

Belowground Mutualisms to Support Prairie Reconstruction—Improving Prairie Habitat Using Mycorrhizal Inoculum
Links
- Document: Report (1.18 MB pdf) , HTML , XML
- Data Releases:
- USGS data release - Management of remnant tallgrass prairie by grazing or fire in western Minnesota, 2016–2017
- USGS data release - Arbuscular mycorrhizal fungi in remnant and reconstructed prairies in Minnesota and Iowa, 2019 (ver. 2.0, April 2022)
- Download citation as: RIS | Dublin Core
Acknowledgments
We are grateful to the following cooperators for permitting access to study sites: U.S. Fish and Wildlife Service: Morris, Litchfield, and Fergus Falls Wetland Management Districts and Neal Smith National Wildlife Refuge; Minnesota Department of Natural Resources: Clinton Prairie Scientific and Natural Area and Pednor Wildlife Management Area; and to the owner: Maminka Prairie.
Abstract
As a first step toward understanding the feasibility of using arbuscular mycorrhizal fungi (AMF) in reconstruction practice, we addressed four objectives: (1) compare root-associated AMF communities of plants between high-quality remnant prairies and reconstructed prairies, (2) compare root-associated AMF communities between plant species that declined in reconstructions and species that were thriving, (3) compare AMF communities collected from roots of plants in geographically separate parts of Minnesota and Iowa, and (4) assess the relationship between AMF communities and soil abiotic factors. We collected soil and root samples in 8 prairies reconstructed in 2005 (and monitored through 2015) and 6 remnant prairies, and the samples were separated into 6 geographically determined clusters, each containing 1–2 reconstructions and 1 remnant. Sequencing was completed on 1,188 deoxyribonucleic acid extracts from individual plant root samples, and fungal sequences were clustered to operational taxonomic units at 97-percent identity. Nonmetric multidimensional scaling was used to visualize differences in species composition of AMF communities among plant species and field sites. Permutational analysis of variance was completed to test for differences in AMF community composition between the 2 types of sites (remnants and reconstructions), among plant species, and among the 6 site clusters. AMF communities differed between remnant and reconstructed prairies, with one exception, and AMF associated with individual plant species also tended to differ, depending on whether the plant species’ roots were collected from remnant or reconstructed prairie. On the other hand, we did not determine that, as a group, species in decline in the reconstructions we had monitored were more likely to harbor different AMF communities compared to species not in decline in the reconstructions. Significant interactions between site type and clusters indicate geographic variation in AMF communities. Total carbon and nitrogen, and organic matter, were higher in remnant soils, whereas phosphorus, which at high concentrations reduces the value of AMF to plants, was much higher in soils collected from reconstructions.
Introduction
Prairie reconstruction is one of the few remaining options to increase connectivity among existing fragments of remnant prairie, buffer remnants from disturbance and pesticide drift, and increase native prairie abundance (Gerla and others, 2012), which is critically important to conservation of pollinators and other native prairie biota. Reconstructed prairies also are increasingly viewed as highly valuable to mainstream production agriculture in the Midwest because they offer habitat for pollinators and other beneficial insects that provide valuable services in agroecosystems, such as control of weeds and pest arthropods (Schulte and others, 2017), and highly land-efficient conservation of soil and water. Thus, from the perspectives of nature preservation and agroecology, reconstructions can be of critical importance; however, the biodiversity conservation value of prairie reconstructions depends strongly on their species richness (Nicolson and Wright, 2017). Unfortunately, reconstruction practitioners struggle to reproduce the species richness and ecosystem services provided by large remnant prairies (Larson and others, 2018). An experimental study of eight reconstructed prairies in Minnesota and Iowa determined that one-half of the planted species declined or failed to establish (Larson and others, 2017). More reliable and cost-effective prairie reconstruction methods are needed. Calls to increase habitat for struggling Danaus plexippus (Linnaeus, 1758; monarch butterflies) and declining pollinators have further emphasized the need for improved reconstruction methods.
Arbuscular mycorrhizal fungi (AMF) may play an important role in improving the efficiency of reconstruction methods. AMF are root-associated symbionts of many prairie plant species and have been documented repeatedly to have strongly mutualistic relationships with such species (Middleton and Bever, 2012). In particular, AMF can assist in the development of species-rich native prairie reconstructions, especially if late-successional species are desired (Middleton and Bever, 2012), some of which seem to have obligate relationships with AMF taxa (Koziol and Bever, 2017). In addition, AMF have been determined to increase pollinator visitation to a flowering plant by increasing general robustness of growth and flower production (Wolfe and others, 2005). Unfortunately, intensive land uses such as agriculture that precede reconstruction commonly leave soils in a degraded state (McLauchlan, 2006). AMF in particular have been determined to have different, and depauperate, community composition and lower abundance in cropland than in grassland (Oehl and others, 2003). Degraded soil conditions are commonly highly stable (Suding and others, 2004), and thus, substantial effort may be required to restore key soil attributes for achieving diversity-related reconstruction goals, such as plant-soil mutualisms with native plant species (Asmelash and others, 2016).
Active inoculation of AMF can be useful for establishment of native prairie plants, but outcomes of such inoculation are complex and contingent on many biotic and abiotic variables. Commercial AMF inoculant does not consistently improve plant establishment and growth (Maltz and Treseder, 2015). The primary role of AMF in mutualistic relationships with prairie plants is in enhancing water and nutrient uptake in exchange for sugars; however, the relationship is not always mutually beneficial. Under some conditions, AMF may act parasitically to obtain carbon resources without providing substantial benefit to the plant. This parasitism may, in part, explain the unreliability of commercial AMF inoculants as tools for restoration (Maltz and Treseder, 2015). Moreover, Koziol and Bever (2016) determined that specificity of the AMF-plant mutualism was especially pronounced for late-successional plant species, species that are typically the most difficult to establish and maintain in prairie reconstructions.
The method by which AMF are introduced into a reconstruction also is an important consideration. A metaanalysis that included 28 manipulative field-based trials, two of which were on temperate grasslands, indicated that inoculum from a reference site was as effective as inoculum cultured from individual species, and both were superior to commercial inoculum (Maltz and Treseder, 2015). Locally sourced inoculum also may have the advantage of including AMF taxa that are adapted to local conditions; AMF ecotypes are not necessarily interchangeable in their functioning (Sanders and Rodriguez, 2016). Maltz and Treseder (2015) also cautioned that sourcing inoculum from remnants should be carefully planned so that the remnant is not damaged in the process.
In this study, we evaluated the difference in AMF taxa colonizing roots of plant species collected in high-quality remnant versus reconstructed prairies. Objectives of the study were to complete the following:
-
1. Compare AMF communities collected from roots of plants in high-quality remnant prairies with AMF communities in reconstructed prairies; differences will indicate that the addition of AMF associated with high-quality remnants may have the potential to improve reconstruction outcomes.
-
2. Compare AMF communities collected from roots of plant species that declined in planted reconstructions with AMF detected in species that were thriving; differences will indicate that improving AMF available to poorly performing plant species has the potential to increase their establishment, growth, or persistence in reconstructed prairies.
-
3. Compare AMF communities collected from roots of plants in geographically separate parts of Minnesota and Iowa; geographic differences will indicate that AMF communities are spatially distinct and will require localized sourcing if AMF are to be added to reconstructions.
-
4. Assess the relationship between AMF communities and soil abiotic factors (for example, nitrogen, phosphorus, micronutrients, and soil texture); if soil abiotic factors affect AMF communities, these factors may require remediation before AMF communities more typical of remnants can establish.
Methods
Eight reconstruction sites on National Wildlife Refuge (NWR) system lands in Iowa and Minnesota were selected (fig. 1). The sites were part of a long-term study of effects of seed mix richness and planting method on establishment success of prairie reconstructions (Larson and others, 2011, 2017). Reconstructions were established in 2005, and cover of native and invasive plant species was evaluated in 2005–7, 2010, and 2015 to determine which species seeded into the reconstructions had increased or declined in abundance over time. No AMF amendments were used in these reconstructions.
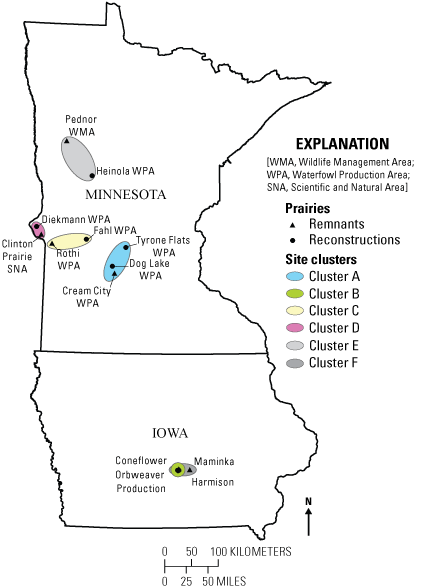
Locations of 8 reconstructed and 6 remnant prairies used in this study. In Iowa, three reconstruction sites and the Coneflower remnant are at Neal Smith National Wildlife Refuge. The Maminka remnant is about 20 kilometers east of the refuge. Colored ovals contain site clusters; that is, reconstructions and remnants selected based on similar characteristics.
Remnant Prairies
A two-tiered approach was used to select remnant prairies for our study. For sites in the Morris Wetland Management District, vegetation data from surveys done during a previous study (Larson and others, 2020) were used to identify prairies with the highest native species richness, a soil drainage class similar to our reconstructions, and a condition ranking of at least good to fair (or BC) for native plant communities under the Minnesota Department of Natural Resources ranking system (Minnesota Biological Survey, 2014). We acknowledge that these sites, although the best we could find, still harbored invasive grasses and sweetclover. For all other Minnesota sites, a search was completed using the Minnesota Department of Natural Resources Native Prairies shapefile (https://gisdata.mn.gov/), which limited acceptable remnant prairies to those with a good to fair ranking or better. Because no remnant prairie sites were within these parameters in the central and north-central parts of the State, we paired one reconstruction in that region with Pednor Wildlife Management Area (fig. 1). Pednor Wildlife Management Area was 80 kilometers (km) from Heinola Waterfowl Production Area, which was 16 km farther than our next most-distant remnant-reconstruction pair. This remnant was chosen based on previous knowledge of the site by one of our coauthors (Laura Aldrich-Wolfe). In Iowa, no equivalent database of native prairie sites is in the public domain. Based on recommendations of staff at Neal Smith NWR, we chose a high-quality remnant prairie site at Neal Smith NWR (Coneflower site on fig. 1) and Maminka Prairie, a privately owned remnant about 20 km east of the refuge (fig. 1).
Field Sampling
Plants were sampled from the 8 reconstructed and 6 nearby high-quality remnant prairie sites in Minnesota and Iowa (a total of 14 sites; table 1). At each site, parts of the root system of at least 5 plants (except Liatris pycnostachya Michx. with only 3 samples total) of each of our target species were excavated and placed in a labeled coin envelope in a plastic bag containing silica gel desiccant packets. Within 12 hours of sampling, roots were dried for 24 hours in an oven at 35 degrees Celsius and subsequently stored with silica gel.
Table 1.
Site names and their locations (State, latitude, and longitude), cluster affiliation (see fig. 1), date when roots were collected, and site type (remnant or reconstruction).[WPA, Waterfowl Production Area; MN, Minnesota; IA, Iowa; SNA, Scientific and Natural Area; WMA, Wildlife Management Area]
At a single site, 4 plant species were sampled (appendix 1, tables 1.1 and 1.2; Dalea purpurea Vent., Liatris ligulistylis [A. Nelson] K. Schum., Symphyotrichum pilosum [Willd.] G.L. Nesom, and Symphyotrichum Nees species), and at 2 reconstructions, 2 plant species were sampled (Monarda fistulosa L. and Symphyotrichum novae-angliae [L.] G.L. Nesom). The remaining 15 species were sampled at reconstructions and remnants, although not necessarily at both types of sites within a cluster.
Ten 2- x 10-centimeter soil cores were collected haphazardly throughout the area in which roots were sampled. Cores were composited, air-dried at room temperature, and stored in paper bags. Soil collected at each study site was analyzed for ammonium, phosphorus and potassium, total carbon and total nitrogen, texture, micronutrients, and pH at the University of Minnesota Research Analytical Laboratory (https://ral.cfans.umn.edu/).
Deoxyribonucleic Acid Extraction and Fungal Sequencing
Five 1-centimeter-long segments of root were ground using two 3-millimeter metal beads (steel shot no. 4, Ballistic Products Inc., Corcoran, Minnesota) on a TissueLyzer (Qiagen, Valencia, California) at 30 hertz for 1.5 minutes. Deoxyribonucleic acid (DNA) was extracted from each root sample using the DNeasy Plant Mini Kit (Qiagen, Valencia, Calif.) following the manufacturer’s protocol. DNA extracts were sent to the University of Minnesota Genomics Center (St. Paul, Minn.) for internal transcribed spacer region ITS2 bidirectional sequencing after amplification with universal fungal primers 5.8SR and ITS4. In every extraction set (1 of 24 samples), 1 water blank (negative controls for DNA extractions) was included to allow for detection of contaminating sequences; 12 water blanks were included for sequencing. Sequencing was completed on 1,188 DNA extracts from individual plant root samples in 6 Illumina MiSeq 2 x 300 base pair lanes at the University of Minnesota Genomics Center.
Bioinformatics
After initial quality control at the University of Minnesota Genomics Center to remove poor-quality sequences, sequences were processed using the PIPITS pipeline (version 2.7, Gweon and others, 2015) at the University of Groningen Peregrine High Performance Computing Cluster (Groningen, Netherlands). Forward and reverse reads from each sample were paired, and low-quality bases (Phred score of less than 30) and reads with ambiguous bases were removed. The fungal ITS2 region was extracted using ITSx software (Bengtsson-Palme and others, 2013). Residual primer sequences in any orientation were detected using the Biostrings (version 2.54.0; Pagès and others, 2019) and DADA2 (version 1.14.1; Callahan and others, 2016) packages in R (version 3.6.3; R Core Team, 2020) and removed using Cutadapt (version 2.10; Martin, 2011) in Peregrine. Using the PIPITS pipeline, short reads (less than 100 base pairs) and singletons, which are likely to be sequencing artifacts, were removed. Sequences were clustered using the VSEARCH centroid clustering methods (default for PIPITS) to operational taxonomic units (OTUs) at 97-percent identity, and chimeric sequences were identified and removed. We used the ITS DADA2 workflow, as described in Callahan (2022), under the heading “Identify Primers.”
These quality filtered reads were used for taxonomic assignment with the RDP Classifier against the UNITE database (version 04.02.2020; Kõljalg and others, 2013) at a 0.7-confidence level using the PIPITS pipeline. Sequence counts for any OTU observed in the negative controls were combined and subtracted from counts of the samples; any negative values were set to zero. The resulting 14,218 fungal OTUs in 1,134 samples (54 of the original samples no longer contained any sequences) were used for further processing and statistical analysis in R.
To reduce the number of zeros in the final AMF sequence dataset (which hinders ordination analysis), sequences were summed within each plant species at each site, reducing the number of samples to 132; this merged dataset was used in the statistical analysis of AMF communities. Before analysis, abundances were converted to relative abundances using the microbiome package (Lahti and others, 2017), and a subset was created to include only AMF taxa (filtered by phylum Glomeromycota) using the phyloseq package (version 1.30.0; McMurdie and Holmes, 2013); of the 14,218 fungal OTUs detected, 651 were identified as AMF. To reduce the disproportional effect of rare AMF taxa on assessment of overall communities, OTUs that were in fewer than 3 of the 132 samples and had a relative abundance less than 1×10−5 were removed from the dataset, with 396 OTUs and 131 samples remaining.
Statistical Analyses
Nonmetric multidimensional scaling (NMDS) using a Bray-Curtis dissimilarity matrix in phyloseq was used to visualize differences in species composition of AMF communities among plant species and field sites. Samples from all plant species were included in this analysis. Permutation analysis of variance was completed in phyloseq to test for differences in AMF community composition between the 2 types of sites (remnants and reconstructions), among plant species, among the 6 site clusters, and including all interactions. The betadisper function of the vegan package (version 2.5–6; Oksanen and others, 2020) was used to assess homogeneity of group dispersion between remnants and reconstructions. We also completed an NMDS analysis using only the plant species that were in both remnant and reconstructed sites to determine if those plants that were in only one site type affected the overall results.
The envfit function of the vegan package (version 2.5–6; Oksanen and others, 2020) was used to test for correlations between environmental variables and AMF community composition. The preliminary model included all environmental variables (Larson and others, 2021 [“2019 prairie remnant and reconstruction soil properties data v1.1” file]), whereas the final model excluded those environmental variables for which the relationship to AMF community composition was not statistically significant at α=0.05, where α is the significance level. To assess if AMF community composition correlated with changes in plant frequency, we completed an envfit analysis using a subset of plants whose abundance was recorded in six of the reconstruction sites in 2010 and 2015 (Larson and others, 2017).
Results
AMF communities in sampled plant roots differed between prairie site type (that is, reconstruction or remnant; table 2, fig. 2A), and with a few exceptions, this pattern was consistent. Nonetheless, interactions also were detected between site type and plant species (fig. 2B) and between site type and site cluster (fig. 2C). AMF communities in reconstructions were more similar to each other than AMF communities in remnants (fig. 2A for all sampled species, fig. 1.1 for only those species that were in both remnant and reconstructed sites; results were qualitatively similar). This finding was confirmed with a beta dispersion test, which indicated dispersion was significantly different between remnants and reconstructions (F=7.82, DF=1, and P=0.008 with 999 permutations, where F is the test statistic, DF is degrees of freedom, and P is significance) with remnants indicating a higher mean distance to their median (that is, were more dispersed) than the reconstructions (0.65 versus 0.63). For most plant species, AMF communities differed between remnant and reconstruction (table 2, fig. 2B); however, it was not uncommon to find one site in which remnant AMF communities were more similar to reconstruction communities for an individual plant species (for example, Andropogon gerardii [andger], Heliopsis helianthoides [helhel]).
Table 2.
Effects of site type, site cluster, and plant species on arbuscular mycorrhizal fungi community composition by three-way permutational analysis of variance.[Bold significance values (P) are significant at P less than (<) 0.05; ***, P<0.001; **, P<0.01; *, P<0.05; DF, degrees of freedom; SS, sum of squares; MS, mean square; F, test statistic; R2, variance accounted for by the ordination; --, no data or not applicable]
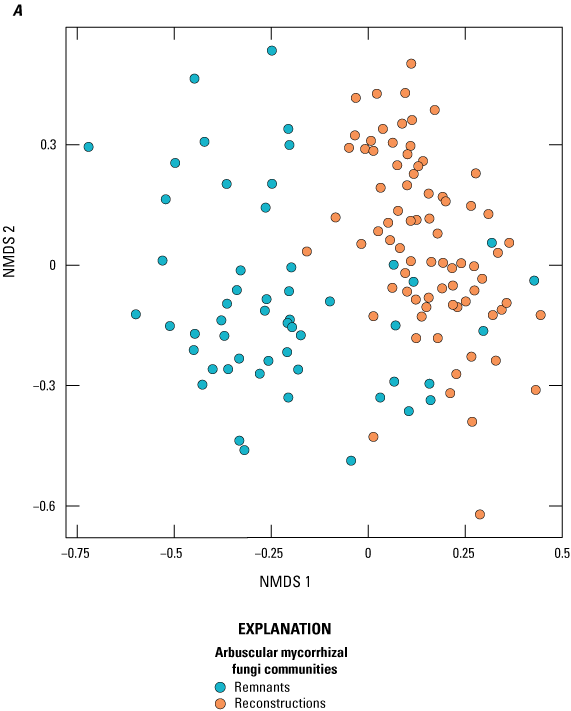
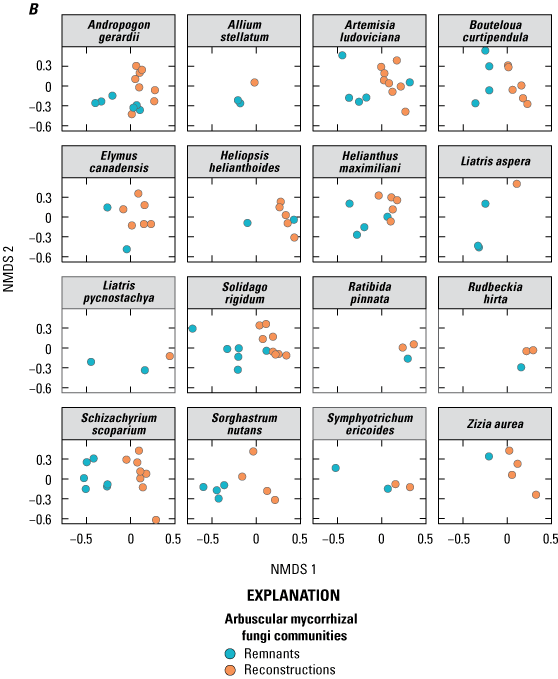
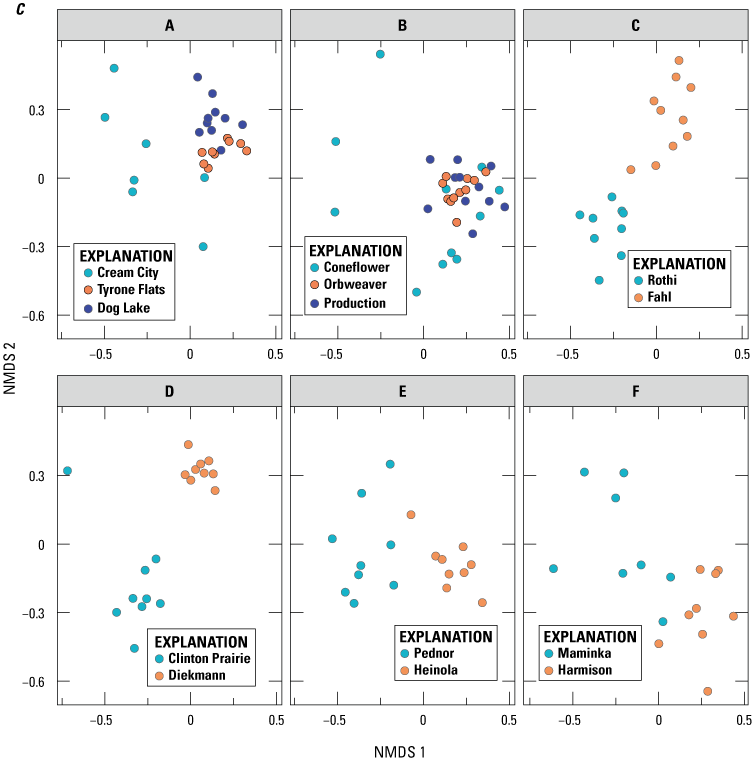
Arbuscular mycorrhizal fungi (AMF) communities in sampled plant roots plotted using nonmetric multidimensional scaling (NMDS) on Bray-Curtis distance matrix of relative abundance data. Points that are closer together represent communities that are more similar. The axes and orientation of the plots are arbitrary. (A) AMF communities in roots of plant species at 14 prairie sites, 6 of which were remnants and 8 of which were reconstructions. Each point represents the AMF community of a single plant species at a single site. (B) Plant species graphed independently to illustrate differences between reconstructions and remnants. Only plant species that had at least one representative from a remnant and a reconstruction site are shown. (C) Clusters (A–F) graphed independently to illustrate differences in AMF communities between reconstructions and remnants within site clusters.
The separation between AMF communities associated with remnants and reconstructions was consistent for most plant species at most site clusters (figs. 2C, 3). Overall, AMF communities differed along axis 1 of the NMDS, with remnant AMF communities on the left-hand side and reconstruction AMF communities on the right-hand side of the graph. The exceptions to this pattern of separation were site cluster A in Minnesota and B in Iowa, where the AMF communities of the remnant site were on the right-hand side of axis 1, indicating that they were similar to those of reconstruction sites (both in that cluster and the other reconstruction sites at the other clusters). In cluster A, only a few of the plant species were similar to species of the reconstruction sites, whereas in cluster B, almost all the plant species of the remnant (Coneflower) harbored AMF communities that resembled those of reconstructions. In none of the site clusters did the reverse happen; no reconstruction sites had AMF communities that resembled those of most of the remnants (that is, were placed on the left-hand side of axis 1).
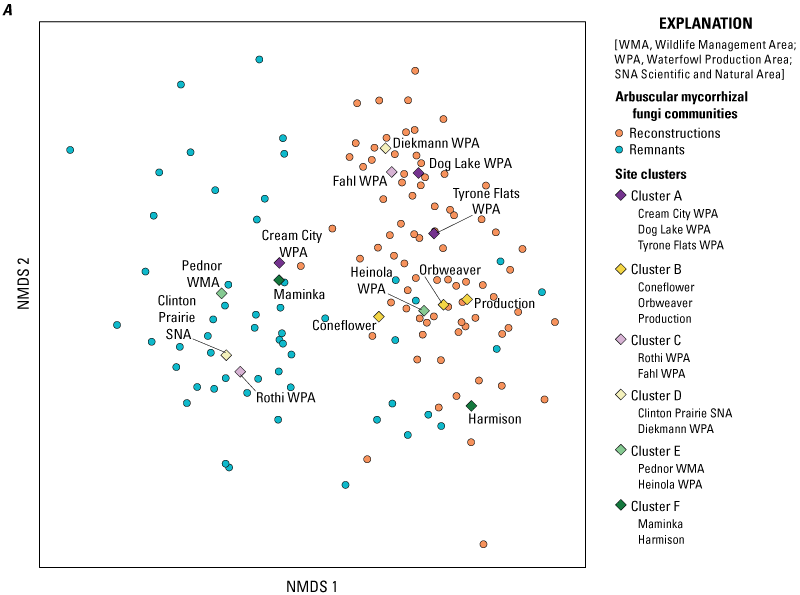
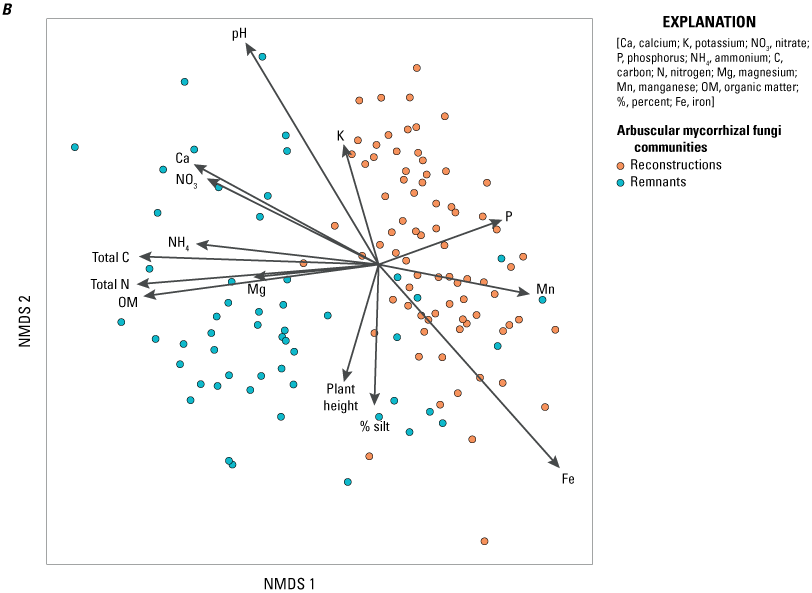
Relationship between arbuscular mycorrhizal fungi communities of samples from remnants or reconstructions and (A) sites (colored to site cluster) or (B) edaphic soil and plant characteristics (arrows). Only edaphic soil and plant characteristics that were significantly correlated (significance of less than 0.05) with arbuscular mycorrhizal fungi communities (as determined by the envfit analysis; table 3) are shown.
Contrary to our expectation, AMF community composition was not significantly related to declining or increasing species (determined in our monitoring study; table 3). The clear differences in AMF communities between remnants and reconstructions could be explained to a degree by edaphic soil characteristics (fig. 3B, table 3, appendix 1). The AMF communities of prairie remnants correlated in particular with higher organic matter, total carbon, and total nitrogen, and to a lesser degree with higher levels of ammonium, nitrate, magnesium, and calcium in the soil. AMF communities in reconstructions were particularly correlated with higher phosphorus and manganese levels. The separation between the sites within reconstructions and remnants was strongly related to iron and pH, and to a lesser degree with soil potassium and silt levels, and mean plant height at each site. Interestingly, the Coneflower remnant site had levels of manganese, iron, and phosphorus that were more in line with those of reconstruction sites (Larson and others, 2021 [“2019 prairie remnant and reconstruction soil properties data v1.1” file]).
Table 3.
Results of reduced model for evaluation of the relationship between environmental variables and nonmetric multidimensional scaling ordination, after removal of environmental variables that were not associated with arbuscular mycorrhizal fungi community composition at α=0.05 (where α is significance). For each variable, association with each of the two nonmetric multidimensional scaling axes, the variance in that variable accounted for by the ordination, and the significance of the variance accounted for by the ordination are listed.[NMDS 1, nonmetric multidimensional scaling axis 1; NMDS 2, nonmetric multidimensional scaling axis 2; R2, variance accounted for by the ordination; P, significance; **, P less than (<) 0.01; ***, P<0.001; %, percent; --, no data or not applicable; n.s., not significant]
Discussion
Overall, we determined that AMF communities differed between remnant and reconstructed prairies and that AMF associated with individual plant species also tended to differ, depending on whether the plant species’ roots were collected from remnant or reconstructed prairie. On the other hand, we did not determine that, as a group, plant species in decline in the reconstructions we monitored (Larson and others, 2017) were more likely to harbor different AMF communities compared to other species, perhaps owing to the generally different AMF communities in the two site types: nearly all the species demonstrated these differences. Significant interactions between site type and clusters indicate geographic variation in AMF communities, making the consistent differences between remnants and reconstructions along axis 1 of the NMDS even more striking.
Asmelash and others (2016) reviewed the ways in which AMF can be used to improve ecological restoration of degraded lands. They emphasized the lack of host specificity among AMF because of the relatively small number of AMF taxa compared to the number of plant species that form interactions with them (but see Hausmann and Hawkes [2010] and Werner and Kiers [2015]). Nonetheless, benefits of the mutualism vary among partners (Bever, 2002), over seasons (Bittebiere and others, 2020), and in differing environmental contexts (Revillini and others, 2016). Fungal taxa have been determined to vary in their “carbon sink strength,” such that some taxa are more beneficial for early development of individual plants (requiring little carbon from their hosts) and others for increasing biomass of larger plants that are able to supply more carbon (Martignoni and others, 2021). In the context of prairie reconstruction, an understanding of carbon sink strength, how it varies depending on AMF and host identity, and how to manipulate it to the advantage of the reconstruction may be an important aspect of characterizing functionally and ecologically significant differences in AMF taxa. Taxa beneficial to mature plants may not benefit seedlings. If taxa that benefit mature plants do not benefit seedlings, a single inoculation may not be sufficient to provide benefits over the life of a reconstruction, especially if AMF are unable to disperse naturally to a reconstruction site. All of our samples were collected from roots of mature plants; future work could evaluate effects of AMF collected in this way on seedling establishment and growth.
Only three of the soil characteristics we measured were clearly related to site type, with mean total nitrogen and carbon (and therefore also organic matter) about twice as high in remnants and mean phosphorus nearly six times higher in reconstructions, possibly as a legacy of farming. Excessive phosphorus, in particular, reduces the value of AMF to plants (Revillini and others, 2016). These measures (and several others) also were significantly related to AMF communities in these site types; however, it should be noted that levels of most environmental characteristics varied considerably at the site level, indicating a more complicated relationship with AMF taxa. This relationship is demonstrated by the remnant site in cluster B (Coneflower) whose AMF communities strongly resembled those of reconstructions. At this site, phosphorus, manganese, and iron were elevated relative to the mean levels for reconstructions; however, several reconstruction sites had low levels of the same elements, yet their AMF communities did not resemble those of remnants. The implications of such variation, especially with respect to prairie reconstruction, may not only relate to soil characteristics, but also relate to local adaptation of plants and AMF (Jack and others, 2021), as revealed by a “home and away” study that demonstrated coadaptation of plant genotypes and AMF communities in nutrient limited sites (Johnson and others, 2010). Likewise, the wider soil microbial community can influence the effect of AMF colonization on plant growth (Hetrick and others, 1988, 1990).
Studies in Estonian forest ecosystems have determined that generalist plant species associate with generalist AMF taxa, whereas habitat specialist plants may associate with generalist and specialist AMF (Davison and others, 2011). We determined greater dispersion of AMF communities in remnant prairies than in reconstructed prairies, perhaps reflecting greater abundance of specialist plants in remnants, each interacting with a few specialist AMF taxa. A global-scale metaanalysis of AMF communities associated with plant functional groups (cool-season and warm-season grasses, forbs, and legumes) determined that grasses tended to associate with a greater breadth of AMF taxa than did forbs (Davison and others, 2020); Tipton and others (2019) likewise determined the warm-season grass, Schizachyrium scoparium, to be a generalist with respect to AMF taxa. On average, 10 years after planting, 35–40 percent of total live cover consisted of warm-season grasses and 5–27 percent of total live cover consisted of forbs at the Iowa reconstructions (Drobney and others, 2020) we sampled for this study, but we do not have corresponding plant community data for the remnants. Although inspection of the graphs of AMF communities for individual species does not indicate widespread variation between warm-season grasses and forbs at our study sites, the relative abundance of the grasses compared to forbs may account for some of the overall variation in AMF communities.
We did not find a clear signal in AMF community composition to explain declining frequency or poor establishment in some of the plant species we had monitored in the reconstructions (Larson and others, 2017). Given the myriad circumstances in which plant species and AMF may encounter each other, and the importance of environmental conditions in shaping the interaction, an observational study such as ours may lack sufficient power to detect such effects. Greater benefit is commonly observed for late-successional plants (Middleton and Bever, 2012; Koziol and others, 2021; Cheeke and others, 2022), but the plants we observed to decline do not fall neatly into this category. Other members of the microbial community, such as rhizobia (Bauer and others, 2012), also may play a role in successful establishment of some species. As noted previously, the dominant difference demonstrated in the NMDS was between remnants and reconstructions, so differences between declining and increasing species may have been masked by the much larger magnitude of the effects of site type. In addition, we did not evaluate the contribution of individual AMF taxa, which may have had important associations for which we did not account.
Soil disturbance, such as by plowing, has been determined to change the composition and function of AMF in grasslands (Schnoor and others, 2011). More intensive land management was observed to result in a similar number of AMF species but different species composition (Börstler and others, 2006). Our results support these observations in that roots from plants in reconstructions, where the soil ecosystem had been subjected to disturbance for row-crop agriculture, had a different AMF community composition than did those in remnants, where soil had never been plowed. Dispersal of AMF is poorly understood (Paz and others, 2021), so the likelihood of their unassisted arrival in new reconstructions is unclear. Oehl and others (2003) determined declining numbers and diversity of AMF in a disturbance gradient ranging from lightly managed grassland to permanent row-crop agriculture. It must be kept in mind, however, that grasslands are disturbance-dependent ecosystems that cannot persist without fire, grazing, or their anthropogenic analogs (Perkins and others, 2019). We have no data on the AMF taxa present before reconstruction, but if the sites were as depauperate as the row-crop sites in the study completed by Oehl and others (2003), our sites experienced substantial colonization by AMF in the decade and a half since reconstruction began. Moreover, these reconstructions were all established by seed, so the planting itself was unlikely to introduce AMF.
The geographic variation we saw in AMF community composition, combined with the differences between reconstructed and remnant prairies, indicates the possibility of short-distance dispersal of AMF, perhaps in a random fashion within the area of an agricultural field, with selection over time by plant species acting as filters that winnow out AMF that provide fewer benefits (Davison and others, 2011). In this regard, the similarity between reconstruction and remnant sites in cluster B in Iowa may be instructive. Although the site labeled a remnant in cluster B may not have been plowed, one of us (P. Drobney) was present when the site was first acquired by the refuge. Glacial till was evident where topsoil was lacking, and the plant community was depauperate. Over the years, the site was overseeded with a few conservative native plant species not originally present and Viola pedatifida was planted as plugs (K. Viste-Sparkman, U.S. Fish and Wildlife Service, unpub. data, 2022). The site’s originally depauperate state may have never supported the diversity of AMF we detected at other sites, and surrounding agriculture may have reduced opportunities for local dispersal.
Conclusion
Results of this study have provided insight into variation in AMF communities in reconstructed versus remnant prairies in Minnesota and Iowa. Consistently across this geographic range, remnant- and reconstruction-based AMF communities remained largely distinct from each other. A recent metaanalysis (Neuenkamp and others, 2019) revealed that inoculation with AMF promoted native and specialist plant species in restorations, which thereby became more similar to reference sites; the effect was especially pronounced in the most disturbed sites. In addition, Koziol and others (2021), in a restoration experiment that spanned grasslands in three States, detected greater resistance to exotic invasion in sites where AMF-inoculated late-successional nurse plants had been introduced. High-quality pollinator habitat requires the establishment of species-rich plant communities, which include late-successional and hard to establish forbs and resistance to exotic infestation. We have determined that even more than 10 years after planting, prairie reconstructions have AMF communities distinct from AMF communities in remnants. These reconstructions have been invaded by exotic cool-season grasses, while losing the native cool-season grasses that might compete with them (Larson and others, 2017). Careful application of AMF inocula may not only reduce the differences in AMF communities between reconstructions and remnants but also may improve multifunctionality and sustainability of increasingly rare prairies and the fauna they support. The differences we have detected indicate that further research to discern the effects of AMF derived from reconstructions versus remnants on plant performance is warranted.
References Cited
Asmelash, F., Bekele, T., and Birhane, E., 2016, The potential role of arbuscular mycorrhizal fungi in the restoration of degraded lands: Frontiers in Microbiology, v. 7, 15 p. [Also available at https://doi.org/10.3389/fmicb.2016.01095.]
Bauer, J.T., Kleczewski, N.M., Bever, J.D., Clay, K., and Reynolds, H.L., 2012, Nitrogen-fixing bacteria, arbuscular mycorrhizal fungi, and the productivity and structure of prairie grassland communities: Oecologia, v. 170, no. 4, p. 1089–1098. [Also available at https://doi.org/10.1007/s00442-012-2363-3.]
Bengtsson-Palme, J., Ryberg, M., Hartmann, M., Branco, S., Wang, Z., Godhe, A., De Wit, P., Sánchez-García, M., Ebersberger, I., de Sousa, F., Amend, A.S., Jumpponen, A., Unterseher, M., Kristiansson, E., Abarenkov, K., Bertrand, Y.J.K., Sanli, K., Eriksson, K.M., Vik, U., Veldre, V., and Nilsson, R.H., 2013, Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data: Methods in Ecology and Evolution, v. 4, no. 10, p. 914–919. [Also available at https://doi.org/10.1111/2041-210X.12073.]
Bever, J.D., 2002, Host-specificity of AM fungal population growth rates can generate feedback on plant growth: Plant and Soil, v. 244, no. 1/2, p. 281–290. [Also available at https://doi.org/10.1023/A:1020221609080.]
Bittebiere, A.K., Vandenkoornhuyse, P., Maluenda, E., Gareil, A., Dheilly, A., Coudouel, S., Bahin, M., and Mony, C., 2020, Past spatial structure of plant communities determines arbuscular mycorrhizal fungal community assembly: Journal of Ecology, v. 108, no. 2, p. 546–560. [Also available at https://doi.org/10.1111/1365-2745.13279.]
Börstler, B., Renker, C., Kahmen, A., and Buscot, F., 2006, Species composition of arbuscular mycorrhizal fungi in two mountain meadows with differing management types and levels of plant biodiversity: Biology and Fertility of Soils, v. 42, no. 4, p. 286–298. [Also available at https://doi.org/10.1007/s00374-005-0026-9.]
Callahan, B.J., 2022, DADA2 ITS pipeline workflow (ver. 1.8): GitHub, Inc., software release, accessed May 9, 2022, at https://benjjneb.github.io/dada2/ITS_workflow.html.
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., and Holmes, S.P., 2016, DADA2—High-resolution sample inference from Illumina amplicon data: Nature Methods, v. 13, no. 7, p. 581–583. [Also available at https://doi.org/10.1038/nmeth.3869.]
Cheeke, T.E., Schneider, M., Saify, A., Brauner, M., and Bunn, R., 2022, Role of soil biota in grassland restorations in high nutrient soils: Restoration Ecology, v. 30, no. 4, art. e13549, 14 p., accessed May 4, 2022, at https://doi.org/10.1111/rec.13549.
Davison, J., de Leόn, D.G., Zobel, M., Moora, M., Bueno, C.G., Barcelό, M., Gerz, M., Leόn, D., Meng, Y.M., Pillar, V.D., Sepp, S.K., Soudzilovaskaia, N.A., Tedersoo, L., Vaessen, S., Vahter, T., Winck, B., and Öpik, M., 2020, Plant functional groups associate with distinct arbuscular mycorrhizal fungal communities: New Phytologist, v. 226, no. 4, p. 1117–1128. [Also available at https://doi.org/10.1111/nph.16423.]
Davison, J., Öpik, M., Daniell, T.J., Moora, M., and Zobel, M., 2011, Arbuscular mycorrhizal fungal communities in plant roots are not random assemblages: FEMS Microbiology Ecology, v. 78, no. 1, p. 103–115. [Also available at https://doi.org/10.1111/j.1574-6941.2011.01103.x.]
Drobney, P., Larson, D.L., Larson, J.L., and Viste-Sparkman, K., 2020, Toward improving pollinator habitat—Reconstructing prairies with high forb diversity: Natural Areas Journal, v. 40, no. 3, p. 252–261. [Also available at https://doi.org/10.3375/043.040.0322.]
Gerla, P.J., Cornett, M.W., Ekstein, J.D., and Ahlering, M.A., 2012, Talking big—Lessons learned from a 9000 hectare restoration in the northern tallgrass prairie: Sustainability (Basel), v. 4, no. 11, p. 3066–3087. [Also available at https://doi.org/10.3390/su4113066.]
Gweon, H.S., Oliver, A., Taylor, J., Booth, T., Gibbs, M., Read, D.S., Griffiths, R.I., and Schonrogge, K., 2015, PIPITS—An automated pipeline for analyses of fungal internal transcribed spacer sequences from the Illumina sequencing platform: Methods in Ecology and Evolution, v. 6, no. 8, p. 973–980. [Also available at https://doi.org/10.1111/2041-210X.12399.]
Hausmann, N.T., and Hawkes, C.V., 2010, Order of plant host establishment alters the composition of arbuscular mycorrhizal communities: Ecology, v. 91, no. 8, p. 2333–2343. [Also available at https://doi.org/10.1890/09-0924.1.]
Hetrick, B.A.D., Wilson, G.T., Kitt, D.G., and Schwab, A.P., 1988, Effects of soil microorganisms on mycorrhizal contribution to growth of big bluestem grass in non-sterile soil: Soil Biology and Biochemistry, v. 20, no. 4, p. 501–507. [Also available at https://doi.org/10.1016/0038-0717(88)90065-X.]
Hetrick, B.A.D., Wilson, G.W.T., and Todd, T.C., 1990, Differential responses of C3 and C4 grasses to mycorrhizal symbiosis, phosphorus fertilization, and soil microorganisms: Canadian Journal of Botany, v. 68, no. 3, p. 461–467. [Also available at https://doi.org/10.1139/b90-061.]
Jack, C.N., Petipas, R.H., Cheeke, T.E., Rowland, J.L., and Friesen, M.L., 2021, Microbial inoculants—Silver bullet or microbial Jurassic Park?: Trends in Microbiology, v. 29, no. 4, p. 299–308. [Also available at https://doi.org/10.1016/j.tim.2020.11.006.]
Johnson, N.C., Wilson, G.W.T., Bowker, M.A., Wilson, J.A., and Miller, R.M., 2010, Resource limitation is a driver of local adaptation in mycorrhizal symbioses: Proceedings of the National Academy of Sciences of the United States of America, v. 107, no. 5, p. 2093–2098. [Also available at https://doi.org/10.1073/pnas.0906710107.]
Kõljalg, U., Nilsson, R.H., Abarenkov, K., Tedersoo, L., Taylor, A.F., Bahram, M., Bates, S.T., Bruns, T.D., Bengtsson-Palme, J., Callaghan, T.M., Douglas, B., Drenkhan, T., Eberhardt, U., Dueñas, M., Grebenc, T., Griffith, G.W., Hartmann, M., Kirk, P.M., Kohout, P., Larsson, E., Lindahl, B.D., Lücking, R., Martín, M.P., Matheny, P.B., Nguyen, N.H., Niskanen, T., Oja, J., Peay, K.G., Peintner, U., Peterson, M., Põldmaa, K., Saag, L., Saar, I., Schüßler, A., Scott, J.A., Senés, C., Smith, M.E., Suija, A., Taylor, D.L., Telleria, M.T., Weiss, M., and Larsson, K.H., 2013, Towards a unified paradigm for sequence-based identification of fungi: Molecular Ecology, v. 22, no. 21, p. 5271–5277. [Also available at https://doi.org/10.1111/mec.12481.]
Koziol, L., Bauer, J.T., Duell, E.B., Hickman, K., House, G.L., Schultz, P.A., Tipton, A.G., Wilson, G.W.T., and Bever, J.D., 2021, Manipulating plant microbiomes in the field—Native mycorrhizae advance plant succession and improve native plant restoration: Journal of Applied Ecology, v. 00, 10 p., accessed May 4, 2022, at https://doi.org/10.1111/1365-2664.14036.
Koziol, L., and Bever, J.D., 2016, AMF, phylogeny, and succession—Specificity of response to mycorrhizal fungi increases for late-successional plants: Ecosphere, v. 7, no. 11, art. e01555, 11 p., accessed May 4, 2022, at https://doi.org/10.1002/ecs2.1555.
Koziol, L., and Bever, J.D., 2017, The missing link in grassland restoration—Arbuscular mycorrhizal fungi inoculation increases plant diversity and accelerates succession: Journal of Applied Ecology, v. 54, no. 5, p. 1301–1309. [Also available at https://doi.org/10.1111/1365-2664.12843.]
Lahti, L., Shetty, S., Turaga, N., Gilmore, R., Leung, E., Salojärvi, J., Blake, T., Obenchain, V., Pagès, H., and Salosensaari, A., 2017, Tools for microbiome analysis in R, version 1.8.0: GitHub, Inc., software release, accessed May 9, 2022, at https://microbiome.github.io/tutorials/.
Larson, D.L., Ahlering, M.A., Drobney, P., Esser, R., Larson, J.L., and Viste-Sparkman, K., 2018, Developing a framework for evaluating tallgrass prairie reconstruction methods and management: Ecological Restoration, v. 36, no. 1, p. 6–18. [Also available at https://doi.org/10.3368/er.36.1.6.]
Larson, D.L., Bright, J.B., Drobney, P., Larson, J.L., Palaia, N., Rabie, P.A., Vacek, S., and Wells, D., 2011, Effects of planting method and seed mix richness on the early stages of tallgrass prairie restoration: Biological Conservation, v. 144, no. 12, p. 3127–3139. [Also available at https://doi.org/10.1016/j.biocon.2011.10.018.]
Larson, D.L., Bright, J.B., Drobney, P., Larson, J.L., and Vacek, S., 2017, Persistence of native and exotic plants 10 years after prairie reconstruction: Restoration Ecology, v. 25, no. 6, p. 953–961. [Also available at https://doi.org/10.1111/rec.12521.]
Larson, D.L., Hernández, D.L., Larson, J.L., Leone, J.B., and Pennarola, N., 2020, Management of remnant tallgrass prairie by grazing or fire in western Minnesota, 2016–2017: U.S. Geological Survey data release, accessed May 4, 2022, at https://doi.org/10.5066/P9N8X0ZY.
Larson, J.L., Aldrich-Wolfe, L., Vink, S., Huerd, S.C., Vacek, S., Drobney, P., Jordan, N.R., and Larson, D.L., 2021, Arbuscular mycorrhizal fungi in remnant and reconstructed prairies in Minnesota and Iowa, 2019 (ver. 2.0, April 2022): U.S. Geological Survey data release, accessed May 4, 2022, at https://doi.org/10.5066/P95R5UNN.
Maltz, M.R., and Treseder, K.K., 2015, Sources of inocula influence mycorrhizal colonization of plants in restoration projects—A meta-analysis: Restoration Ecology, v. 23, no. 5, p. 625–634. [Also available at https://doi.org/10.1111/rec.12231.]
Martignoni, M.M., Garnier, J., Zhang, X., Rosa, D., Kokkoris, V., Tyson, R.C., and Hart, M.M., 2021, Co-inoculation with arbuscular mycorrhizal fungi differing in carbon sink strength induces a synergistic effect in plant growth: Journal of Theoretical Biology, v. 531, art. 110859, 9 p. [Also available at https://doi.org/10.1016/j.jtbi.2021.110859.]
Martin, M., 2011, Cutadapt removes adapter sequences from high-throughput sequencing reads: EMBnet.Journal, v. 17, no. 1, p. 10–12. [Also available at https://doi.org/10.14806/ej.17.1.200.]
McLauchlan, K., 2006, The nature and longevity of agricultural impacts on soil carbon and nutrients—A review: Ecosystems (New York), v. 9, no. 8, p. 1364–1382. [Also available at https://doi.org/10.1007/s10021-005-0135-1.]
McMurdie, P.J., and Holmes, S., 2013, phyloseq—An R package for reproducible interactive analysis and graphics of microbiome census data: PLoS One, v. 8, no. 4, art. e61217, 11 p., accessed May 4, 2022, at https://doi.org/10.1371/journal.pone.0061217.
Middleton, E.L., and Bever, J.D., 2012, Inoculation with a native soil community advances succession in a grassland restoration: Restoration Ecology, v. 20, no. 2, p. 218–226. [Also available at https://doi.org/10.1111/j.1526-100X.2010.00752.x.]
Minnesota Biological Survey, 2014, Upland Prairie System—Condition ranking guidelines: St. Paul, Minn., Minnesota Department of Natural Resources, accessed May 4, 2022, at https://files.dnr.state.mn.us/eco/mcbs/upland_prairie_system_ranking_guidelines.pdf.
Neuenkamp, L., Prober, S.M., Price, J.N., Zobel, M., and Standish, R.J., 2019, Benefits of mycorrhizal inoculation to ecological restoration depend on plant functional type, restoration context and time: Fungal Ecology, v. 40, p. 140–149. [Also available at https://doi.org/10.1016/j.funeco.2018.05.004.]
Nicolson, S.W., and Wright, G.A., 2017, Plant-pollinator interactions and threats to pollination—Perspectives from the flower to the landscape: Functional Ecology, v. 31, no. 1, p. 22–25. [Also available at https://doi.org/10.1111/1365-2435.12810.]
Oehl, F., Sieverding, E., Ineichen, K., Mader, P., Boller, T., and Wiemken, A., 2003, Impact of land use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosystems of Central Europe: Applied and Environmental Microbiology, v. 69, no. 5, p. 2816–2824. [Also available at https://doi.org/10.1128/AEM.69.5.2816-2824.2003.]
Oksanen, J., Gauilaume Blanchet, F., Friendly, M., Kindt, R., Legendre, P., McGlinn, D.J., Minchin, P.R., O’Hara, R.B., Simpson, G.L., Solymos, M., Stevens, H., Szӧcs, E., and Wagner, H., 2020, vegan—Community Ecology Package (version 2.5–7): R Foundation for Statistical Computing software release, accessed May 4, 2022, at https://cran.r-project.org/.
Pagès, H., Aboyoun, P., Gentleman, R., and DebRoy, S., 2019, Biostrings—Efficient manipulation of biological strings (version 2.54.0): R Foundation for Statistical Computing software release, accessed May 4, 2022, at https://bioconductor.org/packages/release/bioc/html/Biostrings.html.
Paz, C., Öpik, M., Bulascoschi, L., Bueno, C.G., and Galetti, M., 2021, Dispersal of arbuscular mycorrhizal fungi—Evidence and insights for ecological studies: Microbial Ecology, v. 81, no. 2, p. 283–292. [Also available at https://doi.org/10.1007/s00248-020-01582-x.]
Perkins, L.B., Ahlering, M., and Larson, D.L., 2019, Looking to the future—Key points for sustainable management of northern Great Plains grasslands: Restoration Ecology, v. 27, no. 6, p. 1212–1219. [Also available at https://doi.org/10.1111/rec.13050.]
R Core Team, 2020, R—A language and environment for statistical computing: Vienna, Austria, R Foundation for Statistical Computing, version 3.6.3 (Holding the Windsock), accessed March 10, 2020, at https://www.r-project.org/.
Revillini, D., Gehring, C.A., and Johnson, N.C., 2016, The role of locally adapted mycorrhizas and rhizobacteria in plant–soil feedback systems: Functional Ecology, v. 30, no. 7, p. 1086–1098. [Also available at https://doi.org/10.1111/1365-2435.12668.]
Sanders, I.R., and Rodriguez, A., 2016, Aligning molecular studies of mycorrhizal fungal diversity with ecologically important levels of diversity in ecosystems: The ISME Journal, v. 10, no. 12, p. 2780–2786. [Also available at https://doi.org/10.1038/ismej.2016.73.]
Schnoor, T.K., Lekberg, Y., Rosendahl, S., and Olsson, P.A., 2011, Mechanical soil disturbance as a determinant of arbuscular mycorrhizal fungal communities in semi-natural grassland: Mycorrhiza, v. 21, no. 3, p. 211–220. [Also available at https://doi.org/10.1007/s00572-010-0325-3.]
Schulte, L.A., Niemi, J., Helmers, M.J., Liebman, M., Arbuckle, J.G., James, D.E., Kolka, R.K., O’Neal, M.E., Tomer, M.D., Tyndall, J.C., Asbjornsen, H., Drobney, P., Neal, J., Van Ryswyk, G., and Witte, C., 2017, Prairie strips improve biodiversity and the delivery of multiple ecosystem services from corn-soybean croplands: Proceedings of the National Academy of Sciences of the United States of America, v. 114, no. 42, p. 11247–11252. [Also available at https://doi.org/10.1073/pnas.1620229114.]
Suding, K.N., Gross, K.L., and Houseman, G.R., 2004, Alternative states and positive feedbacks in restoration ecology: Trends in Ecology & Evolution, v. 19, no. 1, p. 46–53. [Also available at https://doi.org/10.1016/j.tree.2003.10.005.]
Tipton, A.G., Middleton, E.L., Spollen, W.G., and Galen, C., 2019, Anthropogenic and soil environmental drivers of arbuscular mycorrhizal community composition differ between grassland ecosystems: Botany, v. 97, no. 1, p. 85–99. [Also available at https://doi.org/10.1139/cjb-2018-0072.]
Werner, G.D.A., and Kiers, E.T., 2015, Partner selection in the mycorrhizal mutualism: New Phytologist, v. 205, no. 4, p. 1437–1442. [Also available at https://doi.org/10.1111/nph.13113.]
Wolfe, B.E., Husband, B.C., and Klironomos, J.N., 2005, Effects of a belowground mutualism on an aboveground mutualism: Ecology Letters, v. 8, no. 2, p. 218–223. [Also available at https://doi.org/10.1111/j.1461-0248.2004.00716.x.]
Appendix 1. Details of Sampled Sites
Table 1.1.
Sampling sites and plant species used in the study, how the species abundance changed between 2007 and 2015 in reconstructions monitored, and the coefficient of conservatism of each species. The number of samples of each plant species collected at each study site is listed.[CofC, coefficient of conservatism; WPA, Waterfowl Production Area; dec, decreased; --, no data or not applicable; nc, no change; fe, failed to establish; inc, increased]
Table 1.2.
Additional sampling sites and plant species used in the study, how the species abundance changed between 2007 and 2015 in reconstructions monitored, and the coefficient of conservatism of each species. The number of samples of each plant species collected at each study site is listed.[CofC, coefficient of conservatism; WPA, Waterfowl Production Area; SNA, Scientific and Natural Area; WMA, Wildlife Management Area; dec, decreased; --, no data or not applicable; nc, no change; fe, failed to establish; inc, increased]
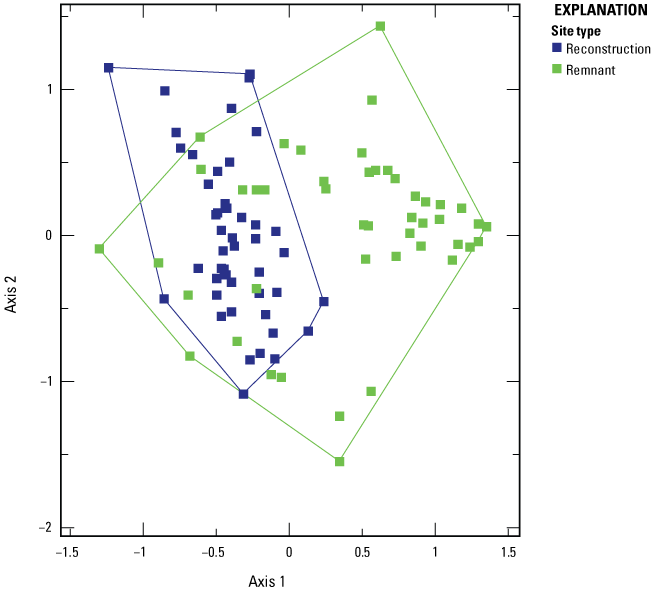
Arbuscular mycorrhizal fungi (AMF) communities in roots of plant species at 14 sites, 6 of which were remnants and 8 of which were reconstructions. Each point represents the AMF community of a single plant species at a single site. Only plant species detected at reconstructions and remnants were used in the ordination. Note that nonmetric multidimensional scaling axes are arbitrary, so this ordination is about 180 degrees from the ordination in figure 2 but shows the same pattern of separation between AMF communities in remnant and reconstructed prairies.
Conversion Factors
For more information about this publication, contact:
Director, USGS Northern Prairie Wildlife Research Center
8711 37th Street Southeast
Jamestown, ND 58401
701–253–5500
For additional information, visit: https://www.usgs.gov/centers/npwrc
Publishing support provided by the
Indianapolis, Reston, and Rolla Publishing Service Centers
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
Vink, S.N., Aldrich-Wolfe, L., Huerd, S.C., Larson, J.L., Vacek, S.C., Drobney, P.M., Barnes, M., Viste-Sparkman, K., Jordan, N.R., and Larson, D.L., 2022, Belowground mutualisms to support prairie reconstruction—Improving prairie habitat using mycorrhizal inoculum: U.S. Geological Survey Open-File Report 2022–1055, 18 p., https://doi.org/10.3133/ofr20221055.
ISSN: 2331-1258 (online)
Study Area
| Publication type | Report |
|---|---|
| Publication Subtype | USGS Numbered Series |
| Title | Belowground mutualisms to support prairie reconstruction—Improving prairie habitat using mycorrhizal inoculum |
| Series title | Open-File Report |
| Series number | 2022-1055 |
| DOI | 10.3133/ofr20221055 |
| Publication Date | June 09, 2022 |
| Year Published | 2022 |
| Language | English |
| Publisher | U.S. Geological Survey |
| Publisher location | Reston, VA |
| Contributing office(s) | Northern Prairie Wildlife Research Center |
| Description | Report: vi, 18 p.; 2 Data Releases |
| Country | United States |
| State | Iowa, Minnesota |
| Online Only (Y/N) | Y |