

Potential Effects of Climate Change on Ambystoma barbouri (Streamside Salamander)
Links
- Document: Report (1.57 MB pdf) , HTML , XML
- Larger Work: This publication is Chapter C of Effects of climate change on fish and wildlife species in the United States
- Data Release: Data Release - U.S. Geological Survey—Gap Analysis Project species range maps CONUS_2001
- Download citation as: RIS | Dublin Core
Acknowledgments
We would like to thank Andrea Drayer for providing insight on streamside salamander ecology and Catherine Nikiel with the Oak Ridge Institute for Science and Education Research Program through the U.S. Geological Survey Southeast Climate Adaptation Science Center for comments on an earlier draft of this report.
We would like to thank Jennifer Cartwright and Chris Pearl of the U.S. Geological Survey for their edits and comments to this report and William Farmer of the U.S. Geological Survey for insight on future stream conditions. This research was funded by the U.S. Geological Survey Midwest Climate Adaptation Science Center and the Southeast Climate Adaptation Science Center.
Abstract
Ambystoma barbouri (streamside salamanders) are stream-breeding mole salamanders that rely on seasonally intermittent, fishless streams for egg and larval development but are primarily fossorial as adults. Climate-driven changes are likely to alter streamflow duration, peak, and seasonality within the range of A. barbouri, reducing reproductive habitat and larval survival. Although future changes in precipitation volume within the geographic range of A. barbouri are uncertain, in the next 90 years, increasing temperatures will likely increase potential evapotranspiration. Decreasing ratio of precipitation to potential evapotranspiration will likely shorten flow duration for intermittent streams, potentially causing earlier stream dry downs before larval metamorphosis. Increased temperatures may also shorten developmental periods buffering A. barbouri larvae from the effects of increased stream no-flow days. Additionally, precipitation in the future will increasingly fall in heavy rainfall events. Heavy rain and subsequent flooding during early larval stages may displace A. barbouri larvae from fishless pools into downstream reaches with vertebrate predators that can reduce survival. Finally, agriculture and urban land cover may amplify the stresses of climate change on A. barbouri, altering reproductive habitat and reducing survival of larval, juvenile, and adult life stages.
Purpose and Scope
The purpose of this report is to provide an overview of potential direct and indirect effects of climate change on the Ambystoma barbouri (streamside salamander) life cycle and habitat based on peer-reviewed literature and government reports (table 1). This report focuses on A. barbouri populations and climate change within the States of Kentucky, Illinois, Indiana, Ohio, Tennessee, and West Virginia based on existing species range maps (USGS GAP, 2018; fig. 1).
Table 1.
Potential direct and indirect effects of climate change on Ambystoma barbouri.| Effect | Climate factor | Direct mechanisms | Indirect mechanism | Compounding stressors | Citations |
|---|---|---|---|---|---|
| Decreased reproduction and recruitment | Decreasing precipitation to evapotranspiration ratio | Larval mortality Early metamorphosis |
Increased predator access | Undetermined | Petranka (1984b), Petranka, (2010), Semlitsch and Wilbur (1988), Semlitsch and others (1988), Holomuzki (1991), and Micheletti and Storfer (2017). |
| Heavy precipitation events | Physical trauma to larvae from flooding | Larval drift to fish-occupied habitats Fish corridors to access larval pools |
Undetermined | Sih and others (1992), Segev and Blaustein (2014), and Petranka (1984b). | |
| Decreased survival | Increasing temperature | Desiccation | Not applicable | Atrazine exposure in agricultural streams reducing salamander water conservation behavior | Rothermel and Luhring (2005), Rothermel and Semlitsch (2006), and Rohr and Palmer (2005). |
| Reproductive habitat degradation in urban environments | Decreasing precipitation to evapotranspiration ratio | Streams dry faster Larval mortality Early metamorphosis |
Not applicable | Increased salinity in urban environments | Hammond and others (2021), Mosley (2015), Drayer and others (2020), and Kaushal and others (2017). |
| Heavy precipitation events | Higher peak flows Physical trauma to larvae |
Increased runoff of pollutants Larval drift to fish-occupied habitats |
Undetermined | Wu and others (2013), and Drayer and others (2020). |
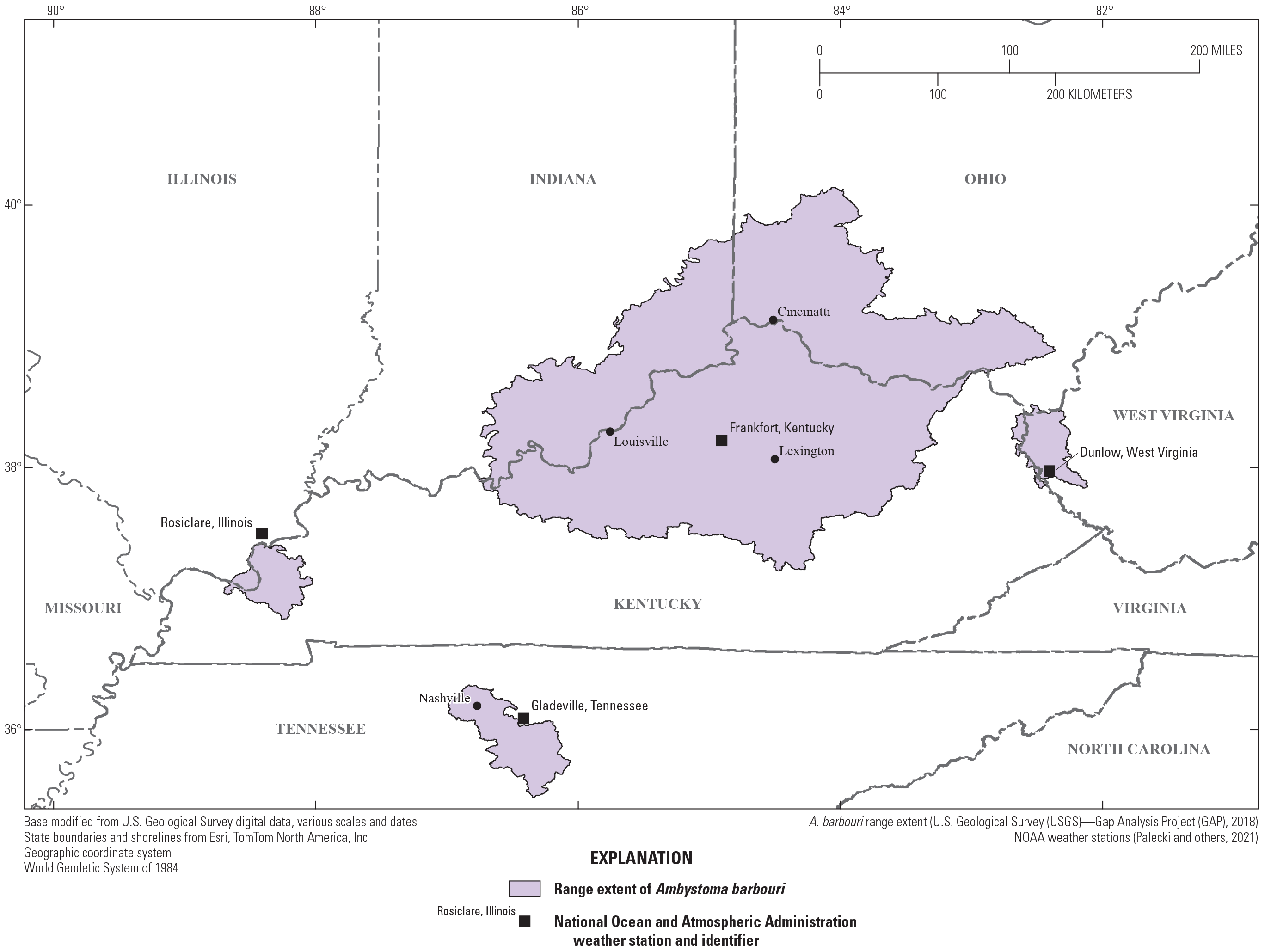
Range of Ambystoma barbouri (streamside salamander).
Climate Context
A. barbouri occupies areas of southeast Indiana, southwest Ohio, and central Kentucky, with disjunct populations in Illinois, Tennessee, and West Virginia (fig. 1). Kentucky, the State with the majority of A. barbouri’s distribution, has a seasonal climate with warm summers and cool winters (Runkle and others, 2022). The Frankfort, Kentucky weather station, at the center of A. barbouri’s distribution (Micheletti and Storfer, 2015), recorded climate normals for 1991–2020 with an average daily high temperature in July of 30.9 °C and average daily low of −3.8 °C in January (fig. 2). Precipitation in Kentucky, southeast Indiana, and southwest Ohio is abundant throughout the year. Monthly precipitation normals in Frankfort, KY ranged from a low of 80.0 millimeters (mm) per month in August to 129.5 mm per month in May. Climate normals for the three disjunct populations in Illinois, West Virginia, and Tennessee are similar to the core of the range; precipitation is approximately constant throughout the year peaking in the spring and early summer and lowest in the late summer and early fall. Between 1900 and 2020, weather stations in Kentucky experienced an average of 2.3 days per year with more than 50.8 mm of precipitation (Runkle and others, 2022). Heavy rainfall events already appear to be increasing. Since 1970, 59 percent of years have been at or above average, potentially related to a general trend of increasing precipitation in the eastern United States since 1970 (Strong and others, 2020). In the 5-year period of 2010–2014, there were on average more than 3.5 days per year with more than 50.8 mm of precipitation (Runkle and others, 2022).

Summary of historical average monthly temperature and precipitation normals from 1991 to 2020 at weather stations in the core and three disjunct distribution areas for Ambystoma barbouri. A, Frankfort, Kentucky, the center of the core of A. barbouri’s distribution; B, Rosiclare, Illinois; C, Dunlow, West Virginia; and D, Gladeville, Tennessee.
Hydrological Context
A. barbouri reproduces in first and second order (Micheletti and Storfer, 2015) primarily intermittent limestone streams (Drayer and others, 2020) that alternate pools and riffles that form natural barriers to fish upstream movement (Petranka, 2010). A. barbouri occasionally breeds in ponds (Petranka, 2010) and streams with permanent pools containing predatory sunfish (Storfer and others, 1999).
For intermittent streams in the contiguous United States, the ratio of precipitation to potential evapotranspiration is an important predictor of the timing and duration of no-flow conditions ((Hammond and others, 2021). Within the A. barbouri range, flows tend to be highest in late winter/early spring and lowest in the late summer/early autumn, with intermittent streams commonly running dry in the autumn (Eng and others, 2016; Hammond and others, 2021). In Kentucky, Illinois, and Ohio, heavy autumn precipitation events are infrequent and not large enough to counter evapotranspiration and low groundwater levels during this season. During typical years, autumn has days with no flow. Historically, during drier years, the number of no-flow days were observed earlier and more frequently, with more dry periods in the late summer and autumn. For more information, see Eng and others (2016).
Climate Change Projections
To explore potential future climate conditions in the study area, we analyze downscaled projections based on Intergovernmental Panel on Climate Change Coupled Model Intercomparison Project Phase 5 models for two greenhouse gas emissions scenarios: a moderate emissions scenario (representative concentration pathway 4.5 [RCP4.5]) and a high emissions scenario (representative concentration pathway 8.5 [RCP8.5]). The output from 20 global climate models with the necessary variables is statistically downscaled using the Multivariate Adaptative Constructed Analogs version 2 (MACAv2) method; this is a multistep constructed analog approach that establishes relations between global climate model output and historical climate observations. Outputs are bias corrected by climate variable to develop higher resolution and localized projections with about a 4-kilometer spatial resolution. The first realization (r1i1p1) of each model is used except for CCSM4 where the sixth realization (r6i1p1) is used. Output can be accessed through https://www.climatologylab.org/maca.html (Abatzoglou and Brown, 2012). MACAv2 was chosen because it is widely vetted and used to explore climate change effects, provides adequate spatial resolution to distinguish differences across A. barbouri’s distribution, and has been documented to better capture signals in rainfall extremes and frequency compared to other downscaled products (Wang and others, 2020; Wootten and others, 2021).
In the distribution of A. barbouri, most global climate models (GCMs) suggest that precipitation will increase during winter and spring by the end of the century under moderate (RCP4.5) and high (RCP8.5) emission scenarios (fig. 3). Changes in summer and fall precipitation volume are more uncertain; climate models project both wetter and drier future conditions (fig. 3). In the lower Midwest and upper Southeast, more frequent heavy precipitation events will likely reflect a larger proportion of precipitation volume under both RCP4.5 and RCP8.5 emissions scenarios (Reidmiller and others, 2018). Increasing temperature, especially in the summer, will lead to increased rates of evapotranspiration (fig. 4). For the region occupied by A. barbouri, climate projections suggest an increase in drought frequency, severity, and duration (Jeong and others, 2014).

Projected seasonal precipitation for four 30-year periods from 1971 to 2099 in the distribution areas for Ambystoma barbouri. Projections are centered on weather station points used for climate normals (figs. 1 and 2). A, Frankfort, Kentucky, the center of the core of A. barbouri’s distribution; B, Rosiclare, Illinois; C, Dunlow, West Virginia; and, D, Gladeville, Tennessee.
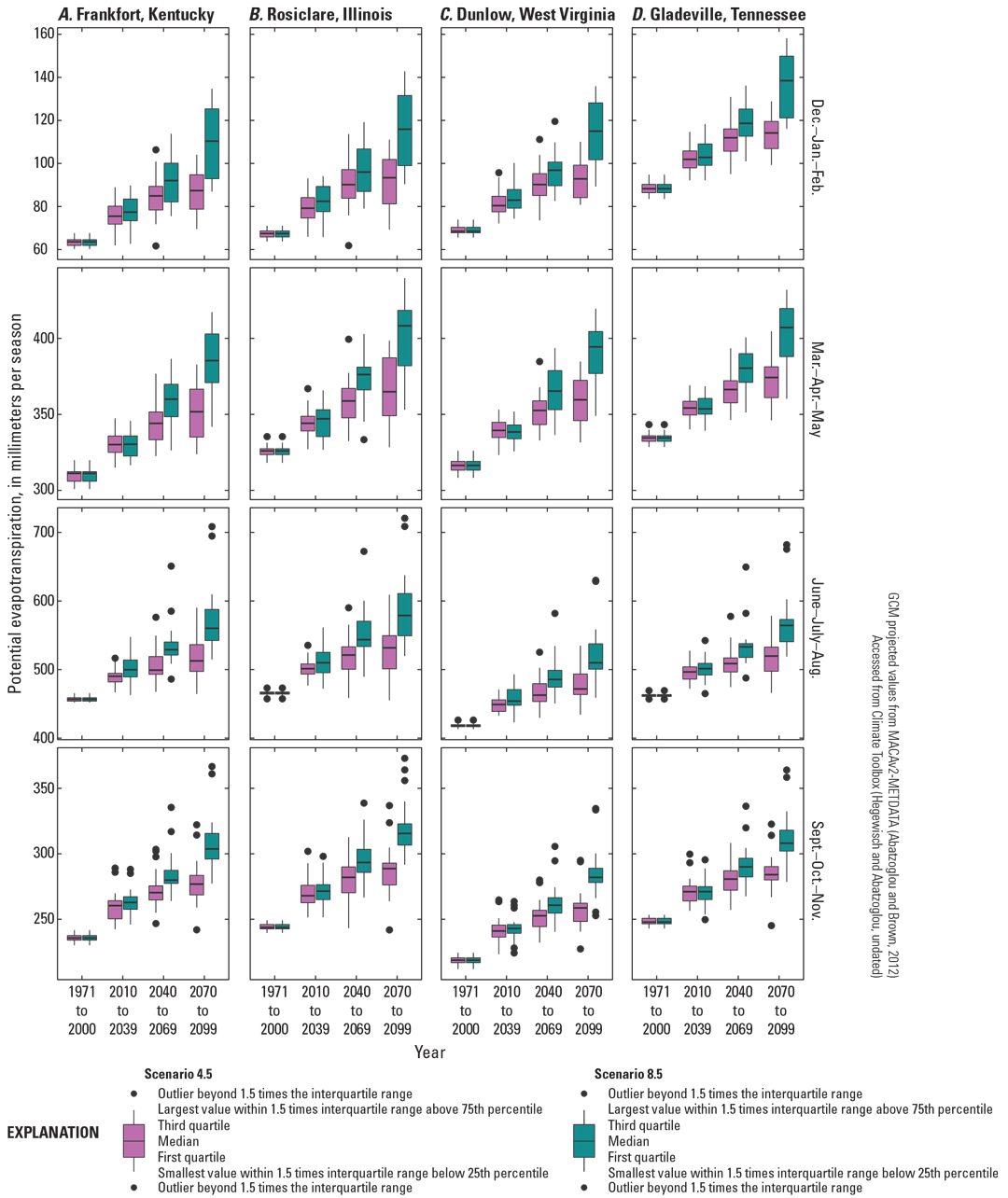
Projected seasonal potential evapotranspiration for four 30-year periods from 1971 to 2099 in the distribution areas for Ambystoma barbouri. Projections centered on weather station points used for climate normals (figs. 1 and 2). A, Frankfort, Kentucky, the center of the core of A. barbouri’s distribution; B, Rosiclare, Illinois; C, Dunlow, West Virginia; and, D, Gladeville, Tennessee. Potential evapotranspiration represents maximum water demand for a well-watered grass surface using Penman-Monteith method (Allen and others, 1998; Abatzoglou, 2013).
Future Hydrology
With precipitation increasingly falling in heavy events and increasing temperatures (see the “Climate Change Projections” section), regional and national hydrology models predict changes in timing and flow for intermittent streams used by A. barbouri. As future precipitation increasingly falls in variable, high-volume events, peak flow and frequency of high-water conditions are likely to increase. In the continental United States, the number of no-flow days for an intermittent stream is inversely correlated with the ratio of precipitation and potential evapotranspiration (P/PET) (Hammond and others, 2021). Increasing summer evapotranspiration (fig. 4) without equal increases in seasonal precipitation (fig. 3) will likely lead to earlier and more no-flow days in intermittent streams (Hammond and others, 2021). Most GCMs project a decrease in P/PET across the distribution of A. barbouri for summer, fall, and winter; some models indicate slight increases in spring under a moderate emissions scenario (fig. 5). A decrease in P/PET may advance the first no-flow date, increase the duration of no-flow conditions for intermittent streams, or both (Hammond and others, 2021).
Simulations from the Variable Infiltration Capacity model using 16 GCMs and 2 emissions scenarios (RCP4.5 and RCP8.5) predict that in the Northeast and Midwest, increasing heavy rainfall events will lead to increased peak flows and more frequent high flow conditions (Demaria and others, 2016). Even in areas with increasing annual precipitation, increasing atmospheric demand on water resources will likely lead to longer low flow seasons as soil moisture is reduced in the summer and autumn (Demaria and others, 2016).
Chattopadhyay and others (2017) used the Soil and Water Assessment Tool to quantify future changes in water yield and surface runoff in the Kentucky River Basin. Consistent with the other studies in the broader region, they predict decreasing water yield and increasing soil moisture deficit in the summer. Further work in Indiana using both the Variable Infiltration Capacity model and the Soil and Water Assessment Tool supports future decreases in runoff and increases in short sudden droughts in the summer as PET exceeds soil water availability (Cherkauer and others, 2021). Within the distribution of A. barbouri, warming in the summer (fig. 6) not offset by increasing precipitation (fig. 3) will likely lead to increasing soil water deficit and low flow. Multiple models across the distribution of A. barbouri indicate that the length of no-flow periods in breeding streams is likely to increase.
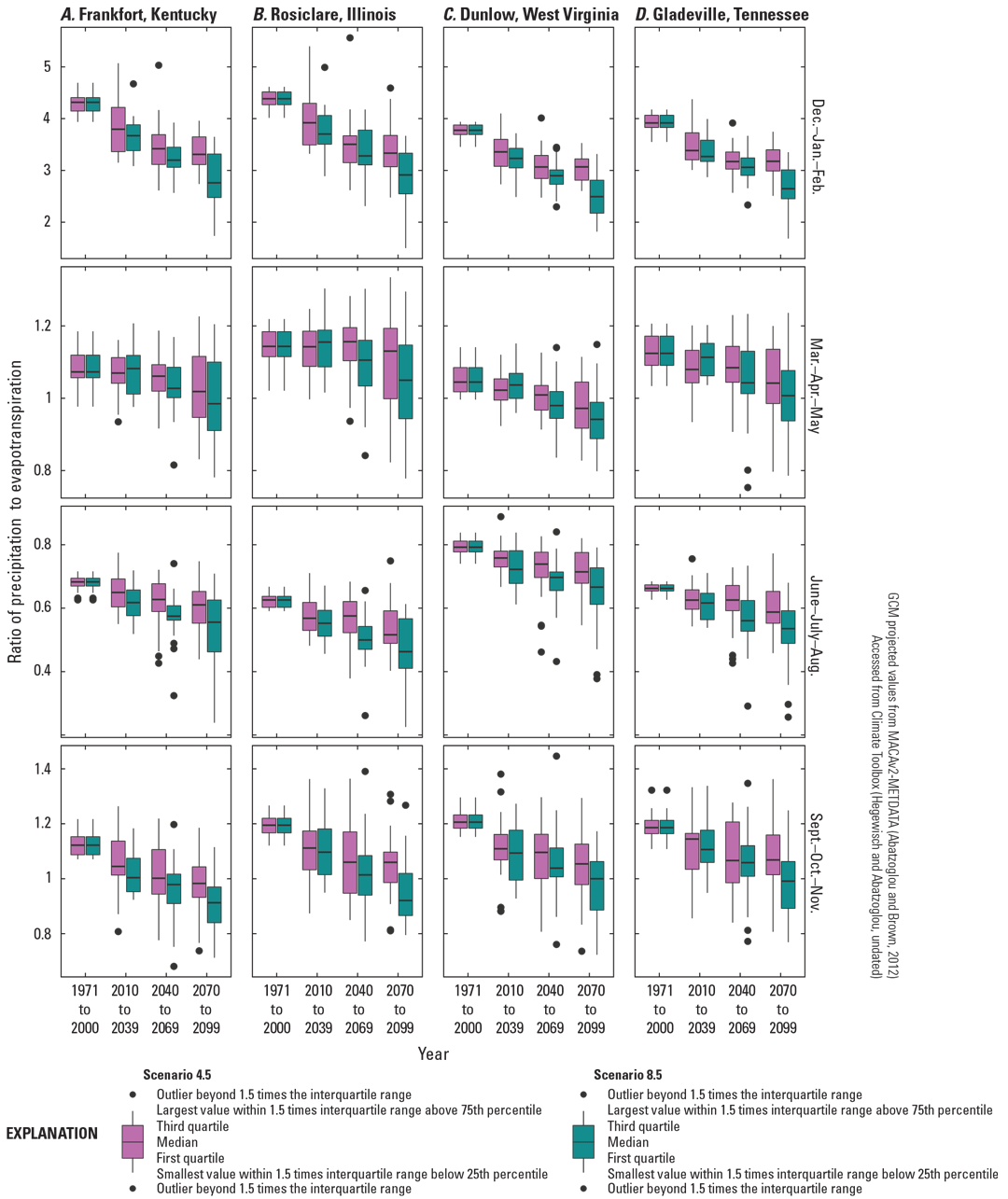
Projected seasonal ratio of precipitation volume to potential evapotranspiration for four 30-year periods from 1971 to 2099 in the distribution areas of Ambystoma barbouri. Projections centered on weather station points used for climate normals (figs. 1 and 2). A, Frankfort, Kentucky, the center of the core of A. barbouri’s distribution; B, Rosiclare, Illinois; C, Dunlow, West Virginia; and, D, Gladeville, Tennessee.
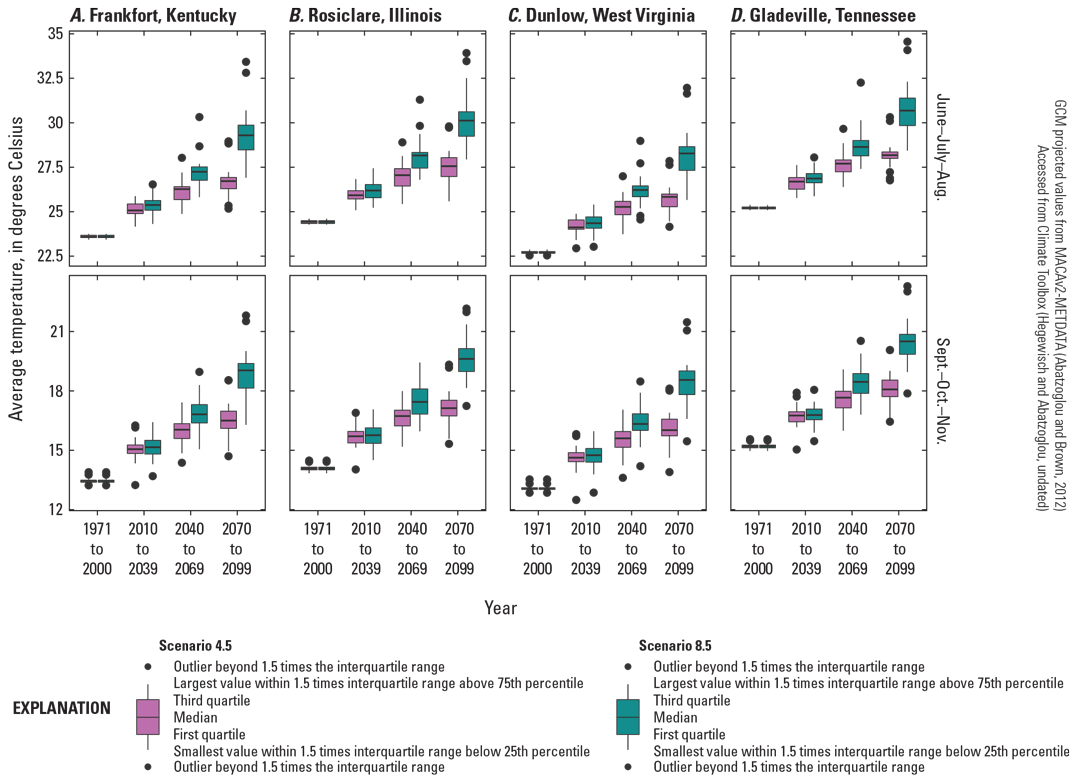
Projected seasonal average temperature for four 30-year periods from 1971 to 2099 in the distribution areas of Ambystoma barbouri. Projections centered on weather station points used for climate normals (figs. 1 and 2). A, Frankfort, Kentucky, the center of the core of A. barbouri’s distribution; B, Rosiclare, Illinois; C, Dunlow, West Virginia; and, D, Gladeville, Tennessee.
Reproduction and Recruitment
Adult A. barbouri are fossorial and secretive. Current available knowledge of this species’ ecology is primarily focused on periods of reproduction and larval development. During the egg and larval period, A. barbouri relies on specific aquatic habitats. As a result, these early life stages are likely very sensitive to changes in the environment.
Earlier Stream Dry Down Reduces Larval Survival
A. barbouri eggs and larvae require submergence for survival. Eggs hatch in 29–82 days; larvae take 6–10 weeks to reach metamorphosis (Petranka, 2010). The length of the incubation and larval period is related to environmental conditions experienced during development; higher temperatures foster faster development (Petranka, 1984b). Like other members of the genus, larvae of A. barbouri likely respond to environmental cues including temperature and stream depth. These responses can advance metamorphosis and help avoid desiccation for larvae in the stream (Semlitsch and Wilbur, 1988). Earlier metamorphosis in other Ambystoma species can reduce body size (McMenamin and Hadly, 2010) and fitness (Semlitsch and others, 1988). Larval survival of A. barbouri decreases with earlier dry down (Holomuzki, 1991). Low water levels increase the risk of desiccation and predation for larvae (that is, facilitating access for terrestrial predators like raccoons and birds; Petranka, 2010). This sensitivity to stream dry down may explain the western and southern range limits of A. barbouri based on evidence from a correlative niche model (Micheletti and Storfer, 2015) and a gene flow model (Micheletti and Storfer, 2017).
Future climate change will likely alter the timing and number of no-flow days in intermittent streams. Increased no-flow duration in intermittent streams may reduce larval survival if streams dry before metamorphosis. However, these effects may be buffered if warming temperatures shorten the egg and larval development period.
Survival
Like most biphasic salamanders, the early aquatic life stages of A. barbouri are likely the most sensitive to changes in the environment. Because of their low energetic demands and use of burrows, adult salamanders can behaviorally thermoregulate. Where it has been studied, adult salamanders only leave burrows during optimal conditions. Salamanders avoid unsuitable conditions by emerging for food acquisition and reproduction at night (Semlitsch and Pechmann, 1985) and when surface temperature is cool and moisture is high (McEntire and Maerz, 2019).
Increasing Temperature May Decrease Survival for Recently Metamorphosized Salamanders
Adult A. barbouri retreat into burrows to avoid desiccation; however, recently metamorphosized salamanders need to find or create burrows, leaving them exposed to the surface environment. In areas where landcover change limits burrow availability, juvenile Ambystoma desiccation-related mortality increases (Rothermel and Luhring, 2005; Rothermel and Semlitsch, 2006). Juvenile A. barbouri emerge from aquatic habitats and seek refuge during the late summer and early fall when temperatures are high and precipitation is low (fig. 2). With future increases in summer and fall temperature (fig. 6), juvenile mortality may increase in marginal habitats with low burrow availability; however, A. barbouri occupy agricultural habitats (Drayer and others, 2020) and may be less sensitive to open habitats than other Ambystoma that rely exclusively on forested habitat for terrestrial life stages.
Exposure to Agriculture Chemicals and Higher Temperatures Increases Desiccation Risk
While behavioral avoidance of water loss is critical to adult and juvenile A. barbouri survival, exposure to certain agricultural chemicals may disrupt these behaviors and reduce survival. A. barbouri frequently occupy catchments with agriculture (Drayer and others, 2020). Atrazine, an herbicide commonly applied in corn production, readily washes from agricultural land into waterbodies (Solomon and others, 1996). The distribution of A. barbouri overlaps areas of heavy atrazine use and atrazine groundwater concentrations (DeSimone and others, 2015). If exposed to atrazine during development, A. barbouri have increased desiccation risk as adults; this is a result of increased salamander activity at the expense of water saving behaviors like huddling to decrease surface area exposure (Rohr and Palmer, 2005). A. barbouri exhibit a decrease in water saving behavior even 8 months after atrazine exposure (Rohr and Palmer, 2005).
Future increases in heavy precipitation events may increase atrazine runoff into streams (Ryberg and others, 2020). This increased runoff may result in more A. barbouri atrazine exposure during egg and larval development. Climate change and atrazine exposure will likely increase the risk of desiccation and impair behavioral plasticity to conserve water, respectively; these changes are likely to further decrease body condition, reduce fitness, and increase mortality.
Biotic Interactions
Increased Streamflow can Lead Larvae to Drift into Fish-Occupied Pools
To avoid depredation of larvae and eggs, A. barbouri exhibits selective ovipositing in fishless streams (Drayer and others, 2020; Kats and Sih, 1992). In intermittent stream systems, heavy precipitation and the consequent high discharge can result in larval drift (Segev and Blaustein, 2014). Young larvae are particularly prone to drift, which can cause physical trauma and wash the larvae into stream reaches occupied by predatory fish (Petranka, 1984b). Sih and others (1992) determined that only 6–8 percent of larvae that drifted into pools occupied by fish survived. Using a correlative niche model, Micheletti and Storfer (2017) identified increased growing-season precipitation as a possible barrier to northward range expansion and hypothesized that this may be because of heavy-rainfall-induced larval drift.
Heavy precipitation (defined here as the amount of precipitation falling in the heaviest 1 percent of events) in the A. barbouri range has already increased in the last century and is projected to increase by more than 40 percent by the end of the 21st century under a high emissions scenario (RCP8.5; Reidmiller and others, 2018). During A. barbouri’s larval stage, increased heavy precipitation events will likely cause larval drift to suboptimal habitat and decreased larval survival.
Phenology
Uncertainty in the Effects of Climate Change on Breeding Onset
Adult A. barbouri start migrating to breeding streams as early as October and breed from December to April (Anderson and others, 2014; Petranka, 1984a). Though not directly studied in A. barbouri, breeding migrations of congeners often correspond to weather cues like temperature thresholds coinciding with rainfall events, likely reflecting a reliance on ephemeral waterbodies (Holomuzki, 1991). Once at the breeding sites, Petranka (1984a) determined that A. barbouri adults increased surface activity with rainfall. Unlike explosive pond breeding observed in other Ambystoma spp., breeding activity for A. barbouri does not appear to be closely tied to specific rainfall or temperature conditions (Petranka, 1984a). Breeding activity starts during the coldest part of the year (fig. 2); it is unlikely that warming temperatures alone will shift breeding timing earlier in the year. If A. barbouri breeding cues are related to specific temperature and precipitation interacting cues, the timing of breeding may shift, demonstrating adaptive capacity. Based on current understanding of A. barbouri ecology, it is unclear if this species will shift breeding timing, compensating for potential earlier stream dry down.
Habitat
Climate Change Intensifies Stressors in an Urban Environment
A. barbouri’s distribution overlaps the metropolitan areas of Nashville, Louisville, Lexington, and Cincinnati (fig. 1), inhabiting areas in and around these urban areas. In urban streams, A. barbouri are present at much lower densities compared streams in rural and natural areas (Drayer and others, 2020). Drayer and others (2020) determined that both occupancy and abundance of larval A. barbouri were significantly lower in urban environments compared to environments with more forest or agricultural land. Drayer and others (2020) estimated abundance of A. barbouri at 35.3 individuals per 10 meter reach transect in forested sites, 22.5 in agricultural sites, and only 0.66 in urban sites. Reduced salamander populations in urban areas may result from changes to hydrology and increased presence of environmental contaminants (Diaz and others, 2020). Roads in urban areas contribute not only to increased contaminants and habitat fragmentation but also mortality as adult salamanders cross roads in search of suitable breeding habitat (Niemiller and others, 2009). Increased urbanization has been linked with reduced larval survival and reduced adult colonization in other salamanders with similar life histories (Price and others, 2012). In the Drayer and others (2020) study, streams with higher sodium concentration had lower A. barbouri larval abundance likely from sodium disrupting osmoregulation. Urban areas often have high sodium concentrations from road salting on impervious surfaces (Kaushal and others, 2017).
A. barbouri populations in and around metropolitan areas are likely to face compound threats of climate change and urbanization (Niemiller and others, 2006). In urban areas, heavy rainfall, which is projected to increase in frequency and intensity, is less likely to be absorbed into soil due to prevalence of impervious surfaces before reaching waterways (Wu and others, 2013). Urban areas will therefore experience increased runoff, higher peak flows, and increased contaminant loads (Wu and others, 2013). Compared to scenarios for climate or landcover change alone, models indicate that scenarios with both climate and landcover change result in higher runoff (Barlage and others, 2002; Wu and others, 2013). Increased nutrient, pollutant, and sediment movement will make urban watersheds less suitable for A. barbouri (Drayer and others, 2020).
The effects of increased future dry periods on A. barbouri are also expected to be more severe in urban watersheds. Intermittent streams in watersheds with more impervious surfaces dry more rapidly (Hammond and others, 2021), which may exacerbate the effects of decreasing future ratio of precipitation to evapotranspiration (fig. 5). Shorter flow periods in urban intermittent streams may decrease availability of A. barbouri breeding habitat and decrease larval survival. Additionally, dry periods may increase stream salinity because of a lack of dilution (Mosley, 2015) and decrease salamander abundance (Drayer and others, 2020).
References Cited
Abatzoglou, J.T., and Brown, T.J., 2012, A comparison of statistical downscaling methods suited for wildfire applications: International Journal of Climatology, v. 32, no. 5, p. 772–780. [Also available at https://doi.org/10.1002/joc.2312.]
Abatzoglou, J.T., 2013, Development of gridded surface meteorological data for ecological applications and modelling: International Journal of Climatology, v. 33, no. 1, p. 121–131. [Also available at https://doi.org/10.1002/ joc.3413.]
Allen, R.G., Pereira, L.S., Raes, D., and Smith, M., 1998, Crop evapotranspiration—Guidelines for computing crop requirements: Rome, Italy, United Nations Food and Agricultural Organization, Irrigation and Drainage Paper No. 56. [Also available at http://www.fao.org/3/ x0490e/x0490e00.htm.]
Anderson, M.A., Campbell, J.R., Carey, A.N., Dodge, D.R., Johnston, R.A., Mattison, E.R., Seddon, R.J., Singer, N.L., and Miller, B.T., 2014, Population survey of the streamside salamander in the Nashville Basin of Tennessee: Southeastern Naturalist, v. 13, no. 1, p. 101–107. [Also available at https://doi.org/10.1656/058.013.0108.]
Barlage, M.J., Richards, P.L., Sousounis, P.J., and Brenner, A.J., 2002, Impacts of climate change and land use change on runoff from a Great Lakes watershed: Journal of Great Lakes Research, v. 28, no. 4, p. 568–582. [Also available at https://doi.org/10.1016/S0380-1330(02)70606-0.]
Chattopadhyay, S., Edwards, D.R., Yu, Y., and Hamidisepehr, A., 2017, An assessment of climate change impacts on future water availability and droughts in the Kentucky River Basin: Environmental Processes, v. 4, no. 3, p. 477–507. [Also available at https://doi.org/10.1007/s40710-017-0259-2.]
Cherkauer, K.A., Bowling, L.C., Byun, K., Chaubey, I., Chin, N., Ficklin, D.L., Hamlet, A.F., Kines, S.J., Lee, C.I., Neupane, R., Pignotti, G.W., Rahman, S., Singh, S., Femeena, P.V., and Williamson, T.N., 2021, Climate change impacts and strategies for adaptation for water resource management in Indiana: Climatic Change, v. 165, article no. 21, p. 1-20. [Also available at https://doi.org/10.1007/s10584-021-02979-4.]
Demaria, E.M.C., Palmer, R.N., and Roundy, J.K., 2016, Regional climate change projections of streamflow characteristics in the Northeast and Midwest U.S: Journal of Hydrology. Regional Studies, v. 5, p. 309–323. [Also available at https://doi.org/10.1016/j.ejrh.2015.11.007.]
DeSimone, L.A., McMahon, P.B., and Rosen, M.R., 2015, The quality of our Nation’s water—Water quality in principal aquifers of the United States, 1991–2010: U.S. Geological Survey Circular 1360, 150 p. [Also available at https://doi.org/10.3133/cir1360.]
Diaz, P.H., Orsak, E.L., Weckerly, F.W., Montagne, M.A., and Alvarez, D.A., 2020, Urban stream syndrome and contaminant uptake in salamanders of Central Texas: Journal of Fish and Wildlife Management, v. 11, no. 1, p. 287–299. [Also available at https://doi.org/10.3996/032018-JFWM-017.]
Drayer, A.N., Guzy, J.C., and Price, S.J., 2020, Factors influencing the occupancy and abundance of streamside salamander (Ambystoma barbouri) in Kentucky streams: Journal of Herpetology, v. 54, no. 3, p. 299–305. [Also available at https://doi.org/10.1670/19-015.]
Eng, K., Wolock, D.M., and Dettinger, M.D., 2016, Sensitivity of intermittent streams to climate variations in the USA: River Research and Applications, v. 32, no. 5, p. 885–895. [Also available at https://doi.org/10.1002/rra.2939.]
Hammond, J.C., Zimmer, M., Shanafield, M., Kaiser, K., Godsey, S.E., Mims, M.C., Zipper, S.C., Burrows, R.M., Kampf, S.K., Dodds, W., Jones, C.N., Krabbenhoft, C.A., Boersma, K.S., Datry, T., Olden, J.D., Allen, G.H., Price, A.N., Costigan, K., Hale, R., Ward, A.S., and Allen, D.C., 2021, Spatial patterns and drivers of nonperennial flow regimes in the contiguous United States: Geophysical Research Letters, v. 48, no. 2. https://doi.org/10.1029/2020GL090794.
Holomuzki, J.R., 1991, Macrohabitat effects on egg deposition and larval growth, survival, and instream dispersal in Ambystoma barbouri: Copeia, v. 1991, no. 3, p. 687. https://doi.org/10.2307/1446394.
Jeong, D.I., Sushama, L., and Naveed Khaliq, M., 2014, The role of temperature in drought projections over North America: Climatic Change, v. 127, no. 2, p. 289–303. [Also available at https://doi.org/10.1007/s10584-014-1248-3.]
Kats, L.B., and Sih, A., 1992, Oviposition site selection and avoidance of fish by streamside salamanders (Ambystoma barbouri): Copeia, v. 1992, no. 2, p. 468. https://doi.org/10.2307/1446206.
Kaushal, S.S., Duan, S., Doody, T.R., Haq, S., Smith, R.M., Newcomer Johnson, T.A., Newcomb, K.D., Gorman, J., Bowman, N., Mayer, P.M., Wood, K.L., Belt, K.T., and Stack, W.P., 2017, Human-accelerated weathering increases salinization, major ions, and alkalinization in fresh water across land use: Applied Geochemistry, v. 83, p. 121–135. [Also available at https://doi.org/10.1016/j.apgeochem.2017.02.006.]
McEntire, K.D., and Maerz, J.C., 2019, Integrating ecophysiological and agent-based models to simulate how behavior moderates salamander sensitivity to climate: Frontiers in Ecology and Evolution, v. 7, p. 22. [Also available at https://doi.org/10.3389/fevo.2019.00022.]
McMenamin, S.K., and Hadly, E.A., 2010, Developmental dynamics of Ambystoma tigrinum in a changing landscape: BMC Ecology, v. 10, article no. 10, p. 1–10. [Also available at https://doi.org/10.1186/1472-6785-10-10.]
Micheletti, S.J., and Storfer, A., 2015, A test of the central-marginal hypothesis using population genetics and ecological niche modelling in an endemic salamander (Ambystoma barbouri): Molecular Ecology, v. 24, no. 5, p. 967–979. [Also available at https://doi.org/10.1111/mec.13083.]
Micheletti, S.J., and Storfer, A., 2017, An approach for identifying cryptic barriers to gene flow that limit species’ geographic ranges: Molecular Ecology, v. 26, no. 2, p. 490–504. [Also available at https://doi.org/10.1111/mec.13939.]
Mosley, L.M., 2015, Drought impacts on the water quality of freshwater systems—Review and integration: Earth-Science Reviews, v. 140, p. 203–214. [Also available at https://doi.org/10.1016/j.earscirev.2014.11.010.]
Niemiller, M.L., Glorioso, B.M., Nicholas, C., Phillips, J., Rader, J., Reed, E., Sykes, K.L., Todd, J., Wyckoff, G.R., Young, E.L., and Miller, B.T., 2006, Status and distribution of the streamside salamander, Ambystoma barbouri, in middle Tennessee: American Midland Naturalist, v. 156, no. 2, p. 394–399. [Also available at https://doi.org/10.1674/0003-0031(2006)156[394:SADOTS]2.0.CO;2.]
Niemiller, M.L., Glorioso, B.M., Nicholas, C., Phillips, J., Rader, J., Reed, E., Sykes, K.L., Todd, J., Wyckoff, G.R., Young, E.L., and Miller, B.T., 2009, Notes on the reproduction of the streamside salamander, Ambystoma barbouri, from Rutherford County, Tennessee: Southeastern Naturalist, v. 8, no. 1, p. 37–44. [Also available at https://doi.org/10.1656/058.008.0104.]
Petranka, J.W., 1984a, Breeding migrations, breeding season, clutch size, and oviposition of stream-breeding Ambystoma texanum: Journal of Herpetology, v. 18, no. 2, p. 106–112. [Also available at https://doi.org/10.2307/1563737.]
Petranka, J.W., 1984b, Incubation, larval growth, and embryonic and larval survivorship of smallmouth salamanders (Ambystoma texanum) in streams: Copeia, v. 1984, no. 4, p. 862. [Also available at https://doi.org/10.2307/1445329.]
Price, S.J., Browne, R.A., and Dorcas, M.E., 2012, Resistance and resilience of a stream salamander to supraseasonal drought: Herpetologica, v. 68, no. 3, p. 312–323. [Also available at https://doi.org/10.1655/HERPETOLOGICA-D-11-00084.1.]
Reidmiller, D.R., Avery, C.W., Easterling, D.R., Kunkel, K.E., Lewis, K.L.M., Maycock, T.K., and Stewart, B.C., 2018, Fourth national climate assessment volume II—Impacts, risks, and adaptation in the United States: U.S. Global Change Research Program, accessed August 19, 2021, at https://nca2018.globalchange.gov/.
Rohr, J.R., and Palmer, B.D., 2005, Aquatic herbicide exposure increases salamander desiccation risk eight months later in a terrestrial environment: Environmental Toxicology and Chemistry, v. 24, no. 5, p. 1253–1258. [Also available at https://doi.org/10.1897/04-448R.1.]
Rothermel, B.B., and Luhring, T.M., 2005, Burrow availability and desiccation risk of mole salamanders (Ambystoma talpoideum) in harvested versus unharvested forest stands: Journal of Herpetology, v. 39, no. 4, p. 619–626. [Also available at https://doi.org/10.1670/251-04A.1.]
Rothermel, B.B., and Semlitsch, R.D., 2006, Consequences of forest fragmentation for juvenile survival in spotted (Ambystoma maculatum) and marbled (Ambystoma opacum) salamanders: Canadian Journal of Zoology, v. 84, no. 6, p. 797–807. [Also available at https://doi.org/10.1139/z06-056.]
Runkle, J., Kunkel, K.E., Champion, S.M., Frankson, R., and Stewart, B.C., 2022, Kentucky state climate summary 2022: National Oceanic and Atmospheric Administration Technical Report NESDIS 150–KY, 4 p., accessed January 2023 at https://statesummaries.ncics.org/downloads/Kentucky-StateClimateSummary2022.pdf.
Ryberg, K.R., Stone, W.W., and Baker, N.T., 2020, Causal factors for pesticide trends in streams of the United States—Atrazine and deethylatrazine: Journal of Environmental Quality, v. 49, no. 1, p. 152–162. [Also available at https://doi.org/10.1002/jeq2.20045.]
Segev, O., and Blaustein, L., 2014, Influence of water velocity and predation risk on fire salamander (Salamandra infraimmaculata) larval drift among temporary pools in ephemeral streams: Freshwater Science, v. 33, no. 3, p. 950–957. [Also available at https://doi.org/10.1086/676634.]
Semlitsch, R.D., and Pechmann, J.H.K., 1985, Diel pattern of migratory activity for several species of pond-breeding salamanders: Copeia, v. 1985, no. 1, p. 86. [Also available at https://doi.org/10.2307/1444794.]
Semlitsch, R.D., Scott, D.E., and Pechmann, J.H.K., 1988, Time and size at metamorphosis related to adult fitness in Ambystoma talpoideum: Ecology, v. 69, no. 1, p. 184–192. [Also available at https://doi.org/10.2307/1943173.]
Semlitsch, R.D., and Wilbur, H.M., 1988, Effects of pond drying time on metamorphosis and survival in the salamander Ambystoma talpoideum: Copeia, v. 1988, no. 4, p. 978. [Also available at https://doi.org/10.2307/1445721.]
Sih, A., Kats, L.B., and Moore, R.D., 1992, Effects of predatory sunfish on the density, drift, and refuge use of stream salamander larvae: Ecology, v. 73, no. 4, p. 1418–1430. [Also available at https://doi.org/10.2307/1940687.]
Solomon, K.R., Baker, D.B., Richards, R.P., Dixon, K.R., Klaine, S.J., La Point, T.W., Kendall, R.J., Weisskopf, C.P., Giddings, J.M., Giesy, J.P., Hall, L.W., Jr., and Williams, W.M., 1996, Ecological risk assessment of atrazine in North American surface waters: Environmental Toxicology and Chemistry, v. 15, no. 1, p. 31–76. [Also available at https://doi.org/10.1002/etc.5620150105.]
Storfer, A., Cross, J., Rush, V., and Caruso, J., 1999, Adaptive coloration and gene flow as a constraint to local adaptation in the streamside salamander, Ambystoma barbouri: Evolution; International Journal of Organic Evolution, v. 53, no. 3, p. 889–898. [Also available at https://doi.org/10.1111/j.1558-5646.1999.tb05383.x.]
Strong, C., McCabe, G.J., and Weech, A., 2020, Step increase in eastern U.S. precipitation linked to Indian Ocean warming: Geophysical Research Letters, v. 47, no. 17, 10 p. [Also available at https://doi.org/10.1029/2020GL088911.]
U.S. Geological Survey [USGS] Gap Analysis Project [GAP], 2018, U.S. Geological Survey—Gap Analysis Project species range maps CONUS_2001: U.S. Geological Survey data release, accessed August 2022 at https://doi.org/10.5066/F798865X.
Wang, G., Kirchhoff, C.J., Seth, A., Abatzoglou, J.T., Livneh, B., Pierce, D.W., Fomenko, L., and Ding, T., 2020, Projected changes of precipitation characteristics depend on downscaling method and training data—MACA versus LOCA using the U.S. Northeast as an example: Journal of Hydrometeorology, v. 21, no. 12, p. 2739–2758. [Also available at https://doi.org/10.1175/JHM-D-19-0275.1.]
Wootten, A.M., Dixon, K.W., Adams‐Smith, D.J., and McPherson, R.A., 2021, Statistically downscaled precipitation sensitivity to gridded observation data and downscaling technique: International Journal of Climatology, v. 41, no. 2, p. 980–1001. [Also available at https://doi.org/10.1002/joc.6716.]
Wu, J.Y., Thompson, J.R., Kolka, R.K., Franz, K.J., and Stewart, T.W., 2013, Using the Storm Water Management Model to predict urban headwater stream hydrological response to climate and land cover change: Hydrology and Earth System Sciences, v. 17, no. 12, p. 4743–4758. [Also available at https://doi.org/10.5194/hess-17-4743-2013.]
For more information about this publication, contact:
Director, USGS Midwest Climate Adaptation Science Center
1954 Buford Avenue
St. Paul, MN 55108
For additional information, visit: https://www.usgs.gov/programs/climate-adaptation-science-centers/midwest-casc
Publishing support provided by the Rolla Publishing Service Center
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
Lyons, M.P., LeDee, O.E., and Boyles, R., 2023, Potential effects of climate change on Ambystoma barbouri (streamside salamander): U.S. Geological Survey Open-File Report 2021–1104–C, 14 p., https://doi.org/10.3133/ofr20211104C.
ISSN: 2331-1258 (online)
Study Area
| Publication type | Report |
|---|---|
| Publication Subtype | USGS Numbered Series |
| Title | Potential effects of climate change on Ambystoma barbouri (streamside salamander) |
| Series title | Open-File Report |
| Series number | 2021-1104 |
| Chapter | C |
| DOI | 10.3133/ofr20211104C |
| Publication Date | April 05, 2023 |
| Year Published | 2023 |
| Language | English |
| Publisher | U.S. Geological Survey |
| Publisher location | Reston, VA |
| Contributing office(s) | Midwest Climate Adaptation Science Center |
| Description | Report: vi, 14 p.; Data Release |
| Country | United States |
| State | Illinois, Indiana, Kentucky, Ohio, Tennessee, West Virginia |
| Online Only (Y/N) | Y |