015), regression (for example, conditional regression, hurdle models; Taylor and others, 2014), or machine learning (for example, random forest). Incorporating contagion, migration, or other models that explicitly account for propagule pressure and the spatial context of areas of interest can improve understanding of resistance to IAGs (Thomas and Moloney, 2015; Maestas and others, 2022). Modeled relationships can be projected across the proposed project area to depict the probability of occurrence (that is, risk of invasion) at any given location according to the variables evaluated by the model. Distribution of IAGs can be forecasted with weather or climate scenarios using modeled relationships of IAG occurrence with temperature and precipitation to describe potential distributions in the future (Homer and others, 2015; Palmquist and others, 2021). Modeling distributions within project areas may permit the assessment of local factors that were not considered in existing distribution maps for IAGs. For example, grazing timing and intensity may be an important factor in IAG distribution that is well documented within a project area but not represented in existing models because information on grazing activities is not widely available and often documented at scales too coarse to explain IAG distribution.
Analyses of IAG distributions may include factors controlled by managers (for example, grazing, vegetation treatments) or other factors that can change through time (for example, climate) to develop a range of potential outcomes for IAG distribution. State and transition models can support such evaluations by describing changes in plant and soil communities and evaluating potential outcomes amid different environmental conditions (Chambers, Pyke, and others, 2014). Regions previously thought to have a low risk of invasion by annual grasses may become threatened as climatic patterns shift and produce abnormal conditions (for example, hotter, more frequent fires) that may favor future invasions (Palmquist and others, 2021; Smith and others, 2021156). For example, Bradley and others (2016) evaluated potential range shifts in Bromus rubens L. (red brome) and cheatgrass with future climate scenarios and found that cheatgrass may expand into southwest and southcentral Montana, whereas red brome may expand into the southern Great Basin and Colorado Plateau as global temperatures rise. Approaches that account for alternative management and climate scenarios to assess current and future IAG distributions can require more time and labor than are available for some NEPA analyses. In addition, the necessary datasets (for example, local vegetation surveys and grazing intensity) may not be available for some project areas.
Cumulative effects may be assessed with time series of vegetation cover and other landscape features that change through time. Vegetation maps that provide annual grass cover estimates annually based on archives of satellite imagery (for example, RCMAP and RAP) can provide baseline information on the distribution of annual grasses before treatments and other disturbances. Using analytical approaches for repeated measures (for example, mixed models), analysts can compare IAG distribution before and after disturbances to assess timelags in the spread of IAGs and identify interactions of multiple disturbances that reflect cumulative effects after accounting for important factors for IAG distribution (for example, topography, weather, and soil type).
4. Environmental Consequences—Information on the Potential Spread of Invasive Annual Grasses from Vegetation Treatments
4.1. Sagebrush Ecosystems
Section 4.1 Highlights
• Weather and pre-existing IAGs affect the likelihood of vegetation treatments leading to an increase in abundance and distribution of IAGs.
• Prescribed fire often reduces or does not increase IAGs.
• Mechanical disturbances to the soil may increase IAGs.
• Herbicides to reduce sagebrush have not increased the proportion of IAGs in the plant community.
4.1.1. What Environmental Factors Affect Invasive Annual Grass Distribution in Sagebrush Ecosystems?
Temperature and precipitation are strong factors in determining IAG responses to vegetation treatments in the northeastern region of the sagebrush ecosystem (Haferkamp and others, 1993; Taylor and others, 2014; Larson, Lehnhoff, and Rew, 2017; Larson and others, 201795). Across the sagebrush ecosystem, mean annual temperature and summer precipitation were the best predictors of cheatgrass responses to fire more than other climate variables, where sites with high temperatures and low precipitation were most likely to show increases in cheatgrass after fire (Taylor and others, 2014). Greater spring and summer precipitation likely provide a competitive edge to native, perennial grasses more than winter annual grasses like cheatgrass (Bradley, 2009; Taylor and others, 2014). At fire-treated areas in eastern Idaho, cover by native grasses was negatively correlated with cheatgrass presence, which is consistent with studies in other regions that suggest competition and species composition affect IAG distribution (Germino, Belnap, and others, 2016). Larson, Lehnhoff, and Rew (2017) concluded that experimental warming and drying were detrimental to cheatgrass and much more effective than spring fire in Montana. Their results suggested that low levels of winter precipitation limited the spread of cheatgrass and that only changes in seasonal precipitation would likely increase invasion risks. Increasing temperature and decreasing summer or winter precipitation from climate change may lead to greater increases in IAGs, particularly in southern and southwest Montana compared to other parts of the focal region (Bradley, 2009; Taylor and others, 2014).
4.1.2. How Does Prescribed Fire Affect Invasive Annual Grass Distribution in Sagebrush Ecosystems?
Most studies (table 3) on vegetation treatments in sagebrush and IAGs in or near the focal region have examined effects of fire, where it often has resulted in either a reduction or no change to IAG cover (Taylor and others, 2014; Porensky and Blumenthal, 2016; Larson, Lehnhoff, and Rew, 2017; Porensky and others, 2018; Seipel and others, 2018; Pastick and others, 2021b). Fire directly affects IAGs through mortality of plants and seeds, and indirectly through the release of nutrients and by increasing available space if existing native vegetation is killed and cannot resprout. These potential effects can be mediated by local conditions such as aspect and weather. For example, Larson, Lehnhoff, and Rew (2017) experimentally applied spring fire with warming and drying treatments to sagebrush steppe sites for 3 years in southwest Montana to evaluate invasions by cheatgrass and potential effects of climate change. Warming and drying treatments reduced cover, biomass, and seed production of cheatgrass and the dominant native species, bluebunch wheatgrass. Fire treatments reduced cover by native grass species but did not change biomass or cover of cheatgrass. However, seed production of cheatgrass increased following fire treatments. Results reflected only short-term vegetation responses because posttreatment monitoring was limited to the growing season for the year of treatment.
Table 3.
Brief results of studies on increases of invasive annual grasses (IAG) from vegetation treatments and wildfires in sagebrush or grassland ecosystems in or near Montana, North Dakota, and South Dakota.| Region | Treatment | Change in IAG | Citation |
|---|---|---|---|
| Western United States | Prescribed fire | None observed in Montana; predicted increased risk to postfire invasion with climate change in Montana and Wyoming | Taylor and others, 2014 |
| Southwest Montana | Prescribed fire | No change in biomass or cover; increased seed production | Larson, Lehnhoff, and Rew, 2017 |
| Southwest Montana | Prescribed fire | No change in burned areas; increased in bulldozed fire breaks | Seipel and others, 2018 |
| East Montana | Mechanical and herbicide | Increased biomass; no change in percent composition of the plant community | Haferkamp and others, 1993 |
| North Wyoming | Herbicide | None | Johnson and others, 1996 |
| Southeast Montana and south Wyoming | Prescribed fire | None or decreased | 1Wade and others, 2024 |
| Northeast Wyoming | Wildfire | Decreased | Porensky and Blumenthal, 2016 |
| Northeast Wyoming | Wildfire | None | Porensky and others, 2018 |
| Northeast Wyoming and southeast Montana | Grazing and precipitation | Decreased with reduced precipitation; slightly increased with heavy grazing and reduced precipitation | Frost and others, 2023 |
| Southwest Montana and north Montana | Wildfire | None in burned areas; increased in bulldozed fire breaks | Tulganyam and Carr, 2022 |
| Northwest Great Plains | Wildfire | None for annual grass functional group | Donovan and others, 2020 |
| East Wyoming, west South Dakota, and west Nebraska | Fire (prescribed or wild) and grazing | None from grazing; slight increase with time since fire | Ashton and others, 2016 |
| Southwest South Dakota | Prescribed fire | Decreased | Whisenant and Uresk, 1990 |
| Southeast Montana | Prescribed fire | Decreased | Vermeire and Russell, 2018 |
| Southeast Montana | Prescribed fire | Decreased | Vermeire and others, 2011 |
| Southeast Montana | Prescribed fire | Decreased | Vermeire and others, 2014 |
| Southeast Montana | Prescribed fire | Decreased | Vermeire and Rinella, 2017 |
| Southeast Montana | Mechanical | Increased | Vermeire and others, 2020 |
| Southeast Montana | Mechanical and fungicide | Decreased from mowing; no change from fungicide | Reinhart and others, 2020 |
| Southeast Montana | Prescribed fire and herbicide | Decreased from fire; herbicide decreased germination but not biomass | Vermeire and others, 2021 |
| Southeast Montana | Grazing | Timing and intensity associated with decrease or increase of two IAG species | Vermeire and others, 2023 |
| Southwest South Dakota and west Nebraska | Prescribed fire and herbicide | Decreased | Symstad and others, 2021 |
Taylor and others (2014) related responses of cheatgrass to fire from 18 studies distributed across several Western States that represented a range of conditions from warm and dry to cool and wet. Responses of cheatgrass to fire were positive at warm, dry sites, whereas fire did not show any response at cool, wet sites, including those sites in the northeastern region of the sagebrush ecosystem—the part of their study directly related to our focal region. The best model indicated that the probability of positive responses of cheatgrass after fire increased with mean annual temperature and lower summer precipitation. Under current climate conditions, they predicted a low risk of invasion by cheatgrass after fire in most of the focal region (Taylor and others, 2014). Under future climate scenarios, risk generally remained low but increased most substantially in southern Montana and western Montana. In addition, they found no increase in cheatgrass across a chronosequence of fires that varied 1–15 years in the time since fire in the upper Snake River Plain of southeastern Idaho, which had intermediate climate conditions.
Similarly, Seipel and others (2018) did not find differences in cheatgrass cover between burned and unburned areas through 3 years of postburn monitoring in southwestern Montana. However, bulldozed firebreaks showed different vegetation communities with higher amounts of cheatgrass relative to burned and unburned areas. High cover of IAGs also occurred in bulldozed firebreaks in grasslands of Montana (Tulganyam and Carr, 2022), which suggests that mechanical disturbance to soil is a distinct factor that can increase IAGs in this focal region.
Burn frequency and severity can be important factors that determine IAG response to fire. Burn severity has been strongly correlated with cheatgrass invasion in the Great Basin (Davies and others, 2009), and simulated responses of IAGs to changing environmental conditions across Wyoming and Utah indicate strongly positive effects of pre-existing IAGs and repeat fires on invasions (Pastick and others, 2021b). However, Taylor and others (2014) did not find differences in cheatgrass invasion at sites with repeat fires and only slight increases at more severely burned sites in southeast Idaho, where increases in perennial grasses after fire may have suppressed cheatgrass. These factors remain poorly understood in much of the focal region and are likely less important than climate for the broader distribution of IAGs in this region.
Although wildfires can produce unique vegetation responses compared to prescribed burns because of differences in burn season, severity, and extent, studies in northeastern Wyoming corroborate findings that fire does not increase IAGs in this region (Porensky and Blumenthal, 2016; Porensky and others, 2018). Cover by cheatgrass and Japanese brome was similar (Japanese brome was described as Bromus arvensis in Porensky and others [2018]) or lower (Japanese brome was described as Bromus arvensis in Porensky and Blumenthal [2016]) in burned compared to unburned areas sampled across several wildfires; measurements occurred 1–26 years postburn. Determinants of higher brome occurrence included sandy and rocky soil types, higher slopes, southeast aspect, higher plant cover, and lower shrub to perennial ratios (Porensky and Blumenthal, 2016).
4.1.3. How Do Herbicide and Mechanical Treatments Affect Invasive Annual Grass Distribution in Sagebrush Ecosystems?
Few studies have examined IAG responses to mechanical or herbicide treatments in the northeastern region of the sagebrush biome. Haferkamp and others (1993) attributed increases in herbaceous vegetation, including IAGs, to soil disturbances from mechanical treatments but did not find concomitant increases in percent composition of IAGs in the plant community for seven treatment types throughout 8 years of monitoring in eastern Montana, where cheatgrass and Japanese brome were common prior to treatments. Treatments included several combinations of mechanical, herbicide, seeding, and grazing, all of which included soil disturbance, except at control sites. Brush, including Wyoming big sagebrush, was completely removed at two sites by chopping or applying 2,4-D, whereas other sites had mechanical treatments of tilling or contour furrows. Temperature and precipitation explained more variation in herbaceous cover than treatment effects. In northcentral Wyoming, cheatgrass was present but not an important part of community composition 8–14 years after tebuthiuron treatments to thin Wyoming big sagebrush (Johnson and others, 1996). Sagebrush thinning increased with increasing concentrations of tebuthiuron, which resulted in some increases in herbaceous cover that were dominated by native species.
4.1.4. Additional Information to Enhance Understanding of Vegetation Treatments in Sagebrush Ecosystems
Research on climate, IAGs, native vegetation, and vegetation treatments can support better understanding of the interaction of these variables in sagebrush ecosystems under a changing climate. Often, studies are focused on the treatment of existing locations of IAGs directly, rather than as an unintended consequence of restoration actions. Focused monitoring and research following prescribed fire, conifer mastication and hand-pulling, and other treatments would help corroborate the inferences leading to the higher resistance observed in the shrublands in the focal region. In addition, whereas increases of IAGs from vegetation treatments has been limited in the focal region, climatic factors and competition with native species have been attributed as primary factors in the resistance and resilience to IAGs in this region (Bradley and others, 2016; Brooks and others, 2016). Forecasted increases in temperature and changes in the amount and seasonality of precipitation are expected to alter vegetation dynamics in ways that could favor IAGs (Palmquist and others, 2021), but confirmatory research is limited. Evaluations of the changes in IAG distribution across landscapes through many years would improve understanding of environmental relationships of IAGs and the likelihood of changing dynamics in invasion risk. Such studies could be facilitated by a time series of fractional cover or IAG distribution provided by archives of satellite imagery and compilation of ground surveys with broad geographic and temporal distributions (for example, surveys conducted through BLM’s AIM Strategy program).
4.2. Grassland Ecosystems
Section 4.2 Highlights
• Prescribed fire often reduces IAGs in grassland ecosystems, but effects depend on the abundance of native species.
• Mowing can increase or decrease IAGs in grassland ecosystems.
• Herbicides can increase control of IAGs following fire.
• Grassland site conditions, such as the southeast aspect, sandier or rockier sites, or lower native species cover or richness, may mediate invasions by IAGs.
4.2.1. What Environmental Factors Affect Invasive Annual Grass Distribution in Grassland Ecosystems?
Studies in grassland ecosystems show the lack of disturbance (for example, longer return intervals or removal of grazing) as a factor leading to an increase in invasive grasses (for example, DeKeyser and others, 2013; Ashton and others, 2016; Augustine and others, 2017; Porensky and others, 2018). Burned areas may have similar or lower cover of cheatgrass compared to unburned sites (Porensky and Blumenthal, 2016; Porensky and others, 2018; Symstad and others, 2021). Grassland site conditions associated with increased cheatgrass cover include the southeast aspect, sandier or rockier sites, or lower native species cover or richness (Ashton and others, 2016; Porensky and Blumenthal, 2016). Managers have limited control over a subset of factors that lead to increased annual brome cover or expansion (fig. 5; Ashton and others, 2016). Several factors like temperature, precipitation, and soils have strong direct and indirect effects on IAGs that constrain management decisions. In contrast, managers can have strong control over other factors that directly affect IAG abundance, such as fire and grazing. In addition, these factors can have interactive or species-specific effects on IAGs. For example, the timing and intensity of grazing can be used to decrease IAGs while maintaining native biomass, but the most effective combination of treatments has differed for cheatgrass as compared to Japanese brome (Vermeire and others, 2023).
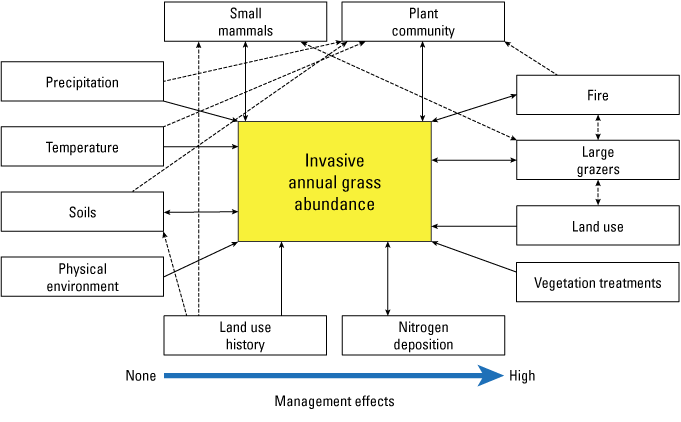
Conceptual diagram showing factors and associated direct (solid arrows), indirect (dashed arrows), or interactive (double arrows) management effects that lead to annual invasive grass abundance. Drivers of the model vary from more (right) or less (left) control by managers. Modified from Ashton and others (2016).
Climate change has the potential to alter community responses and drivers. In an experiment in Wyoming, cheatgrass biomass and seed production increased with treatments of raised temperatures (Blumenthal and others, 2016). In a greenhouse experiment, native blue bunch wheatgrass maintained a competitive balance over cheatgrass in several other treatment scenarios (Larson, Lehnhoff, and Rew, 2017; Larson and others, 201795). Shifts in temperature and precipitation may reduce the resistance of grassland sites to invasion by IAGs following disturbance or extreme weather events. Outcomes will likely be context dependent on factors like occurrence of competing species and may include feedback that results in novel ecosystem dynamics, such as altered fire cycles from increasing IAGs.
4.2.2. How Does Prescribed Fire Affect Invasive Annual Grass Distribution in Grassland Ecosystems?
Prescribed fire has been effective at reducing cover by cheatgrass and Japanese brome through direct mortality of individuals and seeds, whereas native perennial species rapidly recovered to prefire levels with spring burning (Whisenant and Uresk, 1990; Vermeire and Russell, 2018), summer burning (Vermeire and others, 2011, 2014; Vermeire and Russell, 2018), and fall burning (Vermeire and Russell, 2018; Symstad and others, 2021; Vermeire and others, 2021), even at low fuel loadings (Vermeire and Rinella, 2017). Spring burning also reduced Japanese brome in rangelands dominated by black greasewood in eastern Montana (Japanese brome was described as Bromus arvensis in Wade and others [2024]). However, the effectiveness of prescribed fire to reduce IAG cover has been demonstrated to decrease with increasing nonnative cover, such that fire alone may not be sufficient to control IAGs (Symstad and others, 2021), and declines in IAG cover may only last a few years. Therefore, direct effects (for example, remaining seedbank) and indirect effects (for example, available space for germination free from perennial grasses) can interact to govern the vegetation response after treatment.
Prescribed fire mimics wildfires that can control IAG distribution in grasslands of the northwestern Great Plains. Broad geographic studies on the grasslands of the Western United States (primarily the Great Plains) found that the number of large wildfires was higher during 2005–14 compared to 1985–94 (Donovan and others, 2017), but within a few years of a fire, most types of vegetation recovered quickly, except trees in the northern Great Plains (Donovan and others, 2020). Observational studies including wildfire found that longer fire return intervals were associated with higher IAG cover in national park units in the northern Great Plains (Ashton and others, 2016). Wildfire did not increase IAGs at the Thunder Basin National Grassland in northeastern Wyoming, where burned sites had similar or lower cheatgrass cover compared to unburned sites (Porensky and Blumenthal, 2016). Likewise, cheatgrass and Japanese brome were similarly low between burned and unburned sites less than or equal to 2 years following wildfires in Montana (Tulganyam and Carr, 2022). However, high cover by IAGs occurred in bulldozed firebreaks at these sites, indicating that such mechanical disturbances can increase IAGs in grasslands, which was also observed at firebreaks in sagebrush ecosystems (Seipel and others, 2018).
4.2.3. How Do Herbicide and Mechanical Treatments Affect Invasive Annual Grass Distribution in Grassland Ecosystems?
Some experiments have evaluated the responses of grassland sites to treatments such as mowing, changed grazing, or herbicide application, which can be used in conjunction with prescribed fire. A series of studies at Fort Keogh in southeastern Montana have evaluated different single treatments and combinations of treatments (refer to sec. 4.2.2 for prescribed fire alone treatments). For example, one study found that mowing had different effects than prescribed fire, resulting in higher IAG cover and decreased perennial grass cover (Vermeire and others, 2020), but Muscha and others (2023) saw a decrease in seed production of IAGs associated with clipping treatments that mimicked mowing in their greenhouse study. In this series of experiments, the herbicide (aminopyralid) variably reduced brome germination, but not biomass, through time (Vermeire and others, 2021). Although fire and herbicide did not demonstrate an interactive effect on brome biomass—or much of a difference compared to fire alone—the combination did reduce forb biomass (Vermeire and others, 2021). Conversely, different experimental designs found a compounding effect of prescribed fire plus herbicide (imazapic), especially at higher pretreatment annual brome cover values (Symstad and others, 2021). Other mowing experiments at Fort Keogh reduced Japanese brome cover through time and found no effect of fungicide that was applied to increase moss and exclude IAGs (Reinhart and others, 2020; Japanese brome was described as Bromus arvensis). Refer to section 5 “Design Features and Mitigation Measures for Controlling Invasive Annual Grasses” for effects of herbicide on annual bromes.
4.2.3. Additional Information to Enhance Understanding of Vegetation Treatments in Grassland Ecosystems
At least two gaps in information are evident based on grassland studies about IAGs. First, although prescribed fire is initially effective at reducing IAG cover, the effects may only last for a few years. Therefore, knowledge could be developed further on the durability and frequency of prescribed fire to effectively control IAGs throughout longer periods while avoiding other unintended vegetation community responses. Second, the combination of prescribed fire and herbicide may be a needed approach at some invasion levels and in some situations. However, the frequency, timing, and consequences to other vegetation functional groups (for example, shrubs and perennial grasses) in grassland communities are unknown. Longer-term control may center around the removal of litter (with infrequent fire or grazing for example) because some evidence shows that high litter loads foster seed germination (Whisenant and Uresk, 1990). However, there is a paucity of investigations on how to include other treatments, and the specific approaches likely to be effective across variable grasslands and for different objectives.
5. Design Features and Mitigation Measures for Controlling Invasive Annual Grasses
Section 5 Highlights
• Maintaining adequate cover of perennial vegetation creates rangelands that are resistant and resilient to annual grass invasions.
• Vegetation treatments can be a vector for invasion by IAGs, and focusing on treatment type, placement, and seasonal timing can limit invasion potential.
• Herbicidal mitigation measures perform best and can be used unaided or with another measure.
5.1. Conservation Strategies and Mitigation for Invasive Annual Grasses
Managers may follow several overarching strategies in conservation and restoration that affect the choice of mitigation measures. One strategy, known as “protecting the core,” prioritizes conservation of the most ecologically important areas (for example, sage-grouse leks, ungulate migratory corridors, and intact rangelands), where disturbances from IAGs, development, or other factors are minimal (Doherty and others, 2022; Maestas and others, 2022105). Regions that contain robust communities of native perennial grasses and limited disturbance are often more resilient to invasion by annual grasses (Chambers, Roundy, and others, 2007) and may require mitigation measures that differ from areas with abundant IAGs. Another strategy is known as “landscape triage” and prioritizes regions that have substantial perturbation but can be restored (Pyke, 2011). In landscape triage, intensive mitigation measures may be needed to achieve restoration objectives.
Because annual grasses can rapidly invade and expand their range, early detection and rapid response is a critical strategy to identify and eliminate isolated invasions before they become widespread and problematic (Reaser and others, 2020). In addition, it can be important to implement mitigation measures beyond the extent of the current invasion for long-term effectiveness. Barriers to mitigation such as private property or geopolitical boundaries can undermine mitigation measures, especially when seed sources for IAGs remain near project areas. Mitigation measures for annual grass invasions are integral to rangeland management and are often developed proactively to prevent invasions by annual grasses and reactively to limit or reduce existing IAGs in project areas.
5.2. Proactive Measures for Mitigating Invasive Annual Grasses
Proactive measures to prevent invasion by annual grasses are varied and may be most effective when used collectively. Important measures include seeding or planting native species and minimizing soil disturbances (for example, fire and road construction). Other measures include controlled moving of livestock; cleaning rangeland machinery after operating in weed-infested fields; and using weed-free hay, grain, straw, and mulch (Sheley and others, 2001). When seeding or haying, it is important to ensure that sources do not include IAGs (Lehan and others, 2013) because requirements for weed-free certification can vary based on whether species are listed as a “noxious weed” by a State or other factors. For example, the North Dakota Department of Agriculture requires that forage sources are inspected for medusahead, but not cheatgrass, ventenata, or Japanese brome, to be certified weed free based on the State’s noxious weed list (https://www.ndda.nd.gov/divisions/plant-industries/noxious-weeds) and guidelines of the North American Invasive Species Management Association (https://naisma.org/programs/weed-free-products/). In the focal region, ventenata and medusahead are listed as “noxious weeds” only in Montana and Wyoming; Japanese brome is not listed in any State; and cheatgrass is only listed as a “regulated weed” in Montana, where it is prohibited from intentional spread or being sold but allowed as a contaminant in agricultural products.
Managers can minimize increases in IAGs by placing vegetation treatments like conifer removals away from areas most likely to support IAGs. Environmental conditions that support IAGs (for example, aspect and elevation) have been described (refer to sec. 2 “Affected environment—Information on Possible Drivers of Invasion by Nonnative Annual Grasses and Data Sources to Characterize Their Distribution and Dynamics” and sec. 4 “Environmental Consequences—Information on the Potential Spread of Invasive Annual Grasses from Vegetation Treatments”), and resource managers can use tools like INHABIT to help identify areas with low risk of invasion by annual grasses. For example, cheatgrass and Japanese brome tend to be more successful invaders in dry, warm conditions that typically occur on south-facing lower-elevation sites (Brooks and others, 2016). Johnston (2011) found that natural dispersal of cheatgrass seeds averaged 2.4 m and ranged to 20.8 m, suggesting that proximity of vegetation treatments to existing IAGs is a critical factor for spreading IAGs.
Seasonal timing of treatments can also affect the potential for invasion by annual grasses (Harris, 1977; Harmoney, 2007; Reinhart and others, 2020; Vermeire and others, 2023). Vegetation treatments using herbicides or mechanical techniques may be best performed in early spring before most native perennial species are active and therefore not sensitive to disturbance at this time. Nonselective treatments (for example, prescribed burn and mowing) may be most effective when performed in fall, winter, or early spring after IAGs have emerged but before producing seed (Davies and others, 2021). These grasses gain a competitive advantage by germinating before others, which managers can minimize by treating areas during critical stages of growth by IAGs.
5.3. Reactive Measures for Mitigating Invasive Annual Grasses
Reactive measures for mitigating IAGs include herbicide, mechanical, biological (for example, grazing), and fire treatments that vary in cost, disturbance, and efficacy. These techniques can be applied in various combinations to increase effectiveness. Timing of application can be critical and determined by the phenology of the target species.
5.3.1. Herbicide Controls for Invasive Annual Grasses
The most common and often evaluated reactive measures used to control IAGs are herbicides (Masters and Sheley, 2001; DiTomaso and others, 2010) such as glyphosate, imidazolinone (for example, imazapic and imazapyr), and sulfonylurea (for example, chlorsulfuron, metsulfuron, rimsulfuron, and sulfometuron). Glyphosate is a nonselective, postemergent herbicide that is absorbed through the foliage and inhibits production of an enzyme that is important to plant growth. Conversely, imidazolinone and sulfonylurea herbicides are selective, postemergent herbicides that are absorbed through the foliage and roots and inhibit the synthesis of various amino acids (Masters and Sheley, 2001; DiTomaso and others, 2010). Although nonselective and selective herbicides have been shown to reduce IAG cover, nonselective herbicides have been more effective (Metier and others, 2019) and allow for the competitive release of perennial vegetation that can prevent future annual grass invasions. Commonly, managers apply multiple, complementary herbicides to increase effectiveness of mitigation measures (Johnston, 2015; Metier and others, 2019).
The BLM has evaluated effects of the active ingredients in many herbicides in programmatic environmental impact statements (BLM, 2007, 2016) and has permitted the use of aminopyralid, bromacil, chlorsulfuron, clopyralid, 2,4-D, dicamba, diquat, diuron, fluridone, fluroxypyr, glyphosate, hexazinone, imazapic, imazapyr, metsulfuron methyl, picloram, rimsulfuron, sulfometuron methyl, tebuthiuron, and triclopyr (BLM, 2007, 201628). A recent programmatic environmental impact statement includes evaluations for other active ingredients, including aminocyclopyrachlor, clethodim, fluazifop-p-butyl, flumioxazin, imazamox, indaziflam, oryzalin, and trifluralin (BLM, 2024).
Seasonal timing of herbicide application is important to mitigation effectiveness, especially for winter annual grasses with life cycles that start earlier than other grasses. Herbicides often have intended application timelines related to certain plant life-cycle stages. For example, imazapic is typically applied pre- or early postemergence in the fall to prevent emergence or kill new seedlings (Ehlert and others, 2015; Lehnhoff and others, 2019). In semiarid climates such as the northwestern Great Plains, these effects can persist into the spring (Ehlert and others, 2015). Conversely, indaziflam is intended to be applied pre-emergence because it does not affect seedlings, but instead affects a seed’s development of the radicle. However, this herbicide requires measurable precipitation following application for adequate distribution into the soil, and this distribution is inhibited by litter (Clark and others, 2019; Donaldson and Germino, 2022).
5.3.2. Mechanical, Biological, and Fire Treatments to Control Invasive Annual Grasses
Population persistence of IAGs is contingent upon the seed bank. However, seeds are typically viable for 1–2 years (Baskin and Baskin, 1981; Smith and others, 2008) and sensitive to burial depth (Wicks, 1997) and compaction (Thill and others, 1979). Mechanical control techniques that manipulate soils to disrupt germination and bury seeds (for example, disking and hoeing) or directly remove IAGs (for example, hand-pulling and mowing) can be effective. However, these techniques disturb and loosen soil, which can increase IAG occurrence, and are therefore most effective when accompanied by an herbicide (Masters and Sheley, 2001; Johnston 2015).
Biological methods, typically livestock grazing, are often used to control IAGs. Optimal treatment applications vary by grass species and ecoregion, and the effectiveness of these treatments depend on timing, intensity, and frequency of applications (Lehnhoff and others, 2019; Frost and others, 2023; Vermeire and others, 2023178). Grazing to control IAGs is most effective when it occurs before germination in late fall or after germination but before seed production in early spring (DiTomaso and others, 2008; Vermeire and others, 2023). By mid to late spring, IAGs produce seed that livestock may ingest or collect and distribute from their hooves, hair, or feces. Frost and others (2023) reported that grazing had no effect on IAGs or slightly increased IAGs in eastern Montana where responses varied with precipitation.
Biocontrol agents are natural enemies of IAGs in their native range that can be introduced to combat invasions. The effectiveness of biocontrol agents for IAGs may be substantially improved when combined with herbicide treatments (Lehnhoff and others, 2019). However, limited research on the biocontrol agent Pseudomonas fluorescens D7 in northeastern Wyoming suggests it is ineffective for controlling cheatgrass in field applications (Custer and others, 2024). Biocontrol agents that consume invasive plants can be imported as a control measure, but none are currently (2024) approved or available for use on IAGs described in this report (BLM, 2017).
Although fire can facilitate the proliferation, dispersal, and success of IAGs more than their native counterparts (Monty and others, 2012), prescribed fire can be a useful mitigation tool in some environments and circumstances. Most often, prescribed fire is used to reduce the seed bank and can be effective when applied in consecutive years and before plants produce seed (DiTomaso and others, 2006). Prescribed fire, like biological methods, can be ineffective when applied alone but can be coupled with herbicides, competitive reseeding, or other measures to increase success (Monaco and others, 2017).
Although most mitigating measures produce short-term reductions in IAG abundance, the effectiveness of singular applications typically diminishes through time (Monaco and others, 2017). As such, multiple treatments applied simultaneously or staggered shortly thereafter (for example, 1–3 years) can provide more sustained control. Competitive reseeding of perennial grasses through ground or aerial dispersal can improve long-term success in controlling IAGs, particularly in mesic environments, by establishing native communities that are resistant and resilient to invasion (Rinella and others, 2012).
Methods for Developing This Science Synthesis
Rutherford and others (2023) introduced a methodology for developing science syntheses to inform analyses conducted under the NEPA, and relevant text from that report is reproduced herein. This and other syntheses build on that foundation and methodology and apply it to new topics of management concern on western lands. We used a narrative review (Sovacool and others, 2018) and supplemental database search to collect science for this synthesis. We gathered data, science, analysis methods, and mitigation measures to inform environmental impact analyses on responses of IAGs to habitat treatments in shrub and grassland communities (Carter and others, 2023). In conducting the narrative review, we relied on our prior experience, publications, and work from key authors in the field of IAGs. We then conducted a literature review to supplement this approach.
We performed keyword literature searches in Scopus (https://www.scopus.com) and Web of Science (https://www.webofscience.com; accessed through the U.S. Geological Survey library) using the terms (“Montana” OR “North Dakota” OR “South Dakota” OR “Wyoming” OR “northwestern great plains” OR “intermontane grasslands”) AND (“brome” OR “bromus” OR “cheatgrass” OR “Japanese brome” OR “ventenata” OR “non-native annual grass*” OR “nonnative annual grass*” OR “invasive annual grass*” OR “exotic annual grass*”) AND (“treat*”). We conducted this search in June 2022 and considered research from any period, which resulted in 54 related publications in Scopus and 70 related publications in Web of Science. We repeated these searches in July 2024, which added four new publications on vegetation treatments in our focal region. In addition, we used snowballing methods (Wäldchen and Mäder, 2018) to expand our search using sources commonly listed in other publications. We included literature in our synthesis if it mentioned cheatgrass, downy brome, field brome, Japanese brome, ventenata, or other relevant Bromus species and if the study at least partially occurred in our geographic region of interest. Although literature on IAGs is plentiful, research on their responses to vegetation treatments is sparse, particularly in the northwestern Great Plains. Therefore, few of the publications from this search met all the criteria to be considered in this synthesis. Although our methods were designed to gather the best available literature about the effects of vegetation treatments on IAGs, this report is not intended to be a comprehensive literature review. In addition, it is possible that our literature search methods missed some relevant studies.
Throughout the development of this report, we worked with staff from the BLM, U.S. Fish and Wildlife Service, and U.S. Geological Survey to coproduce this report (Beier and others, 2017). We refined the structure and content of the report through close collaboration with multiple BLM staff throughout scoping, writing, and review.
References Cited
Adhikari, A., Mangold, J., Mainali, K.P., and Rew, L.J., 2023, Climate change and more disturbed land-use types will further the invasion of a non-native annual grass, Ventenata dubia: Biological Invasions, v. 25, p. 285–296, accessed August 25, 2024, at https://doi.org/10.1007/s10530-022-02913-4.
Allred, B.W., Bestelmeyer, B.T., Boyd, C.S., Brown, B., Davies, K.W., Duniway, M.C., Ellsworth, L.M., Erickson, T.A., Fuhlendorf, S.D., Griffiths, T.V., Jansen, V., Jones, M.O., Karl, J., Knight, A., Maestas, J.D., Maynard, J.J., McCord, S.E., Naugle, D.E., Starns, H.D., Twidwell, D., and Uden, D.R., 2021, Improving Landsat predictions of rangeland fractional cover with multitask learning and uncertainty: Methods in Ecology and Evolution, v. 12, no. 5, p. 841–849, accessed August 25, 2024, at https://doi.org/10.1111/2041-210X.13564.
Allred, B.W., Creutzburg, M.K., Carlson, J.C., Cole, C.J., Dovichin, C.M., Duniway, M.C., Jones, M.O., Maestas, J.D., Naugle, D.E., Nauman, T.W., Okin, G.S., Reeves, M.C., Rigge, M., Savage, S.L., Twidwell, D., Uden, D.R., and Zhou, B., 2022, Guiding principles for using satellite-derived maps in rangeland management: Rangelands, v. 44, no. 1, p. 78–86, accessed August 25, 2024, at https://doi.org/10.1016/j.rala.2021.09.004.
Ansong, M., and Pickering, C., 2013a, Are weeds hitchhiking a ride on your car? A systematic review of seed dispersal on cars: PLoS ONE, v. 8, no. 11, art. e80275, 11 p., accessed August 25, 2024, at https://doi.org/10.1371/journal.pone.0080275.
Ansong, M., and Pickering, C., 2013b, Long-distance dispersal of black spear grass (Heteropogon contortus) seed on socks and trouser legs by walkers in Kakadu National Park: Ecological Management & Restoration, v. 14, no. 1, p. 71–74, accessed August 25, 2024, at https://doi.org/10.1111/emr.12021.
Applestein, C., and Germino, M.J., 2021, Detecting shrub recovery in sagebrush steppe—Comparing Landsat-derived maps with field data on historical wildfires: Fire Ecology, v. 17, art. 5, 11 p., accessed August 25, 2024, at https://doi.org/10.1186/s42408-021-00091-7.
Applestein, C., and Germino. M., 2022a, How do accuracy and model agreement vary with versioning, scale, and landscape heterogeneity for satellite-derived vegetation maps in sagebrush steppe?: Ecological Indicators, v. 139, art. 108935, 11 p., accessed August 25, 2024, at https://doi.org/10.1016/j.ecolind.2022.108935.
Applestein, C., and Germino, M.J., 2022b, Patterns of post-fire invasion of semiarid shrub-steppe reveals a diversity of invasion niches within an exotic annual grass community: Biological Invasions, v. 24, p. 741–759, accessed August 25, 2024, at https://doi.org/10.1007/s10530-021-02669-3.
Applestein, C., and Germino, M., 2023, Satellite-derived plant cover maps vary in performance depending on version and product: Ecological Indicators, v. 155, art. 110950, 8 p., accessed August 25, 2024, at https://doi.org/10.1016/j.ecolind.2023.110950.
Ashton, I.W., Symstad, A.J., Davis, C.J., and Swanson, D.J., 2016, Preserving prairies—Understanding temporal and spatial patterns of invasive annual bromes in the northern Great Plains: Ecosphere, v. 7, no. 8, art. e01438, 20 p., accessed August 25, 2024, at https://doi.org/10.1002/ecs2.1438.
Augustine, D.J., Derner, J.D., Milchunas, D., Blumenthal, D., and Porensky, L.M., 2017, Grazing moderates increases in C3 grass abundance over seven decades across a soil texture gradient in shortgrass steppe: Journal of Vegetation Science, v. 28, no. 3, p. 562–572, accessed August 25, 2024, at https://doi.org/10.1111/jvs.12508.
Baskin, J.M., and Baskin, C.C., 1981, Ecology of germination and flowering in the weedy winter annual grass Bromus japonicus: Journal of Range Management, v. 34, no. 5, p. 369–372, accessed August 25, 2024, at http://hdl.handle.net/10150/646170. [Also available at https://doi.org/10.2307/3897906.]
Beck, J.L., Connelly, J.W., and Wambolt, C.L., 2012, Consequences of treating Wyoming big sagebrush to enhance wildlife habitats: Rangeland Ecology & Management, v. 65, no. 5, p. 444–455, accessed August 25, 2024, at https://doi.org/10.2111/REM-D-10-00123.1.
Beier, P., Hansen, L.J., Helbrecht, L., and Behar, D., 2017, A how-to guide for coproduction of actionable science: Conservation Letters, v. 10, no. 3, p. 288–296, accessed August 25, 2024, at https://doi.org/10.1111/conl.12300.
Bennett, J., Smart, A., and Perkins, L., 2019, Using phenological niche separation to improve management in a northern glaciated plains grassland: Restoration Ecology, v. 27, no. 4, p. 745–749, accessed August 25, 2024, at https://doi.org/10.1111/rec.12932.
Blank, R.R., Chambers, J.C., and Zamudio, D., 2003, Restoring riparian corridors with fire—Effects on soil and vegetation: Journal of Range Management, v. 56, no. 4, p. 388–396, accessed August 25, 2024, at http://hdl.handle.net/10150/643455. [Also available at https://doi.org/10.2307/4004044.]
Blumenthal, D.M., Norton, U., Derner, J.D., and Reeder, J.D., 2006, Long-term effects of tebuthiuron on Bromus tectorum: Western North American Naturalist, v. 66, no. 4, p. 420–425, accessed August 25, 2024, at https://scholarsarchive.byu.edu/wnan/vol66/iss4/2. [Also available at https://doi.org/10.3398/1527-0904(2006)66[420:LEOTOB]2.0.CO;2.]
Blumenthal, D.M., Kray, J.A., Ortmans, W., Ziska, L.H., and Pendall, E., 2016, Cheatgrass is favored by warming but not CO2 enrichment in a semi‐arid grassland: Global Change Biology, v. 22, no. 9, p. 3026–3038, accessed August 25, 2024, at https://doi.org/10.1111/gcb.13278.
Bradley, B.A., 2009, Regional analysis of the impacts of climate change on cheatgrass invasion shows potential risk and opportunity: Global Change Biology, v. 15, no. 1, p. 196–208, accessed August 25, 2024, at https://doi.org/10.1111/j.1365-2486.2008.01709.x.
Bradley, B.A., 2014, Remote detection of invasive plants—A review of spectral, textural and phenological approaches: Biological Invasions, v. 16, p. 1411–1425, accessed August 25, 2024, at https://doi.org/10.1007/s10530-013-0578-9.
Bradley, B.A., Curtis, C.A., and Chambers, J.C., 2016, Bromus response to climate and projected changes with climate change, in Germino, M., Chambers, J., and Brown, C., eds., Exotic brome-grasses in arid and semiarid ecosystems of the Western US—Causes, consequences, and management implications: Cham, Switzerland, Springer, p. 257–274, accessed August 25, 2024, at https://doi.org/10.1007/978-3-319-24930-8_9.
Brooks, M.L., Brown, C.S., Chambers, J.C., D’Antonio, C.M., Keeley, J.E., and Belnap, J., 2016, Exotic annual Bromus invasions—Comparisons among species and ecoregions in the Western United States, in Germino, M., Chambers, J., and Brown, C., eds., Exotic brome-grasses in arid and semiarid ecosystems of the Western US: Cham, Switzerland, Springer, p. 11–60, accessed August 25, 2024, at https://doi.org/10.1007/978-3-319-24930-8_2.
Brummer, T.J., Taylor, K.T., Rotella, J., Maxwell, B.D., Rew, L.J., and Lavin, M., 2016, Drivers of Bromus tectorum abundance in the Western North American sagebrush steppe: Ecosystems, v. 19, p. 986–1000, 15 p., accessed August 25, 2024, at https://doi.org/10.1007/s10021-016-9980-3.
Bukowski, B.B., and Baker, W.L., 2013, Historical fire regimes, reconstructed from land-survey data, led to complexity and fluctuation in sagebrush landscapes: Ecological Applications, v. 23, no. 3, p. 546–564, accessed August 25, 2024, at https://doi.org/10.1890/12-0844.1.
Bureau of Land Management [BLM], 2007, Vegetation treatments on Bureau of Land Management Lands in 17 Western States: Bureau of Land Management, 539 p., accessed August 25, 2024, at https://eplanning.blm.gov/eplanning-ui/project/70300/510.
Bureau of Land Management [BLM], 2014, 2,4-dicholorophenoxyacetic acid (2,4-D) ecological risk assessment final: Bureau of Land Management, 668 p., accessed August 25, 2024, at https://www.blm.gov/sites/blm.gov/files/BLM_ERA_24D_Final%20%283-28-14%29%20%281%29.pdf.
Bureau of Land Management [BLM], 2016, Final programmatic environmental impact statement for vegetation treatments using aminopyralid, fluroxypyr, and rimsulfuron on Bureau of Land Management Lands in 17 Western States: Bureau of Land Management, 5 p., accessed August 25, 2024, at https://eplanning.blm.gov/eplanning-ui/project/70301/510.
Bureau of Land Management [BLM], 2017, Programmatic analysis—Integrated weed management in the Billings Field Office: Bureau of Land Management NEPA no. DOI-BLM-MT-A010-2017-0004-EA, 41 p., accessed August 25, 2024, at https://eplanning.blm.gov/public_projects/nepa/79008/123388/150454/DOI-BLM-MT-A010-0004-EA.pdf.
Bureau of Land Management [BLM], 2021, BLM Natl AIM TerrADat Hub: Bureau of Land Management, accessed September 16, 2024, at https://gbp-blm-egis.hub.arcgis.com/datasets/BLM-EGIS::blm-natl-aim-terradat-hub/about.
Bureau of Land Management [BLM], 2024, Record of decision for programmatic approval addressing vegetation treatments using herbicides: Bureau of Land Management, 66 p., accessed August 25, 2024, at https://eplanning.blm.gov/eplanning-ui/project/2017138/570.
Carter, S.K., Haby, T.S., Meineke, J.K., Foster, A.C., McCall, L.E., Espy, L.D., Gilbert, M.A., Herrick, J.E., Prentice, K.L., 2023, Prioritizing science efforts to inform decision making on public lands: Frontiers in Ecology and the Environment, v. 21, no. 10, p. 453–460, accessed August 25, 2024, at https://doi.org/10.1002/fee.2672.
Chambers, J.C., Beck, J.L., Campbell, S., Carlson, J., Christiansen, T.J., Clause, K.J., Dinkins, J.B., Doherty, K.E., Griffin, K.A., Havlina, D.W., Henke, K.F., Hennig, J.D., Kurth, L.L., Maestas, J.D., Manning, M., Mayer, K.E., Mealor, B.A., McCarthy, C., Perea, M.A., and Pyke, D.A., 2016, Using resilience and resistance concepts to manage threats to sagebrush ecosystems, Gunnison sage-grouse, and greater sage-grouse in their eastern range—A strategic multi-scale approach: Fort Collins, Colo., U.S. Department of Agriculture Forest Service Rocky Mountain Research Station General Technical Report RMRS-GTR-356, 143 p., accessed August 25, 2024, at https://doi.org/10.2737/RMRS-GTR-356.
Chambers, J.C., Brooks, M.L., Germino, M.J., Maestas, J.D., Board, D.I., Jones, M.O., and Allred, B.W., 2019. Operationalizing resilience and resistance concepts to address invasive grass-fire cycles: Frontiers in Ecology and Evolution, v. 7, art. 185, 25 p., accessed August 25, 2024, at https://doi.org/10.3389/fevo.2019.00185.
Chambers, J.C., Brown, J.L., Bradford, J.B., Board, D.I., Campbell, S.B., Clause, K.J., Hanberry, B., Schlaepfer, D.R., and Urza, A.K., 2023, New indicators of ecological resilience and invasion resistance to support prioritization and management in the sagebrush biome, United States: Frontiers in Ecology and Evolution, v. 10, art. 1009268, 17 p., accessed August 25, 2024, at https://doi.org/10.3389/fevo.2022.1009268.
Chambers, J.C., Germino, M.J., Belnap, J., Brown, C.S., Schupp, E.W., and Clair, S.B.S., 2016, Plant community resistance to invasion by Bromus species—the roles of community attributes, Bromus interactions with plant communities, and Bromus traits, in Germino, M., Chambers, J., and Brown, C., eds., Exotic brome-grasses in arid and semiarid ecosystems of the Western US: Cham, Switzerland, Springer, p. 275–304, accessed August 25, 2024, at https://doi.org/10.1007/978-3-319-24930-8_10.
Chambers, J.C., Maestas, J.D., Pyke, D.A., Boyd, C.S., Pellant, M., and Wuenschel, A., 2017, Using resilience and resistance concepts to manage persistent threats to sagebrush ecosystems and greater sage-grouse: Rangeland Ecology & Management, v. 70, no. 2, p. 149–164, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2016.08.005.
Chambers, J.C., Miller, R.F., Board, D.I., Pyke, D.A., Roundy, B.A., Grace, J.B., Schupp, E.W., and Tausch, R.J., 2014, Resilience and resistance of sagebrush ecosystems—Implications for state and transition models and management treatments: Rangeland Ecology & Management, v. 67, no. 5, p. 440–454, accessed August 25, 2024, at https://doi.org/10.2111/REM-D-13-00074.1.
Chambers, J.C., Pyke, D.A., Maestas, J.D., Pellant, M., Boyd, C.S., Campbell, S.B., Espinosa, S., Havlina, D.W., Mayer, K.E., and Wuenschel, A., 2014, Using resistance and resilience concepts to reduce impacts of invasive annual grasses and altered fire regimes on the sagebrush ecosystem and greater sage-grouse—A strategic multi-scale approach: Fort Collins, Colo., U.S. Department of Agriculture Forest Service Rocky Mountain Research Station General Technical Report RMRS-GTR-326, 73 p., accessed August 25, 2024, at https://doi.org/10.2737/RMRS-GTR-326.
Chambers, J.C., Roundy, B.A., Blank, R.R., Meyer, S.E., and Whittaker, A., 2007, What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum?: Ecological Monographs, v. 77, no. 1, p. 117–145, accessed August 25, 2024, at https://doi.org/10.1890/05-1991.
Chambers, J.C., Strand, E.K., Ellsworth, L.M., Tortorelli, C.M., Urza, A.K., Crist, M.R., Miller, R.F., Reeves, M.C., Short, K.C., and Williams, C.L., 2024, Review of fuel treatment effects on fuels, fire behavior and ecological resilience in sagebrush (Artemisia spp.) ecosystems in the Western U.S.: Fire Ecology, v. 20, art. 32, 36 p., accessed August 25, 2024, at https://doi.org/10.1186/s42408-024-00260-4.
Clark, S.L., da Silva, P.V., Dayan, F.E., Nissen, S.J., and Sebastian, D.J., 2019, The influence of winter annual grass litter on herbicide availability: Weed Science, v. 67, no. 6, p. 702–709, accessed August 25, 2024, at https://doi.org/10.1017/wsc.2019.45.
Custer, G.F., Mealor, B.A., Fowers, B., and van Diepen, L.T.A., 2024, Soil microbiome analysis supports claims of ineffectiveness of Pseudomonas fluorescens D7 as a biocontrol agent of Bromus tectorum: Microbiology Spectrum, v. 12, no. 1, p. 1–5, accessed August 25, 2024, at https://doi.org/10.1128/spectrum.01771-23.
D’Antonio, C.M., and Vitousek, P.M., 1992, Biological invasions by exotic grasses, the grass/fire cycle, and global change: Annual Review of Ecology, Evolution, and Systematics, v. 23, p. 63–87, accessed August 25, 2024, at https://doi.org/10.1146/annurev.es.23.110192.000431.
Dahal, D., Pastick, N.J., Boyte, S.P., Parajuli, S., Oimoen, M.J., and Megard, L.J., 2022, Multi-species inference of exotic annual and native perennial grasses in rangelands of the Western United States using harmonized Landsat and Sentinel-2 data: Remote Sensing, v. 14, no. 4, art. 807, 21 p., accessed August 25, 2024, at https://doi.org/10.3390/rs14040807.
Dahlgren, D.K., Chi, R., and Messmer, T.A., 2006, Greater sage-grouse response to sagebrush management in Utah: Wildlife Society Bulletin, v. 34, no. 4, p. 975–985, accessed August 25, 2024, at https://doi.org/10.2193/0091-7648(2006)34[975:GSRTSM]2.0.CO;2.
Davies, K.W., Bates, J.D., Perryman, B., and Arispe, S., 2021, Fall-winter grazing after fire in annual grass-invaded sagebrush steppe reduced annuals and increased a native bunchgrass: Rangeland Ecology & Management, v. 77, p. 1–8, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2021.03.001.
Davies, K.W., Svejcar, T.J., and Bates, J.D., 2009, Interaction of historical and nonhistorical disturbances maintains native plant communities, Ecological Applications, v. 19, no. 6, p. 1536–1545, accessed August 25, 2024, at https://doi.org/10.1890/09-0111.1.
Davis, M.A., Grime, J.P., and Thompson, K., 2000, Fluctuating resources in plant communities—A general theory of invasibility: Journal of Ecology, v. 88, no. 3, p. 528–534, accessed August 25, 2024, at https://doi.org/10.1046/j.1365-2745.2000.00473.x.
Derner, J.D., Wilmer, H., Stackhouse-Lawson, K., Place, S., Boggess, M., 2023, Practical considerations for adaptive strategies by US grazing land managers with a changing climate: Agrosystems, Geosciences & Environment, v. 6, no. 1, art. e20356, 13 p., accessed July 3, 2025, at https://doi.org/10.1002/agg2.20356.
DeKeyser, E.S., Meehan, M., Clambey, G., and Krabbenhoft, K., 2013, Cool season invasive grasses in northern Great Plains natural areas: Natural Areas Journal, v. 33, no. 1, p. 81–90, accessed August 25, 2024, at https://doi.org/10.3375/043.033.0110.
De Stefano, A., Mealor, B.A., Jones, L.C., Lehnhoff, E.A., Mangold, J.M., Prather, T., Ransom, C.V., and Rew, L.J., 2024, Plot-measured variables indicate landscape-scale patterns of annual grass invasion in northwestern US rangelands: Rangeland Ecology & Management, v. 92, p. 90–99, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2023.10.003.
DiTomaso, J.M., Brooks, M.L., Allen, E.B., Minnich, R., Rice, P.M., and Kyser, G.B., 2006, Control of invasive weeds with prescribed burning: Weed Technology, v. 20, no. 2, p. 535–548, accessed August 25, 2024, at https://doi.org/10.1614/WT-05-086R1.1.
DiTomaso, J.M., Kyser, G.B., George, M.R., Doran, M.P., and Laca, E.A., 2008, Control of medusahead (Taeniatherum caput-medusae) using timely sheep grazing: Invasive Plant Science and Management, v. 1, no. 3, p. 241–247, accessed August 25, 2024, at https://doi.org/10.1614/IPSM-07-031.1.
DiTomaso, J.M., Masters, R.A., and Peterson, V.F., 2010, Rangeland invasive plant management: Rangelands, v. 32, no. 1, p. 43–47, accessed August 25, 2024, at https://doi.org/10.2111/RANGELANDS-D-09-00007.1.
Doherty, K., Theobald, D.M., Bradford, J.B., Wiechman, L.A., Bedrosian, G., Boyd, C.S., Cahill, M., Coates, P.S., Creutzburg, M.K., Crist, M.R., Finn, S.P., Kumar, A.V., Littlefield, C.E., Maestas, J.D., Prentice, K.L., Prochazka, B.G., Remington, T.E., Sparklin, W.D., Tull, J.C., Wurtzebach, Z., and Zeller, K.A., 2022, A sagebrush conservation design to proactively restore America’s sagebrush biome: U.S. Geological Survey Open-File Report 2022–1081, 38 p., accessed August 25, 2024, at https://doi.org/10.3133/ofr20221081.
Donaldson, R., and Germino, M.J., 2022, Intra-site sources of restoration variability in severely invaded rangeland—Strong temporal effects of herbicide-weather interactions; weak spatial effects of plant community patch type and litter: Ecological Solutions and Evidence, v. 3, no. 3, art. e12172, 14 p., accessed August 25, 2024, at https://doi.org/10.1002/2688-8319.12172.
Donovan, V.M., Twidwell, D., Uden, D.R., Tadesse, T., Wardlow, B.D., Bielski, C.H., Jones, M.O., Allred, B.W., Naugle, D.E., and Allen, C.R., 2020, Resilience to large, “catastrophic” wildfires in North America's grassland biome: Earth's Future, v. 8, no. 7, art. e2020EF001487, 16 p., accessed August 25, 2024, at https://doi.org/10.1029/2020EF001487.
Donovan, V.M., Wonkka, C.L., and Twidwell, D., 2017, Surging wildfire activity in a grassland biome: Geophysical Research Letters, v. 44, no. 12, p. 5986–5993, accessed August 25, 2024, at https://doi.org/10.1002/2017GL072901.
Dornbusch, M.J., Limb, R.F., and Sedivec, K.K., 2020, Alternative grazing management strategies combat invasive grass dominance: Natural Areas Journal, v. 40, no. 1, p. 86–95 accessed August 25, 2024, at https://doi.org/10.3375/043.040.0110.
Ehlert, K.A., Engel, R.E., and Mangold, J.M., 2015, Imazapic activity in a semiarid climate in downy brome (Bromus tectorum)–infested rangeland and CRP sites: Weed Technology, v. 29, no. 3, p. 472–479, accessed August 25, 2024, at https://doi.org/10.1614/WT-D-14-00078.1.
Ellsworth, L.M., and Kauffman, J.B., 2017, Plant community response to prescribed fire varies by pre-fire condition and season of burn in mountain big sagebrush ecosystems: Journal of Arid Environments, v. 144, p. 74–80, accessed August 25, 2024, at https://doi.org/10.1016/j.jaridenv.2017.04.012.
Engelstad, P., Jarnevich, C.S., Hogan, T., Sofaer, H.R., Pearse, I.S., Sieracki, J.L., Frakes, N., Sullivan, J., Young, N.E., Prevéy, J.S., Belamaric, P., and LaRoe, J., 2022, INHABIT—A web-based decision support tool for invasive plant species habitat visualization and assessment across the contiguous United States: PLoS ONE, v. 17, no. 2, art. e0263056, 15 p., accessed August 25, 2024, at https://doi.org/10.1371/journal.pone.0263056.
Fox, L.B., Arsenault, A.A., Brewer, C.E., Carpenter, L.H., Jellison, B., Jenks, J.A., Jensen, W.F., Keegan, T.W., Kraft, D.J., Lutz, D.W., Richardson, C.L., Trindle, B.D., Schmidt, A.P., and Stivers, T.S., 2009, Habitat guidelines for mule deer—Great Plains ecoregion: Mule Deer Working Group, Western Association of Fish and Wildlife Agencies, 64 p., accessed March 24, 2025, at https://wafwa.org/wp-content/uploads/2020/08/HabitatGuidelines_MuleDeer_Great_Plains.pdf.
Fridley, J.D., Stachowicz, J.J., Naeem, S., Sax, D.F., Seabloom, E.W., Smith, M.D., Stohlgren, T.J., Tilman, D., and Von Holle, B., 2007, The invasion paradox—Reconciling pattern and process in species invasions: Ecology, v. 88, no. 1, p. 3–17, accessed August 25, 2024, at https://doi.org/10.1890/0012-9658(2007)88[3:TIPRPA]2.0.CO;2.
Frost, M.D.T., Komatsu, K.J., Porensky, L.M., Reinhart, K.O., Wilcox, K.R., and Koerner, S.E., 2023, Consequences of rainfall manipulations for invasive annual grasses vary across grazed northern mixed-grass prairie sites: Rangeland Ecology & Management, v. 90, p. 1–12, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2023.05.007.
Fusco, E.J., Balch, J.K., Mahood, A.L., Nagy, R.C., Syphard, A.D., and Bradley, B.A., 2021, The human–grass–fire cycle—How people and invasives co‐occur to drive fire regimes: Frontiers in Ecology and the Environment, v. 20, no. 2, p. 117–126, accessed August 25, 2024, at https://doi.org/10.1002/fee.2432.
Fusco, E.J., Finn, J.T., Balch, J.K., Nagy, R.C., and Bradley, B.A., 2019, Invasive grasses increase fire occurrence and frequency across US ecoregions: Proceedings of the National Academy of Sciences, v. 116, no. 47, p. 23594–23599, accessed August 25, 2024, at https://doi.org/10.1073/pnas.1908253116.
Gannon, J.J., Shaffer, T.L., and Moore, C.T., 2013, Native prairie adaptive management—A multi region adaptive approach to invasive plant management on Fish and Wildlife Service owned native prairies: U.S. Geological Survey Open File Report 2013–1279, 184 p., accessed August 25, 2024, at https://doi.org/10.3133/ofr20131279.
Gaskin, J.F., Espeland, E., Johnson, C.D., Larson, D.L., Mangold, J.M., McGee, R.A., Milner, C., Paudel, S., Pearson, D.E., Perkins, L.B., Prosser, C.W., Runyon, J.B., Sing, S.E., Sylvain, Z.A., Symstad, A.J., and Tekiela, D.R., 2021, Managing invasive plants on Great Plains grasslands—A discussion of current challenges: Rangeland Ecology & Management, v. 78, p. 235–249, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2020.04.003.
Germino, M.J., Chambers, J.C., and Brown, C.S., 2016a, Introduction—Exotic annual Bromus in the Western USA, in Germino, M., Chambers, J., and Brown, C., ed., Exotic brome-grasses in arid and semiarid ecosystems of the Western US: Cham, Switzerland, Springer, p. 1–7 accessed August 25, 2024, at https://doi.org/10.1007/978-3-319-24930-8_1.
Germino, M.J., Chambers, J.C., and Brown, C.S., eds., 2016b, Exotic brome-grasses in arid and semiarid ecosystems of the Western US: Cham, Switzerland, Springer, 475 p., accessed August 25, 2024, at https://link.springer.com/book/10.1007/978-3-319-24930-8.
Germino, M.J., Belnap, J., Stark, J.M., Allen, E.B., and Rau, B.M., 2016, Ecosystem impacts of exotic annual invaders in the genus Bromus, in Germino, M., Chambers, J., and Brown, C., eds., Exotic brome-grasses in arid and semiarid ecosystems of the Western US: Cham, Switzerland, Springer, p. 61–95 accessed August 25, 2024, at https://doi.org/10.1007/978-3-319-24930-8_3.
Griepentrog, H.W., Noerremark, M., Nielsen, J., and Ibarra, J.S., 2007, Autonomous inter-row hoeing using GPS-based side-shift control: Agricultural Engineering International—The CIGR Ejournal, v. 9, art. ATOE 07 005, 9 p., accessed August 25, 2024, at https://cigrjournal.org/index.php/Ejounral/article/view/936/930.
Guyette, R.P., Stambaugh, M.C., Marschall, J., and Abadir, E., 2015, An analytic approach to climate dynamics and fire frequency in the Great Plains: Great Plains Research, v. 25, no. 2, p. 139–150, accessed August 25, 2024, at https://www.jstor.org/stable/44685204. [Also available at https://doi.org/10.1353/gpr.2015.0031.]
Haferkamp, M.R., Volesky, J.D., Borman, M.M., Heitschmidt, R.K., and Currie, P.O., 1993, Effects of mechanical treatments and climatic factors on the productivity of northern Great Plains rangelands: Journal of Range Management, v. 46, no. 4, p. 346–350, accessed August 25, 2024, at http://hdl.handle.net/10150/644547. [Also available at https://doi.org/10.2307/4002470.]
Harmoney, K.R., 2007, Grazing and burning Japanese brome (Bromus japonicus) on mixed grass rangelands: Rangeland Ecology & Management, v. 60, no. 5, p. 479–486, accessed August 25, 2024, at https://doi.org/10.2111/1551-5028(2007)60[479:GABJBB]2.0.CO;2.
Harris, G.A., 1977, Root phenology as a factor of competition among grass seedlings: Journal of Range Management, v. 30, no. 3, p. 172–177, accessed August 25, 2024, at http://hdl.handle.net/10150/646796. [Also available at https://doi.org/10.2307/3897461.]
Heller, M., 2021, North American central grasslands: U.S. Fish and Wildlife Service web page, accessed September 16, 2024, at https://gs-portal-fws.hub.arcgis.com/datasets/fws::north-american-central-grasslands/about.
Herrick, J.E., Van Zee, J.W., McCord, S.E., Courtright, E.M., Karl, J.W., and Burkett, L.M., 2017, Core methods, v. 1 of Monitoring manual for grassland, shrubland and savanna ecosystems (2d ed.): Las Cruces, New Mex., U.S. Department of Agriculture Agricultural Research Service Jornada Experimental Range, 86 p., accessed August 25, 2024, at https://www.landscapetoolbox.org/wp-content/uploads/2022/05/MMGSSE_vol1_508.pdf.
Homer, C.T., Xian, G., Aldridge, C.L., Meyer, D.K., Loveland, T.R., and O’Donnell, M.S., 2015, Forecasting sagebrush ecosystem components and greater sage-grouse habitat for 2050—Learning from past climate patterns and Landsat imagery to predict the future: Ecological Indicators, v. 55, p. 131–145, accessed August 25, 2024, at https://doi.org/10.1016/j.ecolind.2015.03.002.
Innes, R.J., and Zouhar, K., 2018, Fire regimes of mountain big sagebrush communities: U.S. Department of Agriculture Forest Service Rocky Mountain Research Station Missoula Fire Sciences Laboratory web page, accessed August 25, 2024, at https://www.fs.usda.gov/database/feis/fire_regimes/mountain_big_sagebrush/all.html.
Jarnevich, C.S., and Young, N., 2015, Using the MAXENT program for species distribution modelling to assess invasion risk, in Venette, R.C., ed., Pest risk modelling and mapping for invasive alien species: CABI Digital Library, p. 65–81, accessed August 25, 2024, at https://www.cabidigitallibrary.org/doi/10.1079/9781780643946.0065.
Jeffries, M.I., and Finn, S.P., 2019, The sagebrush biome range extent, as derived from classified Landsat imagery: U.S. Geological Survey data release, accessed September 16, 2024, at https://doi.org/10.5066/P950H8HS.
Johnson, K.H., Olson, R.A., and Whitson, T.D., 1996, Composition and diversity of plant and small mammal communities in tebuthiuron-treated big sagebrush (Artemisia tridentata): Weed Technology, v. 10, no. 2, p. 404–416, accessed August 25, 2024, at https://doi.org/10.1017/S0890037X0004015X.
Johnston, D.B., 2011, Movement of weed seeds in reclamation areas: Restoration Ecology, v. 19, no. 4, p. 446–449, accessed August 25, 2024, at https://doi.org/10.1111/j.1526-100X.2011.00785.x.
Johnston, D.B., 2015, Downy brome (Bromus tectorum) control for pipeline restoration: Invasive Plant Science and Management, v. 8, no. 2, p. 181–192, accessed August 25, 2024, at https://doi.org/10.1614/IPSM-D-14-00001.1.
Johnston, A.N., Beever, E.A., Merkle, J.A., and Chong, G., 2018, Vegetation responses to sagebrush-reduction treatments measured by satellites: Ecological Indicators, v. 87, p. 66–76, accessed August 25, 2024, at https://doi.org/10.1016/j.ecolind.2017.12.033.
Jones, L.C., Norton, N., and Prather, T.S., 2018, Indicators of ventenata (Ventenata dubia) invasion in sagebrush steppe rangelands: Invasive Plant Science and Management, v. 11, no. 1, p. 1–9, accessed August 25, 2024, at https://doi.org/10.1017/inp.2018.7.
Jones, M.O., Allred, B.W., Naugle, D.E., Maestas, J.D., Donnelly, P., Metz, L.J., Karl, J., Smith, R., Bestelmeyer, B., Boyd, C., Kerby, J.D., and McIver, J.D., 2018, Innovation in rangeland monitoring—Annual, 30 m, plant functional type percent cover maps for U.S. rangelands, 1984–2017: Ecosphere, v. 9, no. 9, art. e02430, 19 p., accessed August 25, 2024, at https://doi.org/10.1002/ecs2.2430.
Kachergis, E., Miller, S.W., McCord, S.E., Dickard, M., Savage, S., Reynolds, L.V., Lepak, N., Dietrich, C., Green, A., Nafus, A., Prentice, K., and Davidson, Z., 2022, Adaptive monitoring for multiscale land management—Lessons learned from the Assessment, Inventory, and Monitoring (AIM) principles: Rangelands, v. 44, no. 1, p. 50–63, accessed August 25, 2024, at https://doi.org/10.1016/j.rala.2021.08.006.
Knapp, P.A., 1996, Cheatgrass (Bromus tectorum L) dominance in the Great Basin desert—History, persistence, and influences to human activities: Global Environmental Change, v. 6, no. 1, p. 37–52, accessed August 25, 2024, at https://doi.org/10.1016/0959-3780(95)00112-3.
Kobiela, B., Quast, J., Dixon, C., and DeKeyser, E.S., 2017, Targeting introduced species to improve plant community composition on USFWS-managed prairie remnants: Natural Areas Journal, v. 37, no. 2, p. 150–160, accessed August 25, 2024, at https://doi.org/10.3375/043.037.0204.
Lacey, J.R., Mowbray, J.E., and Wight, J.R., 1981, Ranchers evaluate contour furrows for livestock grazing: Rangelands, v. 3, no. 2, p. 58–60, accessed August 25, 2024, at http://hdl.handle.net/10150/638269.
Larson, C.D., Lehnhoff, E.A., Noffsinger, C., and Rew, L.J., 2017, Competition between cheatgrass and bluebunch wheatgrass is altered by temperature, resource availability, and atmospheric CO2 concentration: Oecologia, v. 186, p. 855–868, accessed August 25, 2024, at https://doi.org/10.1007/s00442-017-4046-6.
Larson, C.D., Lehnhoff, E.A., and Rew, L.J., 2017, A warmer and drier climate in the northern sagebrush biome does not promote cheatgrass invasion or change its response to fire: Oecologia, v. 185, no. 4, p. 763–774, accessed August 25, 2024, at https://doi.org/10.1007/s00442-017-3976-3.
Lavin, M., Brummer, T.J., Quire, R., Maxwell, B.D., and Rew, L.J., 2013, Physical disturbance shapes vascular plant diversity more profoundly than fire in the sagebrush steppe of southeastern Idaho, USA: Ecology and Evolution, v. 3, no. 6, p. 1626–1641, accessed August 25, 2024, at https://doi.org/10.1002/ece3.574.
Lehan, N.E., Murphy, J.R., Thorburn, L.P., and Bradley, B.A., 2013, Accidental introductions are an important source of invasive plants in the continental United States: American Journal of Botany, v. 100, no. 7, p. 1287–1293, accessed August 25, 2024, at https://doi.org/10.3732/ajb.1300061.
Lehnhoff, E.A., Rew, L.J., Mangold, J.M., Seipel, T., and Ragen, D., 2019, Integrated management of cheatgrass (Bromus tectorum) with sheep grazing and herbicide: Agronomy, v. 9, no. 6, art. 315, 21 p., accessed August 25, 2024, at https://doi.org/10.3390/agronomy9060315.
Lohse, K.A., Pierson, D., Patton, N.R., Sanderman, J., Huber, D.P., Finney, B., Facer, J., Meyers, J., and Seyfried, M.S., 2022, Multiscale responses and recovery of soils to wildfire in a sagebrush steppe ecosystem: Scientific Reports, v. 12, art. 22438, 12 p., accessed August 25, 2024, at https://doi.org/10.1038/s41598-022-26849-w.
Lonsdale, W.M., 1999, Global patterns of plant invasions and the concept of invasibility: Ecology, v. 80, no. 5, p. 1522–1536, accessed August 25, 2024, at https://doi.org/10.1890/0012-9658(1999)080[1522:GPOPIA]2.0.CO;2.
Mack, R.N., 2011, Fifty years of ‘waging war on cheatgrass’—Research advances, while meaningful control languishes, chap. 19 of Richardson, D.M., ed., Fifty years of invasion ecology—The legacy of Charles Elton: Wiley-Blackwell, Oxford, United Kingdom, Blackwell Publishing Ltd, p. 253–265, accessed August 25, 2024, at https://doi.org/10.1002/9781444329988.ch19.
Mack, R.N., Simberloff, D., Lonsdale, W.M., Evans, H., Clout, M., and Bazzaz, F.A., 2000, Biotic invasions—Causes, epidemiology, global consequences, and control: Ecological Applications, v. 10, no. 3, p. 689–710, accessed August 25, 2024, at https://doi.org/10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2.
Maestas, J., Jones, M., Pastick, N.J., Rigge, M.B., Wylie, B.K., Garner, L., Crist, M., Homer, C., Boyte, S., and Witacre, B., 2020, Annual herbaceous cover across rangelands of the sagebrush biome: U.S. Geological Survey data release, accessed August 25, 2024, at https://doi.org/10.5066/P9VL3LD5.
Maestas, J.D., Porter, M., Cahill, M., and Twidwell, D., 2022, Defend the core—Maintaining intact rangelands by reducing vulnerability to invasive annual grasses: Rangelands, v. 44, no. 3, p. 181–186, accessed August 25, 2024, at https://doi.org/10.1016/j.rala.2021.12.008.
Masters, R.A., and Sheley, R.L., 2001, Principles and practices for managing rangeland invasive plants: Journal of Range Management, v. 54, no. 5, p. 502–517, accessed August 25, 2024, at http://hdl.handle.net/10150/643585. [Also available at https://doi.org/10.2307/4003579.]
McIver, J., Brunson, M., Bunting, S., Chambers, J., Doescher, P., Grace, J., Hulet, A., Johnson, D., Knick, S., Miller, R., Pellant, M., Pierson, F., Pyke, D., Rau, B., Rollins, K., Roundy, B., Schupp, E., Tausch, R., and Williams, J., 2014, A synopsis of short-term response to alternative restoration treatments in sagebrush-steppe—The SageSTEP Project: Rangeland Ecology & Management, v. 67, no. 5, p. 584–598, accessed August 25, 2024, at https://doi.org/10.2111/REM-D-14-00084.1.
McKenzie, D., Jensen, F.R., Johnsen, T.N., Jr., and Young, J.A., 1984, Chains for mechanical brush control: Rangelands, v. 6, no. 3, p. 122–127, accessed August 25, 2024, at http://hdl.handle.net/10150/638508.
McMahon, D.E., Urza, A.K., Brown, J.L., Phelan, C., Chambers, J.C., 2021, Modelling species distributions and environmental suitability highlights risk of plant invasions in Western United States: Diversity and Distributions, v. 27, no. 4, p. 710–728, accessed September 16, 2024, at https://doi.org/10.1111/ddi.13232.
Meineke, J.K., Maxwell, L.M., Foster, A.C., McCall, L.E., Rutherford, T.K., Samuel, E.M., Selby, L.B., Willems, J.S., Kleist, N.J., and Jordan, S.E., 2024, Annotated bibliography of scientific research on Taeniatherum caput-medusae published from January 2010 to January 2022: U.S. Geological Survey Open-File Report 2023–1089, 164 p., accessed September 9, 2024, at https://doi.org/10.3133/ofr20231089.
Metier, E.P., Lehnhoff, E.A., Mangold, J., Rinella, M.J., and Rew, L.J., 2019, Control of downy brome (Bromus tectorum) and Japanese brome (Bromus japonicus) using glyphosate and four graminicides—Effects of herbicide rate, plant size, species, and accession: Weed Technology, v. 34, no. 2, p. 284–291, accessed August 25, 2024, at https://doi.org/10.1017/wet.2019.112.
Monaco, T.A., Mangold, J.M., Mealor, B.A., Mealor, R.D., and Brown, C.S., 2017, Downy brome control and impacts on perennial grass abundance—A systematic review spanning 64 years: Rangeland Ecology & Management, v. 70, no. 3, p. 396–404, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2016.09.008.
Montana Natural Heritage Program, 2021a, Bromus tectorum (cheatgrass) predicted suitable habitat modeling: Helena, Mont., Montana Natural Heritage Program, 17 p., accessed August 25, 2024, at https://mtnhp.org/models/files/Bromus_tectorum_PMPOA151H0_20210222.pdf.
Montana Natural Heritage Program, 2021b, Bromus japonicus (Japanese brome) predicted suitable habitat modeling: Helena, Mont., Montana Natural Heritage Program, 17 p., accessed August 25, 2024, at https://mtnhp.org/models/files/Bromus_japonicus_PMPOA150M0_20210525.pdf.
Montana Natural Heritage Program, 2021c, Ventenata dubia (ventenata) predicted suitable habitat modeling: Helena, Mont., Montana Natural Heritage Program, 17 p., accessed August 25, 2024, at https://mtnhp.org/models/files/Ventenata_dubia_PMPOA6D010_20240405.pdf.
Montana Natural Heritage Program, 2024a, Montana field guides—Field brome—Bromus arvensis: mt.gov web page, accessed September 9, 2024, at https://FieldGuide.mt.gov/speciesDetail.aspx?elcode=PMPOA15060.
Montana Natural Heritage Program, 2024b, Montana field guides—Great Plains mixedgrass prairie: mt.gov web page, accessed September 16, 2024, at https://FieldGuide.mt.gov/displayES_Detail.aspx?ES=7114.
Montana Natural Heritage Program, 2024c, Montana field guides—Montane sagebrush steppe: mt.gov web page, accessed September 16, 2024, at https://FieldGuide.mt.gov/displayES_Detail.aspx?ES=5455.
Monty, A., Brown, C.S., and Johnston, D.B., 2012, Fire promotes downy brome (Bromus tectorum L.) seed dispersal: Biological Invasions, v. 15, p. 1113–1123, accessed August 25, 2024, at https://doi.org/10.1007/s10530-012-0355-1.
Muscha, J.M., Vermeire, L.T., and Haferkamp, M.R., 2023, Clipping height and frequency effects on Japanese brome seed production and viability: Rangeland Ecology & Management, v. 90, p. 290–293, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2023.04.003.
Natural Resources Conservation Service, 2005, Light disking to enhance early successional wildlife habitat in grasslands and old fields—Wildlife benefits and erosion potential: Natural Resources Conservation Service Technical Note no. 190–32, 9 p., accessed March 24, 2025, at https://meridian.allenpress.com/jfwm/article-supplement/204244/pdf/fwma-08-01-11_s03.
Natural Resources Conservation Service, 2015, Review of the Lawson aerator for brush management: Boise, Idaho, U.S. Department of Agriculture Natural Resources Conservation Service Plant Materials Technical Note no. 64, 11 p., accessed August 25, 2024, at https://www.nrcs.usda.gov/plantmaterials/idpmctn12625.pdf.
Neary, D.G., and Leonard, J.M., 2020, Effects of fire on grassland soils and water—A review, chap 3 of Kindomihou, V.M., ed., Grasses and grassland aspects: IntechOpen, 22 p., accessed August 25, 2024, at https://doi.org/10.5772/intechopen.77919.
Olson, R.A., and Whitson, T.D., 2002, Restoring structure in late-successional sagebrush communities by thinning with tebuthiuron: Restoration Ecology, v. 10, no. 1, p. 146–155, accessed August 25, 2024, at https://doi.org/10.1046/j.1526-100X.2002.10116.x.
Palmquist, K.A., Schlaepfer, D.R., Renne, R.R., Torbit, S.C., Doherty, K.E., Remington, T.E., Watson, G., Bradford, J.B., and Lauenroth, W.K., 2021, Divergent climate change effects on widespread dryland plant communities driven by climatic and ecohydrological gradients: Global Change Biology, v. 27, no. 20, p. 5169–5185, accessed August 25, 2024, at https://doi.org/10.1111/gcb.15776.
Pastick, N.J., Dahal, D., Wylie, B.K., Parajuli, S., Boyte, S.P., and Wu, Z., 2020, Characterizing land surface phenology and exotic annual grasses in dryland ecosystems using Landsat and Sentinel-2 data in harmony: Remote Sensing, v. 12, no. 4, art. 725, 17 p., accessed March 25, 2025, at https://doi.org/10.3390/rs12040725.
Pastick, N.J., Wylie, B.K., Rigge, M.B., Dahal, D., Boyte, S.P., Jones, M.O., Allred, B.W., Parajuli, S., and Wu, Z., 2021a, Historic and future trends in exotic annual grass (%) cover in the Western US (1985 to 2019 and 2025 to 2040): U.S. Geological Survey data release, accessed August 25, 2024, at https://doi.org/10.5066/P9Z85VET.
Pastick, N.J., Wylie, B.K., Rigge, M.B., Dahal, D., Boyte, S.P., Jones, M.O., Allred, B.W., Parajuli, S., and Wu, Z., 2021b, Rapid monitoring of the abundance and spread of exotic annual grasses in the Western United States using remote sensing and machine learning: AGU Advances, v. 2, no. 2, art. e2020AV000298, 22 p., accessed August 25, 2024, at https://doi.org/10.1029/2020AV000298.
Pausas, J.G., Keeley, J.E., and Schwilk, D.W., 2017, Flammability as an ecological and evolutionary driver: Journal of Ecology, v. 105, no. 2, p. 289–297, accessed August 25, 2024, at https://doi.org/10.1111/1365-2745.12691.
Pellant, M., Shaver, P.L., Pyke, D.A., Herrick, J.E., Lepak, N., Riegel, G., Kachergis, E., Newingham, B.A., Toledo, D., and Busby, F.E., 2020, Interpreting indicators of rangeland health, version 5: Denver, Colo., Bureau of Land Management National Operations Center, Technical Reference 1734–6, 202 p., accessed August 25, 2024, at https://www.blm.gov/documents/national-office/blm-library/technical-reference/interpreting-indicators-rangeland-health-0.
Pilliod, D.S., Welty, J.L., and Arkle, R.S., 2017, Refining the cheatgrass-fire cycle in the Great Basin—Precipitation timing and fine fuel composition predict wildfire trends: Ecology and Evolution, v. 7, no. 19, p. 8126–8151, accessed August 25, 2024, at https://doi.org/10.1002/ece3.3414.
Poor, E.E., Kleist, N.J., Bencin, H.L., Foster, A.C., and Carter, S.K., 2021, Annotated bibliography of scientific research on Ventenata dubia published from 2010 to 2020: U.S. Geological Survey Open-File Report 2021–1031, 26 p., accessed August 25, 2024, at https://doi.org/10.3133/ofr20211031.
Porensky, L.M., and Blumenthal, D.M., 2016, Historical wildfires do not promote cheatgrass invasion in a western Great Plains steppe: Biological invasions, v. 18, p. 3333–3349, accessed August 25, 2024, at https://doi.org/10.1007/s10530-016-1225-z.
Porensky, L.M., Derner, J.D., and Pellatz, D.W., 2018, Plant community responses to historical wildfire in a shrubland–grassland ecotone reveal hybrid disturbance response: Ecosphere, v. 9, no. 8, art. e02363, 17 p., accessed August 25, 2024, at https://doi.org/10.1002/ecs2.2363.
Prevéy, J.S., Jarnevich, C.S., Pearse, I.S., Munson, S.M., Stevens, J.T., Barrett, K.J., Coop, J.D., Day, M.A., Firmage, D., Fornwalt, P.J., Haynes, K.M., Johnston, J.D., Kerns, B.K., Krawchuk, M.A., Miller, B.A., Nietupski, T.C., Roque, J., Springer, J.D., Stevens-Rumann,C.S., Stoddard, M.T., and Tortorelli, C.M., 2024, Non-native plant invasion after fire in Western USA varies by functional type and with climate: Biological Invasions, v. 26, p. 1157–1179, accessed August 25, 2024, at https://doi.org/10.1007/s10530-023-03235-9.
Pyke, D.A., 2011, Restoring and rehabilitating sagebrush habitats, in Knick, S.T., and Connelly, J.W., eds., Greater sage-grouse—Ecology and conservation of a landscape species and its habitats: University of California Press, p. 531–548, accessed August 25, 2024, at https://pubs.usgs.gov/publication/70004639.
Rau, B.M., Blank, R.R., Chambers, J.C., and Johnson, D.W., 2007, Prescribed fire in a Great Basin sagebrush ecosystem—Dynamics of soil extractable nitrogen and phosphorus: Journal of Arid Environments, v. 71, no. 4, p. 362–375, accessed August 25, 2024, at https://doi.org/10.1016/j.jaridenv.2007.05.006.
Reaser, J.K., Burgiel, S.W., Kirkey, J., Brantley, K.A., Veatch, S.D., and Burgos-RodrigueaJ., 2020, The early detection of and rapid response (EDRR) to invasive species—A conceptual framework and Federal capacities assessment: Biological Invasions, v. 22, p. 1–19, accessed August 25, 2024, at https://doi.org/10.1007/s10530-019-02156-w.
Reeves, M.C., and Mitchell, J.E., 2011, Extent of coterminous US rangelands—Quantifying implications of differing agency perspectives: Rangeland Ecology & Management, v. 64, no. 6, p. 585–597, accessed September 16, 2024, at https://doi.org/10.2111/REM-D-11-00035.1.
Reinhart, K.O., Williams, A.R., and Vermeire, L.T., 2020, Effects of defoliation, litter, and moss on Bromus arvensis in a northern mixed-grass prairie: Rangeland Ecology & Management, v. 73, no. 5, p. 607–610, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2020.06.005.
Rigge, M., Homer, C., Cleeves, L., Meyer, D.K., Bunde, B., Shi, H., Xian, G., Schell, S., and Bobo, M., 2020, Quantifying Western U.S. Rangelands as fractional components with multi-resolution remote sensing and in situ data: Remote Sensing, v. 12, no. 3, art. 412, 26 p., accessed August 25, 2024, at https://doi.org/10.3390/rs12030412.
Rigge, M., Homer, C., Shi, H., and Meyer, D.K., 2019, Validating a Landsat time-series of fractional component cover across Western U.S. rangelands: Remote Sensing, v. 11, no. 24, art. 3009, 16 p., accessed August 25, 2024, at https://doi.org/10.3390/rs11243009 .
Rigge, M., Homer, C., Shi, H., Meyer, D., Bunde, B., Granneman, B., Postma, K., Danielson, P., Case, A., and Xian, G., 2021, Rangeland fractional components across the Western United States from 1985 to 2018: Remote Sensing, v. 13, no. 4, art. 813, 24 p., accessed August 25, 2024, at https://doi.org/10.3390/rs13040813.
Rinella, M.J., Mangold, J.M., Espeland, E.K., Sheley, R.J., and Jacobs, J.S., 2012, Long-term population dynamics of seeded plants in invaded grasslands: Ecological Applications, v. 22, no. 4, p. 1320–1329, accessed August 25, 2024, at https://doi.org/10.1890/11-1103.1.
Ross, M.R., Castle, S.C., and Barger, N.N., 2012, Effects of fuels reductions on plant communities and soils in a piñon-juniper woodland: Journal of Arid Environments, v. 79, p. 84–92, accessed August 25, 2024, at https://doi.org/10.1016/j.jaridenv.2011.11.019.
Rutherford, T.K., Maxwell, L.M., Kleist, N.J., Teige, E.C., Lehrter, R.J., Gilbert, M.A., Wood, D.J.A., Johnston, A.N., Mengelt, C., Tull, J.C., Haby, T.S., and Carter, S.K., 2023, Effects of noise from oil and gas development on ungulates and small mammals—A science synthesis to inform National Environmental Policy Act analyses: U.S. Geological Survey Scientific Investigations Report 2023–5114, 44 p., accessed July 22, 2024, at https://doi.org/10.3133/sir20235114.
Ryan, C.M., Cerveny, L.K., Robinson, T.L., and Blahna, D.J., 2018, Implementing the 2012 forest planning rule—Best available scientific information in forest planning assessments: Forest Science, v. 64, no. 2, p. 159–169, accessed August 25, 2024, at https://doi.org/10.1093/forsci/fxx004.
Saher, D.J., Shyvers, J.E., Tarbox, B.C., Van Schmidt, N.D., Heinrichs, J.A., and Aldridge, C.L., 2022, Compendium to invasive annual grass spatial products for the Western United States, January 2010–February 2021: U.S. Geological Survey Data Report 1152, 63 p., accessed August 25, 2024, at https://doi.org/10.3133/dr1152.
Sankey, J.B., Germino, M.J., Sankey, T.T., and Hoover, A.N., 2012, Fire effects on the spatial patterning of soil properties in sagebrush steppe, USA—A meta-analysis: International Journal of Wildland Fire, v. 21, no. 5, p. 545–556, accessed August 25, 2024, at https://doi.org/10.1071/WF11092.
Santín, C., Doerr, S.H., Kane, E.S., Masiello, C.A., Ohlson, M., de la Rosa, J.M., Preston, C.M., and Dittmar, T., 2016, Towards a global assessment of pyrogenic carbon from vegetation fires: Global Change Biology, v. 22, no. 1, p. 76–91, accessed August 25, 2024, at https://doi.org/10.1111/gcb.12985.
Savage, S., and Slyder, J., 2022, Evaluation of fractional vegetation cover products: Denver, Colo., Bureau of Land Management National Operations Center Technical Note 456, 44 p., accessed August 25, 2024, at https://www.blm.gov/noc/blm-library/technical-note/evaluation-fractional-vegetation-cover-products.
Seavy, N.E., and Howell, C.A., 2010, How can we improve information delivery to support conservation and restoration decisions?: Biodiversity and Conservation, v. 19, no. 5, p. 1261–1267, accessed August 25, 2024, at https://doi.org/10.1007/s10531-009-9752-x .
Seipel, T., Rew, L.J., Taylor, K.T., Maxwell, B.D., and Lehnhoff, E.A., 2018, Disturbance type influences plant community resilience and resistance to Bromus tectorum invasion in the sagebrush steppe: Applied Vegetation Science, v. 21, no. 3, p. 385–394, accessed August 25, 2024, at https://doi.org/10.1111/avsc.12370.
Sheley, R.L., Jacobs, J.S., and Lucas, D.E., 2001, Revegetating spotted knapweed infested rangeland in a single entry: Journal of Range Management, v. 54, no. 2, p. 144–151, accessed August 25, 2024, at https://doi.org/10.2307/4003175.
Shyvers, J.E., Tarbox, B.C., Van Schmidt, N.D., Saher, D.J., Heinrichs, J.A., and Aldridge, C.L., 2022, Database of invasive annual grass spatial products for the Western United States January 2010 to February 2021: US Geological Survey data release, accessed August 25, 2024, at https://doi.org/10.5066/P9VW97AO.
Smith, J.T., Allred, B.W., Boyd, C.S., Davies, K.W., Jones, M.O., Kleinhesselink, A.R., Maestas, J.D., Morford, S.L., and Naugle, D.E., 2021, The elevational ascent and spread of exotic annual grass dominance in the Great Basin, USA: Diversity and Distributions, v. 28, no. 1, p. 83–96, accessed August 25, 2024, at https://doi.org/10.1111/ddi.13440.
Smith, J.T., Allred, B.W., Boyd, C.S., Davies, K.W., Kleinhesselink, A.R., Morford, S.L., and Naugle, D.E., 2023, Fire needs annual grasses more than annual grasses need fire: Biological Conservation, v. 286, art. 110299, 9 p., accessed August 25, 2024, at https://doi.org/10.1016/j.biocon.2023.110299.
Smith, D.C., Meyer, S.E., and Anderson, V.J., 2008, Factors affecting Bromus tectorum seed bank carryover in western Utah: Rangeland Ecology & Management, v. 61, no. 4, p. 430–436, accessed August 25, 2024, at https://doi.org/10.2111/07-035.1.
Sovacool, B.K., Axsen, J., and Sorrell, S., 2018, Promoting novelty, rigor, and style in energy social science—Towards codes of practice for appropriate methods and research design: Energy Research & Social Science, v. 45, p. 12–42, accessed August 25, 2024, at https://doi.org/10.1016/j.erss.2018.07.007.
Sturges, D.L., 1993, Soil-water and vegetation dynamics through 20 years after big sagebrush control: Journal of Range Management, v. 46, no. 2, p. 161–169, accessed August 25, 2024, at https://doi.org/10.2307/4002275.
Symstad, A.J., Buhl, D.A., and Swanson, D.J., 2021, Fire controls annual bromes in northern Great Plains grasslands—Up to a point: Rangeland Ecology & Management, v. 75, p. 17–28, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2020.11.003.
Symstad, A.J., and Leis, S.A., 2017, Woody encroachment in northern Great Plains Grasslands—Perceptions, actions, and needs: Natural Areas Journal, v. 37, no. 1, p. 118–127, accessed August 25, 2024, at https://doi.org/10.3375/043.037.0114.
Symstad, A.J., Gitzen, R.A., Wienk, C.L., Bynum, M.R., Swanson, D.J., Thorstenson, A.J., and Paintner-Green, K.J., 2012, Plant community composition and structure monitoring protocol for the northern Great Plains I&M Network—Standard operating procedures—Version 1.01: National Park Service Natural Resource Report NPS/NGPN/NRR—2012/489, 276 p., accessed August 25, 2024, at https://irma.nps.gov/DataStore/DownloadFile/447562.
Tarbox, B.C., Van Schmidt, N.D., Shyvers, J.E., Saher, D.J., Heinrichs, J.A., and Aldridge, C.L., 2022, Bridging the gap between spatial modeling and management of invasive annual grasses in the imperiled sagebrush biome: Rangeland Ecology & Management, v. 82, p. 104–115, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2022.01.006.
Taylor, K., Brummer, T., Rew, L.J., Lavin, M., and Maxwell, B.D., 2014, Bromus tectorum response to fire varies with climate conditions: Ecosystems, v. 17, p. 960–973, accessed August 25, 2024, at https://doi.org/10.1007/s10021-014-9771-7.
Thill, D.C., Schirman, R.D., and Appleby, A.P., 1979, Influence of soil moisture, temperature, and compaction on the germination and emergence of downy brome (Bromus tectorum): Weed Science, v. 27, no. 6, p. 625–630, accessed August 25, 2024, at https://doi.org/10.1017/S0043174500046014.
Thomas, S.M., and Moloney, K.A., 2015, Combining the effects of surrounding land-use and propagule pressure to predict the distribution of an invasive plant: Biological Invasions, v. 17, p. 477–495, accessed August 25, 2024, at https://doi.org/10.1007/s10530-014-0745-7.
Toevs, G.R., Karl, J.W., Taylor, J.J., Spurrier, C.S., Karl, M., Bobo, M.R., and Herrick, J.E., 2011, Consistent indicators and methods and a scalable sample design to meet assessment, inventory, and monitoring information needs across scales: Rangelands, v. 33, no. 4, p. 14–20, accessed August 25, 2024, at https://doi.org/10.2111/1551-501X-33.4.14.
Tortorelli, C., Krawchuk, M., and Kerns, B., 2020, Expanding the invasion footprint—Ventenata dubia and relationships to wildfire, environment, and plant communities in the Blue Mountains of the Inland Northwest, USA: Applied Vegetation Science, v. 23, no. 4, p. 562–574, accessed August 25, 2024, at https://doi.org/10.1111/avsc.12511.
Tulganyam, S., and Carr, C.A., 2022, Short-term vegetation response to bulldozed fire lines in northern Great Plains grasslands: Fire, v. 5, no. 3, art. 79, 10 p., accessed August 25, 2024, at https://doi.org/10.3390/fire5030079.
Van Schmidt, N.D., Shyvers, J.E., Saher, D.J., Tarbox, B.C., Heinrichs, J.A., and Aldridge, C.L., 2022, A user guide to selecting invasive annual grass spatial products for the Western United States: U.S. Geological Survey Fact Sheet 2022–3001, 6 p., accessed August 25, 2024, at https://pubs.usgs.gov/fs/2022/3001/fs20223001.pdf.
Vermeire, L.T., Crowder, J.L., and Wester, D.B., 2011, Plant community and soil environment response to summer fire in the northern Great Plains: Rangeland Ecology & Management, v. 64, no. 1, p. 37–46, accessed August 25, 2024, at https://doi.org/10.2111/REM-D-10-00049.1.
Vermeire, L.T., Crowder, J.L., and Wester, D.B., 2014, Semiarid rangeland is resilient to summer fire and postfire grazing utilization: Rangeland Ecology & Management, v. 67, no. 1, p. 52–60, accessed August 25, 2024, at https://doi.org/10.2111/REM-D-13-00007.1.
Vermeire, L.T., and Rinella, M.J., 2017, Fire alters emergence of invasive plant species from soil surface-deposited seeds: Weed Science, v. 57, no. 3, p. 304–310, accessed August 25, 2024, at https://doi.org/10.1614/WS-08-170.1.
Vermeire, L.T., Rinella, M.J., and Strong, D.J., 2021, Individual and combined effects of fall fire and growth-regulator herbicide on annual bromes: Rangeland Ecology & Management, v. 76, p. 129–138, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2021.02.009.
Vermeire, L.T., and Russell, M.L., 2018, Seasonal timing of fire alters biomass and species composition of northern mixed prairie: Rangeland Ecology & Management, v. 71, no. 6, p. 714–720, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2018.06.005.
Vermeire, L.T., Strong, D.J., Gates, E.A., Marlow, C.B., and Waterman, R.C., 2020, Can mowing substitute for fire in semiarid grassland?: Rangeland Ecology & Management, v. 73, no. 1, p. 97–103, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2019.08.006.
Vermeire, L.T., Waterman, R.C., Reinhart, K.O., and Rinella, M.J., 2023, Grazing intensity and seasonality manipulate invasive annual grasses and native vegetation: Rangeland Ecology & Management, v. 90, p. 308–313, accessed August 25, 2024, at https://doi.org/10.1016/j.rama.2023.04.001.
Wade, N.M., Vermeire, L.T., and Scasta, J.D., 2024, Prescribed fire and exotic plant responses in phreatophytic Sarcobatus vermiculatus rangelands with contrasting degradation: Global Ecology and Conservation, v. 52, art. e02955, 16 p., accessed August 25, 2024, at https://doi.org/10.1016/j.gecco.2024.e02955.
Wäldchen, J., and Mäder, P., 2018, Plant species identification using computer vision techniques—A systematic literature review: Archives of Computational Methods in Engineering, v. 25, no. 2, p. 507–543, accessed August 25, 2024, at https://doi.org/10.1007/s11831-016-9206-z.
Whisenant, S.G., and Uresk, D.W., 1990, Spring burning Japanese brome in a western wheatgrass community: Rangeland Ecology & Management, v. 43, no. 3, p. 205–208, accessed August 25, 2024, at https://doi.org/10.2307/3898673.
Wicks, G.A., 1997, Survival of downy brome (Bromus tectorum) seed in four environments: Weed Science, v. 45, no. 2, p. 225–228, accessed August 25, 2024, at https://doi.org/10.1017/S0043174500092754.
Williams, C.J., Johnson, J.C., Pierson, F.B., Burleson, C.S., Polyakov, V.O., Kormos, P.R., and Nouwakpo, S.K., 2020, Long-term effectiveness of tree removal to re-establish sagebrush steppe vegetation and associated spatial patterns in surface conditions and soil hydrologic properties: Water, v. 12, no. 8, art. 2213, 35 p., accessed August 25, 2024, at https://doi.org/10.3390/w12082213.
Williams, D.A., Jarnevich, C.S., Engelstad, P., Henderson, G., Shadwell, K., Pearse, I.S., and Prevey, J.S., 2023, Potential distribution of Japanese brome (Bromus japonicus) across the contiguous United States (October 2023): U.S. Geological Survey data release, accessed September 16, 2024, at https://doi.org/10.5066/P9HSW0AI.
Wozniak, S.S., Strand, E.K., Johnson, T.R., Hulet, A., Roundy, B.A., and Young, K., 2020, Treatment longevity and changes in surface fuel loads after pinyon-juniper mastication: Ecosphere, v. 11, no. 8, art. e03226, 18 p., accessed August 25, 2024, at https://doi.org/10.1002/ecs2.3226.
Wyborn, C., Louder, E., Harrison, J., Montambault, J., Montana, J., Ryan, M., Bednarek, A., Nesshöver, C., Pullin, A., Reed, M., Dellecker, E., Kramer, J., Boyd, J., Dellecker, A., Hutton, J., 2018, Understanding the impacts of research synthesis: Environmental Science & Policy, v. 86, p. 72–84, accessed August 25, 2024, at https://doi.org/10.1016/j.envsci.2018.04.013.
Zachman, D., 2003, Mechanical treatments to meet resource needs: Bureau of Land Management Resource Notes no. 67, 2 p., accessed August 25, 2024, at https://www.blm.gov/sites/default/files/documents/files/Library_BLMResourceNote67.pdf
Zhang, J., Okin, G.S., and Zhou, B., 2019, Assimilating optical satellite remote sensing images and field data to predict surface indicators in the Western US—Assessing error in satellite predictions based on large geographical datasets with the use of machine learning: Remote Sensing of Environment, v. 233, art. 111382, 11 p., accessed August 25, 2024, at https://doi.org/10.1016/j.rse.2019.111382.
Zhou, B, Okin, G.S., and Zhang, J., 2020, Leveraging Google Earth Engine (GEE) and machine learning algorithms to incorporate in situ measurement from different times for rangelands monitoring: Remote Sensing of Environment, v. 236, art. 111521, 19 p., accessed August 25, 2024, at https://doi.org/10.1016/j.rse.2019.111521.
Zouhar, K., 2021, Fire regimes of plains grassland and prairie ecosystems: U.S. Department of Agriculture Forest Service web page, accessed August 25, 2024, at www.fs.usda.gov/database/feis/fire_regimes/PlainsGrass_Prairie/all.html.
Glossary
- aerator
A single or double drum with offset blades oriented in a spiral pattern pulled behind a tractor that is used to reduce shrub cover and increase grasses and forbs. Also known as a spiral blade chopper (Natural Resources Conservation Service, 2015).
- broadcast burn
A type of prescribed fire applied under specific weather and fuel conditions that allow existing fuels to carry the fire through a targeted area to achieve management objectives.
- chain
Heavy metal links dragged in a U- or J-shaped pattern between two tractors traveling parallel to each other to knockdown brush and small trees (McKenzie and others, 1984).
- cumulative effects
“effects on the environment that result from the incremental effects of the action when added to the effects of other past, present, and reasonably foreseeable actions regardless of what agency (Federal or non-Federal) or person undertakes such other actions. Cumulative effects can result from actions with individually minor but collectively significant effects taking place over a period of time.” (40 CFR 1508.1(i))
- direct effects
Effects “which are caused by the action and occur at the same time and place.” (40 CFR 1508.1(i))
- disking
A mechanical treatment in which a farming implement with vertical metal disks is towed behind a tractor to dig up the top layer of soil (Natural Resources Conservation Service, 2005).
- excavator
A vehicle equipped with a boom and bucket designed to dig, lift, and move heavy materials, particularly soil and vegetation.
- furrow
A farming implement with V-shaped metal scoops that is towed behind a tractor to dig up the top layer of soil and create small linear valleys where water can pool (Lacey and others, 1981).
- hand-pulling
Manual removal of vegetation using chainsaws or other cutting methods by workers on foot.
- hoe
A farming implement with metal scoops that is towed behind a tractor to dig up the top layer of soil (Griepentrog and others, 2007).
- harrow
A spike-toothed farming implement that is pulled behind a tractor to grab and rip vegetation and soil (Dahlgren and others, 2006).
- indirect effects
Effects “which are caused by the action and are later in time or farther removed in distance, but are still reasonably foreseeable.” (40 CFR 1508.1(i))
- masticator
Mechanical attachment to tractors or excavators with metal teeth and blades used to mulch trees and shrubs (Wozniak and others, 2020).
- mowing
Removal of vegetation at a specified height above ground using a blade and machinery
- noxious weed
An undesirable plant that is regulated by law.
- prescribed fire
“A fire ignited under known conditions of fuel, weather, and topography to achieve specified objectives.” (Agee, 1993, p. 418)
- resilience
The ability of an area to regain its fundamental structure, process, and function despite stressors, disturbances, or invasive species (Chambers, Miller, and others, 2014).
- resistance
The ability of an area to retain its fundamental structure, process, and function despite stressors, disturbances, or invasive species (Chambers, Miller, and others, 2014).
- rollerchopper
A cylindrical roller equipped with several full-length blades that is towed to shear small trees and shrubs off at the base and chop up the stems and slash (Zachman, 2003).
- skid steer
A vehicle equipped with a bucket designed to dig, lift, and move heavy materials, particularly soil and vegetation.
- wildfire
“A fire, naturally caused or caused by humans, that is not meeting land management objectives.” (Agee, 1993, p. 419).
Abbreviations
AIM
Assessment, Inventory, and Monitoring
BLM
Bureau of Land Management
CFR
Code of Federal Regulations
IAG
invasive annual grass
LandCART
Landscape Cover Analysis and Reporting Tools
MaxEnt
maximum entropy modeling
NEPA
National Environmental Policy Act
RAP
Rangeland Analysis Platform
RCMAP
Rangeland Condition Monitoring Assessment and Projection
ssp.
subspecies
spp.
several species of
U.S.C.
United States Code
USGS
U.S. Geological Survey
Species Names
Common name
Scientific name
antelope bitterbrush
Purshia tridentata (Pursh) DC.
arrowleaf balsamroot
Balsamorhiza sagittata (Pursh) Nutt.
big sagebrush subspecies
Artemisia tridentata ssp.
black greasewood
Sarcobatus vermiculatus (Hook.) Torr.
blue grama
Bouteloua gracilis (Kunth) Lag. ex Griffiths
bluebunch wheatgrass
Pseudoroegneria spicata (Pursh) Á. Löve
brome
Bromus spp.
cactus species
Opuntia spp.
cheatgrass
Bromus tectorum L.
cinquefoil species
Potentilla spp.
common chokecherry
Prunus virginiana L.
common snowberry
Symphoricarpos albus (L.) S.F. Blake
cream sacs
Castilleja rubicundula (Jeps.) T.I. Chuang & Heckard
curl-leaf mountain-mahogany
Cercocarpus ledifolius Nutt.
dotted blazing star
Liatris punctata Hook.
fescue species
Festuca spp.
field brome
Bromus arvensis L.
fleabane species
Erigeron spp.
green needlegrass
Nassella viridula (Trin.) Barkworth
Idaho fescue
Festuca idahoensis Elmer
Indian paintbrush species
Castilleja spp.
Indian ricegrass
Achnatherum hymenoides (Roem. & Schult.) Barkworth
Japanese brome
Bromus japonicus Thunb. ex Murray
Juniper species
Juniperus spp. L.
Kentucky bluegrass
Poa pratensis L.
low sagebrush
Artemisia arbuscula Nutt.
medusahead
Taeniatherum caput-medusae (L.) Nevski
milkvetch species
Astragalus spp.
mountain big sagebrush
Artemisia tridentata ssp. vaseyana (Rydb.) Beetle
needle-and-thread
Hesperostipa comata (Trin. & Rupr.) Barkworth
needlegrass species
Achnatherum spp.
phlox species
Phlox spp.
pine species
Pinus spp. L.
purple prairie clover
Dalea purpurea Vent.
red brome
Bromus rubens L.
rough fescue
Festuca campestris Rydb.
rubber rabbitbrush
Ericameria nauseosa (Pall. ex Pursh) G.L. Nesom & Baird
sage-grouse
Centrocercus spp.
saltbush species
Atriplex spp.
Sandberg bluegrass
Poa secunda J. Presl
sandwort species
Arenaria spp.
scarlet globemallow
Sphaeralcea coccinea (Nutt.) Rydb.
shortbristle needle and thread
Hesperostipa curtiseta (Hitchc.) Barkworth
silver sagebrush
Artemisia cana Pursh
smooth brome
Bromus inermis Leyss.
squirreltail
Elymus elymoides (Raf.) Swezey
subalpine big sagebrush
Artemisia spiciformis Osterh.
sumac species
Rhus spp. L.
thickspike wheatgrass
Elymus lanceolatus ssp. lanceolatus (Scribn. & J.G. Sm.) Gould
three-tip sagebrush
Artemisia tripartita Rydb.
ventenata
Ventenata dubia (Leers) Coss.
western wheatgrass
Pascopyrum smithii (Rydb.) Á. Löve
Wyoming big sagebrush
Artemisia tridentata ssp. wyomingensis Beetle & Young
Publishing support provided by the Science Publishing Network, Denver and Lafayette Publishing Service Centers
For more information concerning the research in this report, contact the
Center Director, USGS Northern Rocky Mountain Science Center
2327 University Way, Suite 2
Bozeman, MT 59715
(406) 994-7544
Or visit the Northern Rocky Mountain Science Center website at
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
Johnston, A.N., Wood, D.J.A., Ebenhoch, K.G., Rutherford, T.K., Maxwell, L.M., and Carter, S.K., 2025, Potential risks of vegetation treatments to introduce and increase invasive annual grasses in rangelands of Montana, North Dakota, and South Dakota—A science synthesis to inform National Environmental Policy Act analyses: U.S. Geological Survey Scientific Investigations Report 2025–5058, 36 p., https://doi.org/10.3133/sir20255058.
ISSN: 2328-0328 (online)
Study Area
| Publication type | Report |
|---|---|
| Publication Subtype | USGS Numbered Series |
| Title | Potential risks of vegetation treatments to introduce and increase invasive annual grasses in rangelands of Montana, North Dakota, and South Dakota—A science synthesis to inform National Environmental Policy Act analyses |
| Series title | Scientific Investigations Report |
| Series number | 2025-5058 |
| DOI | 10.3133/sir20255058 |
| Publication Date | September 29, 2025 |
| Year Published | 2025 |
| Language | English |
| Publisher | U.S. Geological Survey |
| Publisher location | Reston VA |
| Contributing office(s) | Northern Rocky Mountain Science Center |
| Description | ix, 36 p. |
| Country | United States |
| State | Montana, North Dakota, South Dakota |
| Online Only (Y/N) | Y |