BENTHIC COMMUNITIES IN LONG ISLAND SOUND
The ability to interpret and predict changes in the LIS ecosystem by scientists and managers in relation to continuing degradation of the system, or in response to efforts to stem human impacts, rests on the accuracy and completeness of the information available on the system and the ecological models based on this information. With respect to soft-sediment communities in LIS, much work has been done to characterize their structure and understand their dynamics. In this portion of the project, available information on benthic communities in the Sound was assessed and in some case re-analyzed in order to develop an integrated view of benthic community structure, reassess models concerning community dynamics in light of this review and to identify potentially important gaps in our understanding of these communities.
The review was initiated by identifying studies which addressed benthic community structure in the off-shore waters of Long Island Sound. In particular, the focus was on survey studies which provided information on community structure within and among different areas of the Sound, thus allowing for an assessment of the spatial characteristics and variability of community structure. Comparing information from different studies conducted at different times over the last 40 years which used different methods is difficult to say the least and caution must be applied in order to not overextend the analysis and the interpretations based on these data. The approach here was to identify information that would enhance our general understanding of the types and distribution of the Sound's benthic communities (not trying to compare, for example, absolute species densities among studies), and which would provide further insights as to the dynamics of the communities. Therefore, comparisons among studies were based on species composition and identification of dominant species based on general abundance trends. More detailed analyses were carried out on data collected by Pellegrino and Hubbard (1983). Their study collected benthic samples with a relatively high spatial resolution and allowed for more detailed analyses of community composition, distribution and relationships with various habitat characteristics. The specific methods used in this set of analyses are given below. Based on the overall analyses, a model of benthic community structure and dynamics was developed relative to the information available for the Sound.
Soft Sediment Communities in Long Island Sound - A Historical Overview
There have been numerous studies which have focused on organisms which inhabit the soft-sediment habitats of the Sound (Lewis and Coffin, 1985). However, only a small subset of these have focused on community level characteristics of the soft-sediment fauna (Table 3) or on selected aspects of infaunal ecology such as population ecology, organism / sediment interactions, specific types of disturbances and syntheses of previous work (Table 4).
Our knowledge of the spatial and temporal characteristics of LIS infaunal communities, on a Sound-wide basis, is derived primarily from the studies listed in Table 3. Soft-sediment studies in the Sound were initiated in 1953 by Sanders (1956), but the bulk of the available information was collected during the 1970's and early 1980's. Since then few community level studies have been conducted . There have been only two sound-wide surveys. Reid et al. (1979) sampled 142 sites covering most of the deep water habitats of the Sound, while Pellegrino & Hubbard (1982) samples 413 sites which were all located in Connecticut waters (Figure 8). Other surveys focused on two specific areas of the Sound (Figure 8): the central basin (Sanders 1956, McCall 1978, Rhoads and Germano 1982) and Fishers Island Sound (Franz 1976, Swanson 1977, Biernbaum 1979). There were also studies which focused on the dynamics of communities in the central basin of the Sound with respect to responses to disturbance and organism / sediment interactions (McCall 1977, Rhoads et al. 1977).
Several key points should be made with regard to this information base. Most of the work on soft-sediment communities in the Sound has been conducted in the central basin; our knowledge of community structure in the western and eastern basins is limited. Therefore, resolving important spatial characteristics of soft-sediment communities in the Sound will be limited by this uneven spatial focus of past studies. Furthermore, the temporal resolution of the surveys, both those conducted on a sound-wide basis and those in specific areas, was coarse. Most sites were only sampled one to three times over the study period (Table 3). Therefore, we do not possess a detailed view of important seasonal changes that can occur in community structure. Finally, almost all of these studies used a 1 mm sieve to collect infaunal organisms, therefore missing a potentially important size-fraction of the macrobenthos comprised of small organisms and the juvenile stages of larger fauna.
These studies have all made important contributions to our knowledge of benthic community structure and dynamics in LIS and to marine benthic ecology in general. The work of Sanders (1956) laid the foundation of our knowledge and from this work subsequent studies either provided further information on community structure and/or dynamics. By considering and comparing some of the main findings of these studies we can assess our current understanding of the LIS benthos and how this may relate to management issues. I present below a review of the main findings of these studies in chronological order, followed by several analyses which address relationships between habitat and community characteristics and then assess these results relative to present models of benthic community structure and dynamics.
The Seminal Work of Howard Sanders
Sanders (1956) initiated studies on several basic aspects of infaunal organisms in the Sound, including relationships between sediment and other habitat characteristics and community structure, the feeding modes of infauna, their life histories, secondary production and the classification of community types. Indeed, this work stands as the seminal work in soft-sediment ecology in North America, developing many of the major themes which subsequent benthic ecologists have investigated throughout the world. His work was part of the first wave of detailed and integrated studies of the physical, chemical and biological characteristics and dynamics of Long Island Sound by researchers at the Bingham Oceanographic Laboratory at Yale University (Riley 1956).
Sanders (1956) conducted his studies between the summer of 1953 and the fall of 1954 at 8 sites in the central portion of the Sound, roughly between Stratford Shoal and the Thimble Islands, at depths of 6 to 29 m (Figure 8). Samples were taken with a modified anchor dredge and fauna were collected on 1 and 2 mm sieves. This method is subject to error with respect to quantifying infaunal abundances as the area sampled was determined by dividing the volume of the material by the estimated depth of penetration by the dredge. Nonetheless, Sanders' work provided an in-depth characterization of this portion of the Sound and a fairly detailed framework which later studies have built on.
The stations Sanders (1956) sampled were comprised of varied bottom types. Variation in habitat characteristics both within and among the sampling areas was recognized by Sanders as a potentially important determinant of Sound-wide trends in benthic community structure. He also noted that while there were predominant sediment grain-size classes at each station, the sediments were often heterogenous. For example, even areas which were predominantly comprised of coarse sands had significant amounts of clay and silt. Also noted were the potential effects of other habitat characteristics on infaunal community characteristics, such as the presence of shell hash. Based on comparisons between habitat and community characteristics, Sanders (1956) found that community structure and the abundance of organisms was related to the percent silt-clay composition of the sediments, with intermediate levels (13 - 35 %) being correlated with the highest biomass values.
Comparisons to previous studies done elsewhere at the time established LIS as being highly productive with respect to benthic secondary production and biomass. Sanders argued that this may be related to the Sound's hydrodynamics and nutrient additions from the watershed which resulted in high levels of primary production being readily available to the benthos.
During the course of his study, Sanders (1956) identified 119 species of benthic fauna. Among the eight stations the dominant species included the polychaetes Nephtys incisa and Cistenoides gouldii, two species of amphipods in the genus Ampelisca, and the bivalves Yoldia limatula, Nucula proxima, Pandora gouldiana, Macoma tenta, Lyonsia hyalina and Mulinia lateralis. These species were distributed unevenly among the stations, in terms of abundance. Production to biomass was estimated to range between 5.0, for short-lived organisms, and 2.1, for larger infauna. Based on his analyses, Sanders (1956) was able to recognize one community type that was consistent among four of the eight sites he sampled: the Nephtys incisa - Yoldia limatula community. This community was found at 4 - 30 m depths in sediments of > 25 % silt-clay content where temperature and salinity ranged between 5E and 22EC and 24.7 - 29.2 l, respectively. These four stations were located in the central basin of the sound (Figure 8).
The 1970s: Surveys and Dynamics
After Sanders' (1956) study, work on benthic communities in LIS during the 1960s was limited. Carey (1962) studied the population ecology of two abundant species in the central basin, the polychaete worm Nephtys incisa and the lamellibranch bivalve Nucula proxima. The benthic epifauna of Long Island Sound was surveyed by Richards and Riley (1967). The 1970's, however, saw an upsurge in studies not only on benthic communities but also on many geological, chemical, oceanographic and environmental (i.e. human impacts) aspects of the Sound (Squires 1993). Studies of benthic communities were conducted primarily in two geographical areas. A large body of work that focused on specific aspects of benthic dynamics was generated by researchers at Yale University working in the central basin of LIS.. This included work on the dynamics of soft-sediment communities and organisms, particularly responses to disturbance and successional dynamics (Fisher and McCall 1973; McCall 1975, 1977, 1978; Rhoads et al. 1978) and physical, chemical and microbiological aspects of sediment / organism relationships (e.g. Aller 1978, 1980 1982; Rhoads et al. 1977, 1978; Rhoads and Boyer 1982; Yingst and Rhoads 1978). Much work was also done to characterize benthic communities in relation to specific impacts such as dredge spoil disposal at the New Haven dredge disposal site (e.g. Rhoads et al. 1977) and dredging and industrial operations in New Haven harbor (e.g. Rhoads and Michael 1974). It should be noted that a significant number of studies have been conducted to assess the impacts of dredging and dredge spoil disposal in Long Island Sound, providing important information on benthic communities in these areas and the potential impacts of these activities (Fredette et al. 1993, and references therein). Of the studies noted above, those by McCall (1977, 1978) will be considered relative to assessing the distribution and characteristics of infaunal communities in the Sound.
The other geographical area in which benthic communities were studied in some detail is Fishers Island Sound, from Groton to Watch Hill (Figure 8). Studies conducted by Franz (1976), Swanson (1977) and Biernbaum (1979) surveyed molluscan, polychaete and amphipod assemblages, respectively, in this area and assessed relationship to habitat characteristics. In addition to detailed studies in these two portions of the Sound, one sound-wide survey was conducted during the 1970's by Ried et al. (1979). Below, I summarize the findings of this survey and those conducted in Fisher's Island Sound by Franz (1976), Swanson (1977) and Biernbaum (1979) and in the central basin of the Sound by McCall 1977, 1978).
Reid et al's. Surveys
The first sound-wide survey of benthic communities in LIS was conducted between 1972 and 1973 by Reid at al. (1979). They sampled 142 stations three times: during the summer of 1972 all stations were sampled, and in April and September 1973, 40 to 95 stations were re-sampled. The stations were located every 3-5 km on north-south transects spaced 8.7 km apart for the length of LIS (Figure 8). In addition to characterizing infaunal communities and sediment type, synoptic measurements of temperature, salinity, nutrient concentrations, dissolved oxygen, sediment organic matter and heavy metals were also made. Ried (1979) revisited 45 of the stations once each year between 1975 and 1978.
During the course of the study, most of the species collected were annelids, mollusks and arthropods, accounting for 46%, 21% and 33% of the 248 species identified within these taxa, respectively. Species diversity, calculated for each station using the Shannon-Weaver (1949) index, H', varied spatially (Figure 9). The lowest diversities were often found at deep water stations in sediments with a high silt-clay content, although some pockets of low diversity were also noted in shallow areas. Low to moderate levels were found throughout the Sound at all depths, while the highest diversity was found at stations located in Fishers Island Sound and south of Niantic Bay. Areas of moderate to high levels of diversity were also found throughout much of the eastern basin, along several transects form nearshore to deep-water sites in the central basin, in the area of Stratford Shoal and at several nearhore stations in the western basin (Figure 9).
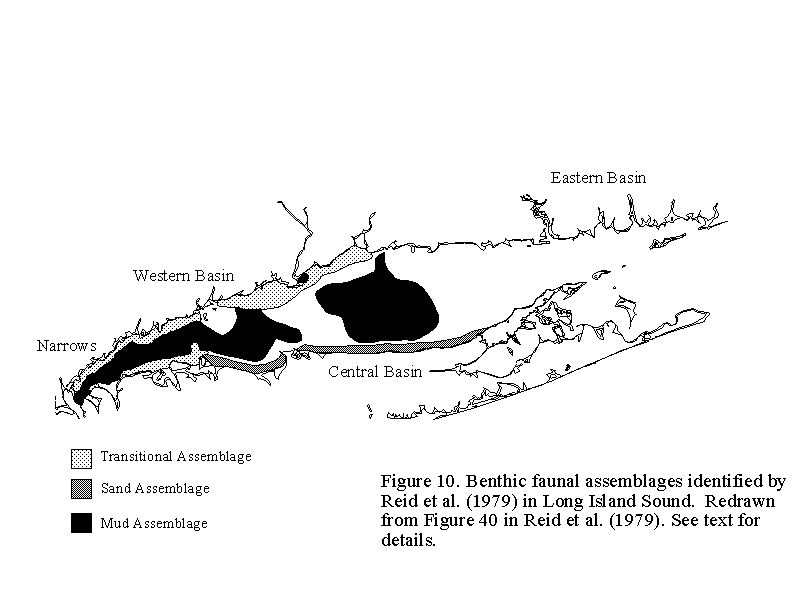
Using data collected from the samples taken during the summer of 1972, Reid et al. (1979) used cluster analysis (e.g. Ludwig and Reynolds, 1988) to determine the relative similarity of infaunal community structure among the stations. They recognized three faunal groups in the central and western basins of the Sound; no consistent groups were identified for the eastern basin (Figure 10). The three faunal groups consisted of a muddy, deep-water assemblage distributed throughout much of the central and western basins, a shallow sandy assemblage along much of the north shore of Long Island, NY, except in western portions of the Sound, and a transitional shallow water assemblage in the western portion of the Sound and along the Connecticut shore roughly between Fairfield and Branford. The general characteristics of these three groups are given in Table 5.
The muddy deep-water and the shallow sandy groups were both dominated by bivalves (Table 5). Although bivalves were also abundant in the transitional group, several species of polychaetes were the most abundant infauna at these stations. Species richness was lower in the muddy, deep-water and shallow sandy groups than in the transitional group. Reid et al. (1979) suggested that the overlap in community composition among the three groups (when all the species were considered, not just those given in Table 5) indicated that the benthic fauna in LIS is not distributed as discrete, well defined communities, but rather forms a faunal continuum from one area to another.
Ried et al. (1979) noted a distinct temporal change in the benthic communities of the Sound during their study period. Each assemblage experienced sharp declines in the total number of individuals per station, as is reflected in the declines in abundance in almost all of the dominant species listed in Table 5. The cause of this widespread population crash is not known and several potential causes were discussed by Reid (1979). Rhoads and Michael (1974, cited in Reid et al. 1979 and Reid 1979), also noted the sharp declines over this period and suggested that a major erosion event in the spring of 1973 may have altered habitat conditions to the extent that most infaunal populations experienced recruitment failures. Other potential explanations offered by Reid (1979) included "normal fluctuations in opportunistic species," trophic group amensalism (e.g. Rhoads and Young 1970), recruitment declines due to negative biotic interactions between established dense infaunal populations and settling larvae (e.g. Woodin 1974) and high levels of predation on the abundant populations found in 1972.
Although two of the community groups included opportunistic species at high population densities in 1972 (e.g. Mulinia lateralis and Polydora ligni in the muddy deep-water group, and Polydora ligni and Streblospio benedicti in the transitional group), Reid (1979) argued that since population declines were found in all species, "normal fluctuations in opportunistic species" could be discounted. Furthermore, he compared his data with that collected by Sanders (1956) at several sampling stations occupied in both studies. Recognizing the potential pitfalls in comparing data collected using different methods, his comparison suggested that the 1972 abundances were not abnormally high. He calculated that at these stations total macrofaunal abundance in 1953 - 1954 was 7,220 individuals m2 as compared to 4,993 individuals m2 in 1972 and 985 individuals m2 in September 1973 after the crash. In the absence of further information, Reid (1979) suggested that the erosion event was probably the most parsimonious explanation for the widespread population crashes, although several of the other mechanisms may have been involved. For example, extensive sediment reworking by deposit feeders may have destabilized the sediments which, coupled with a major erosion event, led to the loss of the high density communities found in 1972.
McCall's Studies in the Central Basin
Between 1972 and 1973, McCall (1977, 1978) conducted experimental and observational studies that addressed several aspects of the dynamics of infaunal communities. Primary among these was how infaunal communities responded to disturbance and subsequent successional dynamics that led to the reestablishment of the community. In the context of this work, McCall (1977) also addressed the adaptive strategies of infaunal organisms and how, in relation to disturbance, they may account for spatial and temporal differences in population abundances. The experimental studies were conducted at two sites at about 15 m depth just south of the New Haven Harbor area (Figure 8). Surveys were conducted in the central potion of the central basin of the Sound, extending from the Connecticut shore to the north shore of Long Island (Figure 8).
The experimental work consisted of setting out trays of defaunated sediment to simulate disturbances which locally extirpate the fauna, and following recolonization and succession over a period of one year. From these experiments, McCall (1977) was able to distinguish three groups of species based on their responses to disturbance and changes in population abundance during succession. Group I consisted of species which initially colonized the defaunated areas in very high numbers, including Streblospio benedicti, Capitella capitata and Ampelisca abdita. Their abundance declined significantly as succession proceeded. These species have been shown to respond in an opportunistic manner to disturbance in many studies and are often found in high numbers in impacted habitats (e.g. Grassle and Grassle 1974; Rosenberg 1976; Pearson and Rosenberg 1978). Group II species, attained peak abundances during the middle portions of succession and included species such as the bivalves Nucula proxima and Tellina agilis. Group III species were the dominants at the end of the experimental period, and as such were interpreted by McCall (1977) to represent members of the successional endpoint community (e.g. climax / equilibrium community). These included the polychaete Nephtys incisa and the bivalve Ensis directus.
Table 6 contrasts the ecological characteristics of Group I and Group III species. Group I species are opportunists which are generally small in size, live in tubes in the upper layers of the sediment, and have life histories which allow for rapid population increases. Group III, or equilibrium, species are large, mobile, live deeper in the sediment and have life histories which do not often lead to rapid population increases. Group II species have intermediate characteristics. A more detailed discussion of the contrasting characteristics of these groups of species can be found in McCall (1977) and Rhoads et al. (1978). Based on the experiments, McCall suggested that areas which are frequently disturbed will primarily consist of patches of opportunistic species, whereas areas which are disturbed less frequently will be comprised more of equilibrium type species. To determine the generality of these results, McCall (1977, 1978) surveyed portions of the central basin in LIS. He noted that the frequency of bottom disturbance was higher in shallower (<20 m) areas of the Sound. Therefore, opportunists should be found in higher abundances in these areas and that equilibrium species should be found at relatively constant abundances in deeper waters. The results of the surveys bear out this prediction. Opportunistic species displayed greater changes in abundance between years in shallow portions of the central basin and also displayed a higher degree of patchiness in their abundance distributions (measured by variance to mean ratios) relative to equilibrium species. The general information on species composition in the survey data corresponded to that provided in Reid et al. (1979) for the central basin.
McCall (1978) analyzed his survey data in more detail in order to partition the effects of disturbance, substratum type and other factors which may affect species distributions and population abundances. Note that this is the period over which there was a significant reduction in the abundance of infaunal populations in this portion of LIS as was noted by McCall (1977) and also by Reid et al. (1979, see above). He found that while faunal differences among bottom types did exist, differences in population abundance (and variation) associated with depth were more pronounced. Addressing the population crashes, McCall (1978) found that many of the affected species were bivalves and other species with long-lived planktotrophic larvae. Seventy five percent of the species which had lecithotrophic larvae or larvae which are brooded for some period of time were unaffected. Large-scale changes in the water column (e.g. suspended load, increased turbulence) may have had significant effects on the larvae and also on the adults of filter-feeding species. On an overall basis, McCall (1978) suggested that "Benthos distribution will be patchy depending on the frequency and distribution of ... disturbance and initial heterogeneity of the substratum" and that other patterns may be "most clearly related to plankton phenomena."
During the same period McCall was carrying out his work, other researchers at Yale were investigating other aspects of benthic dynamics in LIS (e.g. Rhoads et al. 1977, 1978; Yingst and Rhoads 1978). Much of this work, and that conducted by McCall, is summarized in a model proposed by Rhoads et al. (1978, see also Rhoads and Boyer 1982; Rhoads and Germano 1982, 1986) which depicts the response and structural aspects of soft-sediment communities in relation to disturbance and pollution stress (Figure 11). The model recognizes three phases in community structure along a spatial gradient with respect to pollution or along a temporal gradient with respect to disturbance and subsequent succession. A Type I community is found following disturbance or in stressed habitats. It is comprised of some combination of tube-dwelling polychaetes, amphipods and clams which primarily live in the upper few cm of the sediment. The oxygenated portion of the sediments is relatively shallow. The tube building activities of the infauna bind the upper layers of the sediments and bioturbation is low. Type III communities represent the successional endpoint or community structure under unstressed conditions. According to the model, Type III communities are comprised of deeper dwelling polychaetes, bivalves and crustaceans which through their activities mix and oxygenate sediments to a deeper level. Type II communities exhibit transitional characteristics. I will return to this model when comparing the results of the studies being reviewed.
Benthic Studies in Fishers Island Sound
Fishers Island Sound (FIS) is the eastern-most extension of LIS (Figure A). While it has not experienced the environmental degradation of the western portion of the Sound (e.g. Wolfe et al. 1991), this area supports important commercial and recreational activities and as such is an integral component of the resources in LIS. Between 1972 and 1974, three studies were conducted in FIS which focused on separate taxonomic components of the benthic communities. Franz (1976) investigated molluscan assemblages, Biernbaum (1974, 1979) considered amphipods, while Swanson (1977) studied the polychaete assemblages. The main focus of each of the studies was to determine effects of sediment type on the distribution and composition of these taxonomic groups. Samples were taken at 17 to 24 stations in FIS located on a variety of bottom types (Figure 8).
Franz (1976) identified 52 species of molluscs and analysed his data using clustering methods. These analyses indicated that the species found at the stations which comprised each cluster differed in their distribution primarily on the basis of sediment type. He identified three assembalges of molluscs living in very fine sands (Group A), medium sand (Group B) and coarse sand (Group C), and a fourth assemblage that was not sediment correlated apart from not occurring in very fine sediments (Figure 12). These groups of species were relatively similar between summer and winter. One of the groups, Group A, corresponded to the mollusc component of the Nucula / Nephtys / Yoldia infaunal mud community described by Sanders (1956) for LIS and for Buzzards Bay (Sanders 1958, 1960). Franz (1976) suggested that the distributions found in FIS might represent an extension of this benthic assemblage into very fine sand sediments.
Biernbaum's (1974, 1979) study of amphipods at the same set of stations also indicated relationships to sediment type. Multivariate analyses indicated that while several distinct assemblages of benthic amphipods could be recognized, those found in seagrass beds and on well-sorted sand bottoms, most of the assemblages were related to a clinal gradation from very fine to very coarse sediments. Differences in assembalges associated with particular bottom types were also related to feeding type. In this case, the amount of gravel was the most important sediment factor that appeared to influence feeding type. In contrast to Franz's (1976) study on the molluscs, Biernbaum (1979) found significant changes between the summer and winter. The relationship between amphipod distributions and sediment type was largely absent in the winter. Biernbaum (1979) suggested that bottom disturbance in the winter makes the sediment less stable and subject to scour.
Similar relationships were found by Swanson (1977) in her study of polychaete assemblages in Fishers Island Sound. The distribution of polychaete species was primarily related to feeding type and bottom types. Direct deposit feeders were found predominantly in muddy bottoms, indirect deposit feeders in sandy bottoms, and carnivores in coarse sand and gravel. Species diversity was usually higher in coarse sediments and increased in the winter. Polychaete abundances declined at all stations from summer to winter.
To develop a preliminary mapping of benthic communities in FIS, I combined information on the abundance of amphipod and polychaete species from Biernbaum (1974) and Swanson (1978), respectively, at the stations where both researchers took samples and performed a multivariate analysis. [I did not include data on the molluscs from Franz's (1976) study because the raw data for each sampling station was not available.] The matrix was comprised of 16 stations and 40 species of amphipods and polychaetes (Table 7). The matrix was developed by averaging summer and winter abundances of the five most numerically abundant species at each station and combining the species lists and abundances from the 16 stations. The data were then analysed using multivariate routines in NTSYS (Rohlf 1993). The similarity among stations, based on the relative abundances of all the species in the matrix, was determined using the Bray-Curtis (1957) index. The resulting matrix of station similarities was then classified using the unweighted pair-grouping (UPGMA) method (Sneath and Sokal 1973). The resulting dendrogram was analyzed to determine groupings of stations and their overall level of similarity. Each station was assigned a group designation and a GIS coverage depicting the station groupings was developed to display the spatial distribution of the groups within FIS. Descriptive statistics were generated for each species in the groups revealed by the classification using data for each station belonging to an identified group.
The analysis revealed three main clusters of stations and several stations "chaining" onto these groups (Figure 13, Table 7). The stations included in the three main clusters had more species than the stations chaining onto these main clusters. Group I was comprised of four stations. Two of the stations, Z1 and Z2, were located in Zostera seagrass beds in inshore areas (Figure 14). The other two stations, stations 16 and 17, were located in the eastern end of FIS. These two groups of stations formed sub-clusters in this group at a similarity level of about 50% but clustered together to form the group at a similarity level of 10%. The dominant amphipod species in the cluster included Corophium acutum, C. bonelli, C. insidiosum, Lysianopsis alba, Microduetopus anomalus and Phoxocephalus holbolli. The dominant polychaete species included Clymenella torquata, Lumbrineris tenuis, Prionospio heterobranchia, Scoloplos robustus and Spio filicornis. For both taxa, abundances at these stations were generally higher on an overall basis relative to the other clusters of stations. The sediment type in the seagrass beds differed from that found at stations 16 and 17, being comprised of silt-clays and coarse sand and gravel, respectively. However at all four stations the level of organic content was high both in summer and winter.
Cluster II which was comprised of five stations and two chaining stations designated as II-1 and II-2 (Figure 13, Table 7). These stations were located through the central portion of FIS (Figure 14). The dominant amphipods included Ampelisca abdita and A. vadorum, Phoxocephalus holbolli, and Unicola irrorata. The dominant polychaetes included Tharyx acutus, Lumbrineris tenuis and Harmathoe extenuata. The abundances of the polychaetes were relatively low. Two stations, 5 and 6, chained onto this main group. The communities at these stations had lower numbers of species than the main Cluster II, but the dominants were similar. At station 5 (Cluster II-1), the dominant species was Lumbrineris tenuis. The amphipods Ampelisca vadorum, Lembos websteri and Unicola irrorata were also found at relatively high abundances. At Station 6 (Cluster II-2), the suite of amphipods found in the main Cluster II were also abundant, but the dominant species was the amphipod Byblis serrata. The only abundant polychaete was Nephtys picta. The stations in these clusters were found on coarse to medium sediments in the summer, but sediment characteristics were quite heterogenous in the winter (Swanson 1977), reflecting the potential effects of winter storms. Organic content was variable in the summer, but generally high in the winter, except for cluster II-2 where sediment organic content in the winter was low.
The other large grouping of stations was Cluster III. Three of the stations (C, E and F) were located close together in the vicinity of Ram Island in shallow waters (Figure 14). The fourth station, station 12, was also found in shallow waters but further east in FIS. The chaining station, III-1, was also found in shallow waters, but in the western end of FIS, just north of Fishers Island. The dominant species in Cluster III included the amphipods Ampelisca abdita, A. vadorum, A. verrilli, Leptocherius tenuis, and Unicola irrorata, and the polychaetes Nephtys picta, Tharyx acutus, Lumbrineris tenuis and Clymenella torquata. The abundances of the dominant polychaetes were lower that those of the dominant amphipods. The dominant species at the chaining station, station 10, were similar but abundances of the amphipods were lower than in the main cluster III. These five stations were found on fine sands. Sediment grain sizes did not change greatly between the summer and winter, but sediment organic content did. During the summer, the organic content of the sediments was low to moderate and in the winter it was high. The exception was station 10, where sediment organic content remained low in the winter.
Pellegrino and Hubbard's Sound-Wide Survey
In the early 1980's energy-related activities in LIS became an important aspect of managing the Sound. With increasing population pressures along the LIS coastal zone, increasing coastal energy activities were anticipated. Within the scope of the Connecticut Coastal Energy Impact Program, Pellegrino and Hubbard (1983) conducted a benthic survey of Connecticut waters in LIS in order to provide "natural resource data bases needed to assess the impact of energy related environmental accidents and to properly plan for future expansion of energy facilities and activities." Their study comprised a natural resource inventory which focused on characterizing and quantifying benthic communities at 413 stations throughout the sound (Figure 8) with specific emphasis on shellfish populations. Sediment characteristics (Figure 4) and temperature and salinity were also recorded at each station. Each station was sampled once either during 1981 or 1982. Here, I will focus on their benthic community data. Benthic samples were collected with a 0.25 m2 Van Veen grab and washed on a 1 mm sieve. Pellegrino and Hubbard (1983) divided the sampling stations into ten regions and analysed their data to present differences in community structure among these regions. The regions and summaries of community structure in each are given in Table 8.
Pellegrino and Hubbard's (1983) data confirmed several general trends in community structure which were found previously by Reid et al. (1979) and workers in Fishers Island Sound. Species richness increased from west to east in the Sound, and the mean density of individuals per sample was generally higher in the central and eastern basins of LIS than in the western portions. Community structure in the western and central basins was dominated by a group of species found in previous studies (i.e. the bivalves Mulinia lateralis and Nucula annulata, and the polychaete Nephtys incisa), whereas dominants in the eastern portion of the Sound were more varied among the regions (Table 8). Their report also provides detailed resource maps for each region with information on substrate type, species numbers and total abundance and shellfish abundances at each station. Pellegrino and Hubbard (1983) also devised a ranking system that could be used to calculate the ecological value of any particular station based on differences in criteria such as number of species, total abundance, species diversity and the relative abundance of various shellfish species. The station-specific values were not calculated in the report however.
Detailed Analyses Using Pellegrino and Hubbard's Data
The data provided in Pellegrino and Hubbard (1983) are unique in that they represent the most spatially detailed information available on benthic communities in Long Island Sound, albeit that the sampling stations are all located in the northern portion of the Sound, in Connecticut waters. One of the main goals of my project was to assess variability in benthic community structure in LIS at several spatial scales and relationships between community structure and factors which can determine the nature, distribution and dynamics of these communities. To this end, I reanalyzed Pellegrino and Hubbard's data in order to determine in more detail the spatial distribution of various community types within the Sound and assess what factors may be responsible for the spatial patterns.
The analyses centered on species richness and community composition. To determine patterns of variation in species richness, a GIS coverage was developed which depicts the total number of species found at each of the 413 stations. This coverage is refered to as SPRICH. To assess variation in community composition, a condensed data set was developed based on the five most abundant polychaete, bivalve and arthropod species at each of the stations sampled by Pellegrino and Hubbard (1983). If any ties in the number of individuals per station were found for any set of species, all those species were included in the data set. Separate community matrices, were then developed for polychaetes, bivalves, arthropods and the overall community. The overall community matrix was developed using the most abundant species among the three taxonomic groups across all stations. A listing of the species included in each of the matrices for the separate taxonomic groups can be obtained from the author.
Classification (clustering) analysis (e.g. Ludwig and Reynolds 1988) was used to determine the similarity in taxonomic assemblages and overall community structure among the sampling stations. Analyses were performed using NTSYS software (Rohlf 1993). The similary among stations, based on the relative abundances of all the species in a matrix, was determined using the Bray-Curtis (1957) index. The resulting matrix of station similarities was then classified using the unweighted pair-grouping (UPGMA) method (Sneath and Sokal 1973). The resulting dendrograms were analysed to determine groupings of stations and their overall level of similarity. Each station was assigned a group designation and GIS coverages depicting the station groupings were developed to display the spatial distribution of the groups within the Sound. Descriptive statistics were generated for species in the groups revealed by the classification, using data for each station belonging to an identified group.
Further analyses were done using GIS overlay techniques to determine relationships between habitat characteristics and community structure. These analyses represent a "snapshot" view as the stations were only sampled once, at different times over a one and a half year period. However, they do allow us to make an initial assessment of the spatial distribution and variation in benthic community structure in the Sound, and recognize trends that can be explored further as the focus of future studies.
Species Richness
While it is well known that benthic species richness increases from west to east in the Sound, the detailed data provided in Pellegrino and Hubbard indicates a fair degree of spatial variation in species richness (Figure 15). Groupings of stations with only 0 - 5 species per sample were found in several areas of LIS including the region between Port Chester, NY, and Stamford, CT; in a band from shallow to deeper waters south of Fairfield to the east of Stratford Shoal; and in an area between Clinton and the Connecticut River. Many stations had slightly higher species richness (6 -10 species per grab) and were generally found in the same geographical areas as those with 0 - 5 species per sample (Figure 15). There were several areas of stations with moderate species richness (11 - 25 species per sample), including the areas south of the Norwalk Islands, just west of Stratford Shoal, in shallower waters south of the New Haven area, and at all depths between Branford and Clinton. The only extensive area of high species richness (26 - 60 species per sample) was from the Connecticut River east into Fishers Island Sound. However, there were a number of stations and smaller areas of high species richness in the western and central portions of LIS, with values comparable to those found in the eastern portion of the Sound. These included one station south of Stamford which had 44 species, several stations in shallow waters near the Norwalk Islands where richness ranged from 26 to 40 species per sample, several deep-water stations in the Stratford Shoal / Middle Ground area, and about 12 stations distributed throughout the central basin at varied depths which had 26 to 40 species per sample.
From the spatial distribution of species richness given in Figure 15, it is clear that while general spatial trends in species richness do occur in the Sound, they are not monotonic on a east - west and/or depth gradient. The general, large-scale trends reflect to some extent differences in several factors that may control species richness. For example, the proximity of the eastern end of the sound to a larger species pool outside the waters of LIS and physical conditions, such as salinity, which would allow establishment of those species, may contribute to higher species richness in this area. Greater habitat heterogeneity in this area may also help to increase species richness. In contrast, lower species richness in the western portion of the Sound may be due to some combination of a reduced species pool, lower habitat heterogeneity and also long-term environmental deterioration. The "islands" of higher species richness in the central and western basins might be related to higher habitat heterogeneity or other factors which may promote local increases in species richness (e.g. less harsh conditions).
Characterization and Spatial Distribution of Taxonomic Assemblages
Analyses were conducted to determine the spatial distribution of assemblages of three groups of taxa: polychaetes, bivalves and arthropods. For each taxonomic group, the numerically abundant species in the data set were identified and data matrices were developed comprising these species and their abundances across the 413 sampling sites. The matrices were then analyzed to classify communities and the resulting clusters were then mapped. The spatial distribution of the identified groupings for polychaete, bivalve and arthropod assemblages are given in Figures 16, 17 and 18, respectively. These figures are included here to indicate that these coverages and related analyses are available from the author. They will not be discussed in any more detail here.
Characterization and Spatial Distribution of Community Types
The analysis of overall community structure was based on 35 species that were most abundant throughout the Sound, including 19 polychaete species, 8 bivalve species and 8 arthropod species (Table 9). Classification analysis resulted in a dendrogram which was resolved into 12 main clusters of stations, or community types, and several sub-clusters in several of the main clusters (Figure 19, Table 9). The similarity among stations within a cluster varied. For clusters A, B, and C, within-cluster similarity ranged between 60% and 90%. For clusters D through K, similarity among stations varied to greater extent with values ranging, approximately, between 90% and 20%. One cluster, L, was a small group of stations were no fauna was found. The level of similarity among the main clusters ranged between 5% and 30% (Figure 19). The overall clustering pattern suggests that community structure is quite variable throughout the Sound. There were few distinct associations of species among the stations, as would be suggested by higher levels of similarity within clusters.
The spatial distribution of community types indicated by the classification analysis was determined by mapping out the stations comprising each main and sub-cluster (Figure 20). This map is referred to as the LISCOM coverage in the benthic GIS. Stations comprising Cluster A were located in the far western portion of the Sound, except for one station along Stratford Shoal and one station in shallow water off Sachem Head near Guilford. The community at these stations was dominated by the coot clam Mulinia lateralis, in association with moderate densities of the polychaetes Pectinaria gouldii and Nephtys incisa.
Clusters B and C were joined at a similarity level of 27%, but exhibited the highest within-cluster levels of similarity among component stations. This was especially the case among stations in sub-clusters C1 and C2. These communities were distributed throughout most of the western and central basins of LIS (Figure 20). Community characteristics of stations in Cluster B included high abundance of Mulinia lateralis and Nucula annulata, moderate abundances of several other bivalves, and high abundances of Nephtys incisa. Community types indicated in sub-clusters C1 and C2 were characterized by higher abundances of Mulinia lateralis, Nucula annulata and Nephtys incisa than in Cluster B, as well as higher abundances of several bivalves (Pitar morhuanna, Yoldia limatula and Pandora gouldiana), the polychaete Mediomastus ambiseta and the amphipod Ampelisca abdita. Differences between C1 and C2 were primarily based on the relative abundances of Mulinia lateralis and Nucula annulata.
Cluster D was comprised of a group of 3 clusters that chained on to the larger cluster formed by Groups B and C (Figure 19). This group of stations, like those in Cluster A, was found primarily in the far western portion of the Sound, with sporadic occurrences in the central and more eastern portion of LIS (Figure 20). High abundances of the polychaete Pectinaria gouldii, the amphipod Corophium acheruscum, and moderate densities of several bivalves including Mulinia lateralis, Nucula annulata, Pitar morhuanna and Tellina agilis were found at these stations. Clusters E, F and G also comprised stations which chained onto the larger cluster formed by stations in clusters A - D. The community at stations comprising Group E, which were found primarily in the western Sound and at a few stations in the middle basin (Figure 20), were comprised of species which were present at stations in clusters A - D, but at relatively low abundances. Cluster F was comprised of stations which were located along Stratford Shoal, nearshore areas between the mouth of the Housatonic River and Clinton and along an offshore vector south of Sachem Head. The benthic communities in this group of stations were comprised by high densities of a number of species including very high abundances of Mulinia lateralis, the polychaetes Clymenella zonalis and Mediomastus ambiseta, and relatively high abundance of Nephtys incisa, Pectinaria gouldii, Streblospio benedicti, Nucula annulata and Tellina agilis. Cluster G was comprised of stations which had only 10 of the 35 dominant species present, and these were found at low abundances, except for Pectinaria gouldii, Clymenella zonalis, Asabellides occulata and Pitar morhuanna. All Cluster G stations were found in the western end of the Sound except for one nearshore station south of Branford (Figure 20).
Cluster H was comprised of three sub-clusters. Stations comprising sub-cluster H1 were found mainly in four areas of the Sound: southeast of the Norwalk Islands, along Statford Shoal, in nearshore waters between Milford and West Haven, and in large concentration at all depths between Guilford (Sachem Head) and Old Saybrook (Figure 20). The community at these stations was the most species-rich found, with 31 of the possible 35 dominant species present. Although many of these were found in low abundance, several occurred at moderate to high abundances including Asabellides occulata, Tellina agilis, Spiophanes bombyx and Clymenella zonalis. Sub-cluster H2 was also comprised of stations which were relatively species-rich, but the composition of numerically abundant species differed from H1. Stations in H2 had communities dominated by crustaceans, including Ampelisca abdita, Ampelisca vadorum and Corophium acheruscum. This community type was distributed throughout the Sound, at several deeper water stations in the western basin, at nearshore stations between Branford and Guilford, and in a more extensive area south of the mouth of the Thames River in the eastern basin (Figure 20). Stations comprising cluster H3 were primarily found east of Mattituck Sill in the eastern basin of LIS (Figure 20). This group of stations was characterized by communities which only had about half of the species included in the analysis, and all at relatively low abundances, in contrast to H1 and H2. The most abundant species was a polychaete, Aricidea jeffersii.
Most of the stations included in Cluster I were east of the Connecticut River (Figure 20), between Old Lyme and Waterford and in Fishers Island Sound. This group of stations had a high number of species and was dominated by the mussel Mytilus edulis, and two polychaetes, Cirratulis grandis and Prionospio heterobranchia. Several species were also found at moderate abundances at these stations including Aricidea jeffersii, Cirratulis grandis, Ampharete arctica, Prionospio tenuis, and Aeginina longicornis (Table 9). There were four stations which comprised Cluster J and these were located throughout the Sound (Figure 20). There were only six species in this group and their abundance was very low (Table 9). Cluster K was comprised of stations in deep to shallow waters, primarily between Clinton and Old Saybrook, although one station was found near Watch Hill, RI, and another in the western basin of LIS south of the Norwalk Islands. This cluster was characterized by moderate species richness, and numerical dominance by the amphipod Protohaustorius wigleyi. Cluster L was comprised of stations where no fauna was found.
The variation associated with mean abundance values for species comprising each cluster type was high in a large number of cases (Table 9). This variation may be due to: a) multi-scale differences in physical factors among stations, b) temporal variation due to differences in sampling times and/or c) other factors. The variation indicates that while clusters formed by the classification analysis can be recognized as community types, individual stations within clusters differed to various extent from the characteristics determined by calculating mean abundances per station for the each of the species in the analysis. As such, any subsequent analyses and interpretations should look towards assessing this variation and incorporating it into interpretations and models of community dynamics. This subject is discussed further when the results of the various studies conducted in the sound are compared below.
Relationships between community variation and habitat characteristics
The results of the classification analysis suggests that community structure varies in LIS on several spatial scales. Sound-wide trends are evident in the general make up of communities and there is also a fair degree of local variation in community structure. For example, communities represented by Clusters A - G are primarily found in the central and western basins of LIS, whereas communities represented by Clusters H - K are generally found in the eastern end of the Sound (Figure 20). Each cluster also usually had one geographical "hot spot" where many of the stations belonging to a cluster were found. In some cases smaller spatial clusters were distributed over large areas of the Sound (e.g. Cluster H1), in others there was only one cluster of stations (e.g. Cluster K). However, many of the stations belonging to a particular cluster were also found spread throughout LIS (e.g. Clusters D, E, and H2).
The relative levels of among-station similarity within clusters also provide some information on variation in community structure at different spatial scales. For example, the highest overall levels of among-station similarity were found in Clusters B and C, and in sub-clusters C1 and C2 in particular. These two groups also had the highest among-cluster similarity. This suggests that the local variation in community structure among stations in both clusters is lower than for stations in other clusters, and that transitions in community structure among adjacent or nearby stations in Clusters B and C are less variable. Indeed, the primary difference in community structure among Clusters B and C was the relative difference in the abundances of Mulinia lateralis and Nucula annulata. This situation can be contrasted with relatively higher levels of among-station variation in Clusters H and I. Stations belonging to these groups had a higher level of local variation, and transitions in community structure among these clusters were more distinct. Even though the stations were in the same cluster and may have been geographically close to one another, the variation in species composition was reactively high.
This suggests that spatial variation in community structure is low for large areas of the western and central basins and relatively higher in the Narrows in the west and in the eastern basin. Overall trends in community variation may be related to several habitat characteristics including sediment grain-size, the geographical location of the habitat, as suggested above, and also the size of the habitat. These relationships may be similar to the various factors that appear to control species richness and diversity (e.g. Hart & Horowitz 1991). GIS habitat coverages such as those generated during this project (see above) and community coverages, as developed here from Pellegrino and Hubbard's (1983) study can be used to explore these relationships by relating habitat and community characteristics in specific locations and habitats. As examples, I addressed the question of how variation in community structure might be related to sediment type and the size of the habitat.
To determine the general relationship between sediment type and species richness and abundance, Pellegrino and Hubbard's data on species richness and abundance were sorted in relation to the bulk sediment characteristics at each station (i.e. the BULKSED coverage). The relationships between species and community richness and habitat area were explored by overlaying the LISSED coverage with the SPRICH and LISCOM coverages and sorting the data on the different patches of sediment and their areas.
Both species richness and abundance varied significantly with sediment type (Table 10). Species richness was usually highest in sediments that were comprised primarily of gravel and shell (Figure 21). This may be related to greater heterogeneity associated with these types of sediments. The lowest species richness was found in mud, sand with rocks and rocks. Low species richness in the latter two sediment types may, however, have been related to sampling problems associated with trying to take grab samples on these bottom types. In muds and sands, species richness increased as sediment heterogeneity increased with the addition of other sediment components, i.e. gravel, mud or sand, and/or shell. The highest abundances of individuals were found in gravel sediments with about 1,200 individuals per 0.25 m2 (Figure 22). Mean abundances in other sediment types generally ranged between 100 and 300 individuals per grab. Abundances in muds and sands were usually higher when other sediment components were found. These trends indicate that infaunal communities in LIS will generally have more species and higher abundances as sediment heterogeneity increases. Interestingly, the differences in species richness and abundance among sands and muds, the most common sediment types in LIS, were not great, suggesting that other factors, such as geographic position or patch size, may play an important role in shaping community structure as well.
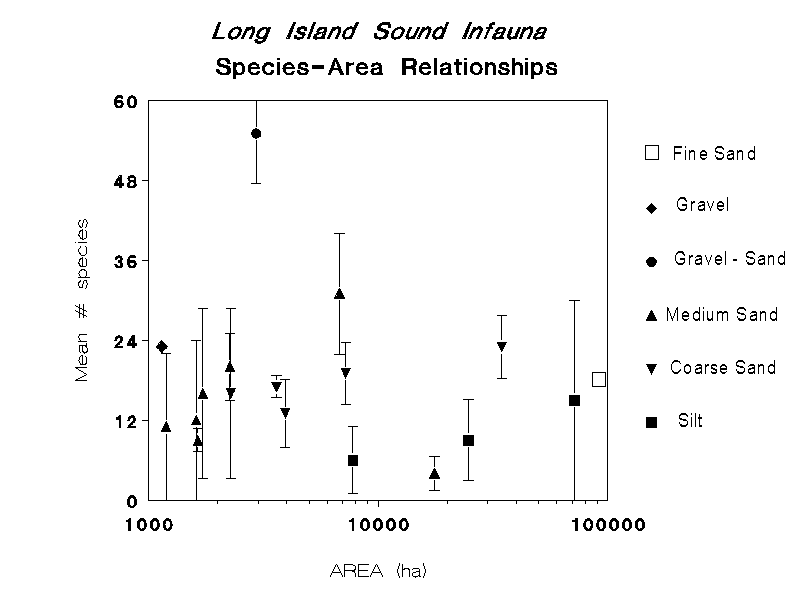
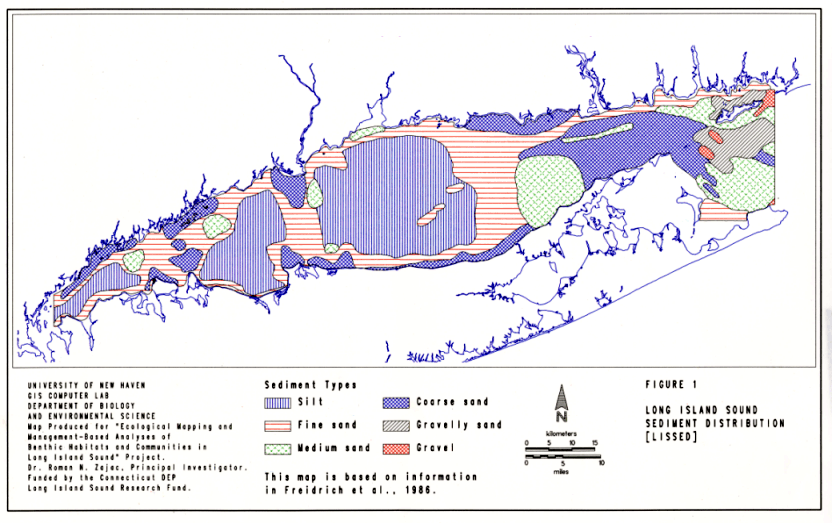
The relationship between species richness and patch size in LIS differs depending on sediment type (Figure 23). In this analysis, there were three sediment types for which there were patches of different size that could be analyzed: medium sand, coarse sand and silt. In medium sands, the mean number of species per sample varied between 10 and 20 in patches on the order of 1000 to 2000 hectares, increased sharply to about 30 species in a patch 7,000 hectares in size, but decreased to the lowest value in the largest patch. This medium sand patch is located on the boundary between the eastern and central basins of the Sound, along the north fork of Long Island (Figure 1). The medium sand patch with the highest species richness is located in the eastern basin between Niantic Bay and the Thames River.
In coarse sands, there was a slight increase in mean species richness per core with patch area (Figure 23). In the largest coarse sand patch, comprising much of the eastern basin, mean species richness per sample was 23, in the smallest patch, south of the Norwalk Islands, a mean of 15 species per sample was found. In silt patches, mean species richness per sample increased with patch size from approximately six to 14 species per sample. The sizes of the three silt patches also correspond to their location in the Sound, with the smallest being in the Narrows area, the next largest in the western basin, and the largest being the mud patch comprising much of the central basin (Figure 1). As such, the trends in species / patch area are confounded by geographical position.
Perhaps the most striking aspect in these relationships is the high variation in species richness found in patches of all sizes (and geographical position) and sediment types. For example, in the largest silt patch the variation in species richness ranged over most of the values, indicating that smaller-scale factors may be influencing species richness. One important factor may be variation in overall sediment composition. For example, the large silt patch in the central basin is defined as just silt in the LISSED coverage, but the BULKSED coverage indicates a high degree of variation in bulk sediment characteristics among the stations found in this patch (compare Figures 1 and 4).
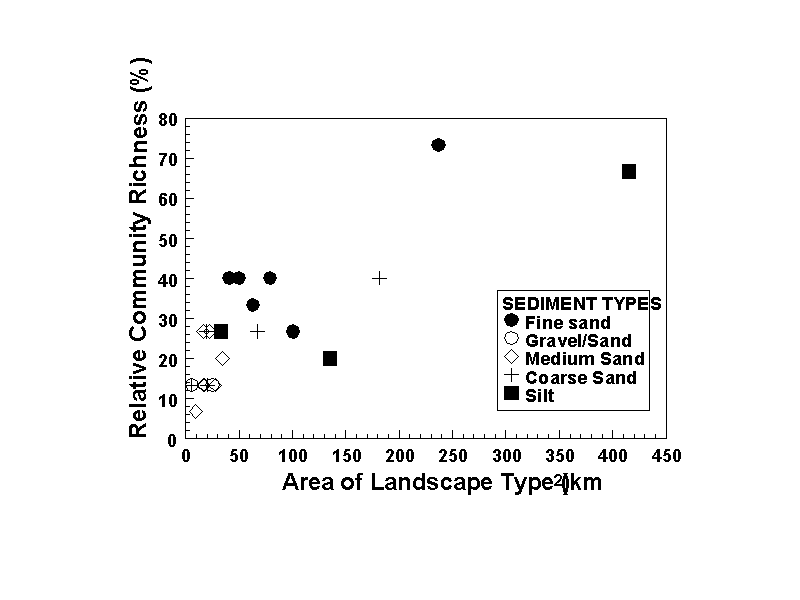
Patch size and the richness of community types that might be found in a particular patch may also be related. Are we likely to find more different types of communities in larger patches than in smaller ones, and is this dependent on the sediment type? For example, larger patches may have greater habitat heterogeneity and therefore might support a greater variety of assemblages. For LIS infaunal communities, the richness of community types found increases rapidly with patch size when all sediment types are considered collectively (Figure 24). Between patch sizes of 10 to 50 km2 the relative community richness (i.e. the percentage of overall community types recognized in the Sound based on the LISCOM coverage) rose from 6 to 40%; large increases above this did not occur until patch size increased over 230 km2. At these patch sizes, relative community richness increased to about 70%. However, there were differences between sediment types. Community richness did not increase over the small size range of gravel / sand patches. Richness increased strongly in medium sand from a low of 3% to a maximum of 26%. No patches >50 km2 were included in the data for these sediment types. In coarse sands, richness increased to a maximum of 40% in a patch about 190 km2 in size. In fine sand and silt patches there appeared to be a threshold value of community richness. For muds this value was between 21 and 28% and for sands it was between 28 and 40%. The highest community richness was found in fine sands and silts, but only in the very large patches located in the central basin.
These analyses suggest that the structural characteristics of infaunal communities in Long Island Sound may be related to habitat size in several ways. The mean number of species that may be collected per sample shows a weak increase with patch size (Figure 23), and variations about the means suggest that smaller-scale habitat characteristics will have a stronger influence on local species richness. This may especially be the case for some sediment types, such as muds, in which the full range of species richness was observed in patches of varying size. A key factor influencing local species richness may be small-scale sediment heterogeneity (Figure 4).
With respect to community richness, there does appear to be an overall increase in the variety of assemblages with increasing patch size. However, the overall trend was comprised of data for several different sediment types, and when considered separately the trend is less evident for specific sediment types. Coarse and medium sands show the strongest relationship with area. It may be that for these sediment types, habitat heterogeneity increases to a greater extent as area increases than in other sediment types. These findings are tentative and data from a wider array of patch sizes is needed to explore these relationships in more detail. More detailed analyses should also be conducted as well which account for sampling effort. The trends in species and community richness may be related strictly to area, apart from ecological factors such as habitat heterogeneity, in that as area increases the probability of accumulating and sampling new species and combinations of species may also increase.
Comparison of the Results of the Studies
The studies reviewed above provide a basic understanding of the spatial structure of soft-sediment communities in Long Island Sound with respect to species composition and possible "states" of relative species abundances. These "states" (Figure 20) represent community types to the extent that they are temporally consistent. The spatial resolution of the information available is high in the central basin, in Fishers Island Sound, and more generally in Connecticut waters of the Sound. However, many portions of the LIS benthos have only been sampled once and therefore it is difficult to assess the temporal dynamics of these communities. This is especially the case in terms of detecting any long-term trends. Several studies have looked at changes in community and population characteristics over several years at selected sites (e.g. McCall's 1977 data on changes in ambient community structure; Zajac and Whitlatch's (1988, 1989) studies of the population dynamics of Nephtys incisa). These studies generally show that benthic communities and populations in LIS exhibit expected seasonal changes in abundance and composition, as does the limited temporal data in the surveys reviewed above.
However, on a long-term basis, component species populations in the community can fluctuate to various extent. For example, Zajac and Whitlatch (1988, 1989) discuss fluctuations in the abundance of Nephtys incisa in the central basin of LIS over periods of several years that may be related to poor recruitment in some years. The Nephtys - Yoldia community that Sanders (1956) recognized was generally present at the time Reid et al. (1979) and McCall (1977 and 1978) did their sampling in this area of the Sound, although both of the latter studies did find different relative species abundances. Likewise the species groupings identified in the re-analysis of Pellegrino and Hubbard's (1983) data indicate that the community types in the central basin are generally dominated by Nephtys incisa, Nucula annulata and Mulinia lateralis. Yoldia limatula was found at high densities only in one community type identified via the classification analyses (Cluster C1). This community type was found at just three stations in the central basin (Figure 20). Thus, general community types appear to be consistent, but longer-term changes may have occurred in the populations of some of the dominant organisms. For example, in Sanders' (1956) study, the abundance of Mulinia lateralis was generally low, but subsequent surveys indicated that this was a numerically dominant species in various areas of the Sound. These trends, and those discussed above, indicate that consistent groups of species, forming community types, are recognizable in portions of LIS, such as the central basin. For other areas, our knowledge is limited and what constitutes the climax community under prevailing conditions is still an open question. Future and ongoing studies (e.g. U.S. EPA EMAP) of the benthos will help to fill in these gaps and allow scientists to develop a better understanding of benthic community dynamics. In turn, this will enhance the use of benthic community structure as a monitoring tool to assess coastal and estuarine environmental quality.
Benthic communities are important components of many monitoring programs. The main intent of such monitoring is to determine to what extent the benthic communities are impaired based on knowledge and models of what constitutes healthy and productive communities under ambient conditions. As discussed above, one such model is that proposed by Rhoads et al. (1978), which is illustrated in Figure 11. This model was developed based on studies conducted in the central basin of the Sound and has been used to map and interpret community changes and sediment organism relationships. Applications of the model include, for example, responses to natural physical disturbances such as storms, dredge spoil disposal and other types of human impacts, and mapping benthic habitats to determine to what extent they are comprised of impacted communities in the absence of direct knowledge of any impacts (e.g. Rhoads et al. 1978, Rhoads and Germano 1986). The key element of the model is the recognition of specific stages of benthic community development following a disturbance. The stages can also represent different states of degradation in response to a pollution gradient. Therefore, following disturbances such as storms or dredge spoil disposal events, or in habitats that are degraded by pollutants, the communities will be dominated by opportunistic species, the infauna will be concentrated in the upper portions of the sediment, sediment binding may be high and there will be a shallow redox potential discontinuity (RPD) level. This is referred to as a Type I or Stage I community. In contrast, unimpacted and successional endpoint, or climax, communities will be comprised of more equilibrium type species which live deeper in the sediments. Their activities will tend to mobilize the sediments and also to create a deeper RPD layer. This is referred to as a Type III or Stage III community.
As shown above, Long Island Sound is comprised of a variety of habitats and communities. The question can be posed, "Can we recognize the community types predicted by the model on a consistent basis throughout the Sound, across the variety of benthic habitats, and to what extent do they represent different stages of community development and/or impairment (i.e. climax or ambient, as opposed to impacted, communities)?" By addressing this question we can begin to fine tune our hypotheses concerning how infaunal communities respond to disturbance, what might occur after human impacts and how we interpret the results of monitoring studies. Recognizing what constitutes the successional endpoint is critical, as it is the community configuration against which we determine the level and scale of environmental impact.
We can begin to answer this question by considering the results of the sound-wide surveys that have been conducted. Reid et al. (1979) recognized three community types in the central and western basins of LIS; no consistent community groupings were discerned in the in the eastern portion of the Sound. Of note is that in each of three community types recognized, species composition was a mixture of species that would be characteristic of Type I (Mulinia, Ampelisca, Polydora, Streblospio ), Type II (Nucula, Tellina ) and Type III (Nephtys, Ensis, Pherusa) communities. In the regions surveyed by Pellegrino and Hubbard (1982), we can see a similar trend (Table 4). In Regions I - III (the eastern basin), the communities were dominated by Mulinia, Nucula, Pitar and Pectinaria; in Regions IV to VI (central basin) a suite of species similar to that described by Sanders (1956) and McCall (1978) was dominant; while in Regions VII to X a variety of polychaete and amphipod species were the dominant species. The multivariate analysis of Pellegrino and Hubbard's (1983) data (Figure 20) reinforces the view that benthic communities in LIS vary on several spatial scales and that this variation is based on different degrees of change in species composition and abundance.
Each of the assemblages noted above were comprised of species which would exhibit the organism / sediment relations predicted by each stage in the Rhoads et al. (1978) model, and no consistent Type III assemblage is easily identified. The communities in the Sound appear to be much more heterogenous in terms of the types of species they are comprised of. This suggests several possibilities. First, survey type studies may not be able to resolve the types of differences in community structure that the model predicts. The survey studies had little temporal resolution thus not allowing for any insights as to the temporal dynamics of the assemblages recognized. As such, given the high variation in community structure that is evident, it would be difficult to interpret the stage of succession or deterioration for any particular area and, indeed, the direction of change. On a large-scale, if we interpret the model strictly, the data suggest that there is high spatial variability in the distribution of Type I, II and III communities and that most areas are at some intermediate stage of succession. As such, spatially controlled and more fine-grained temporal sampling may be necessary to accurately distinguish between natural variability in community structure and human impacts.
The second possibility is that the communities described in the surveys are not structured as predicted by the model. Rhoads and Germano (1986) have noted mixtures of Type I and III communities. Therefore, the variety of infaunal assemblage depicted, for example, in Figure 20 may represent different successional endpoints, and community "states" with related characteristics and dynamics. These communities may exhibit different successional pathways and associated time frames. However, the sediment / organism relations predicted by the Rhoads et al. (1978) model (Figure 11) may still hold. A simple graphical model of this alternative view is shown in Figure 25. Each of the three community types (Types I, II and III) represent particular endpoints (or climax communities) which are usually found in response to a combination of factors including, for example, the available species pool, physiological tolerances, ability to inhabit specific sediment types, water depth, and natural patterns of periodic and stochastic temporal variation. These community types can be recognized as persisting through time under prevailing natural conditions. because of environmental heterogeneity combinations of community types, e.g. a Type I/ Type III community, will be found. When these community types are disturbed the successional pathways will also differ. For example, for a Type I community, comprised of primarily short-lived species living in the upper layers of the sediment with characteristics noted in Table 6, succession may proceed relatively rapidly, and without easily recognizable seral stages. In contrast for Type III communities, succession may proceed more slowly along the pathway predicted by the model shown in Figure 11. I feel that the potential variation in community dynamics in estuarine systems such as LIS can be quite high. A greater richness of persistent community types, and associated successional dynamics may be present than that depicted by the Rhoads et al. (1978) model. This potentially can make the task of interpreting community structure relative to environmental assessment and management issues more difficult. Therefore, it may be wise to be more cautious in applying this model to interpret changes in and status of benthic communities in the Sound. Only one controlled successional study has been conducted in the offshore waters of Long Island Sound (McCall 1977). This was in the mud habitats of the central basin.
Whilst mud sediments comprise a large portion of the LIS seafloor, the data and analyses presented above point to a high degree of habitat and community heterogeneity throughout the Sound. We know a fair amount about the spatial heterogeneity, but too little work has been done to adequately assess temporal heterogeneity in offshore infaunal communities. Thus, some important questions remain regarding seafloor habitats and communities in LIS. Primary among these is, "Given this information can we adequately assess changes in benthic community structure in relation to further environmental degradation and/or following activities to improve conditions in the Sound?" Part of the answer to this question rests on how well we can interpret community changes relative to anthropogenic signals in the context of natural variations. Comparison of the survey data and predictions of the successional models suggests that a complete picture of soft-sediment community structure and dynamics in Long Island Sound is not available at this time. Further information is needed to determine the temporal dynamics of the varied benthic communities in LIS relative to their spatial distribution and relationships to habitat characteristics. Based on this additional information a better framework for assessing changes and ecological interactions (e.g. food web dynamics) can be developed. Future work can address these gaps in our knowledge by studying benthic communities in several areas of LIS with the general goals of:
1. Determining the spatial and temporal characteristics of soft-sediment communities within and among different basins in the sound,
2. Assessing these differences relative to models of soft-sediment community dynamics, and
3) Providing a more detailed description of the ecology of these habitats and communities for management needs.
[an error occurred while processing this directive]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}