

Open Water Control of Invasive Mussels Using Benthic Mats—Part 1, Short-Term Infusion of Carbon Dioxide Under a Mat
Links
- Document: Report (2.33 MB pdf) , HTML , XML
- Data Release: USGS data release - Evaluation of benthic barrier layers/tarps for open water control of invasive mussels in 2024 in Loon Lake, Benzie Co., Michigan, USA
- Software Release: USGS software release - Analysis of open water control of invasive mussels using benthic mats. Part 1—Short-term infusion of carbon dioxide under a mat
- Download citation as: RIS | Dublin Core
Acknowledgments
The project was funded by the U.S. Environmental Protection Agency Great Lakes Restoration Initiative and U.S. Geological Survey Biological Threats and Invasive Species Research Program. Our deepest thanks to National Park Service staff at Sleeping Bear Dunes National Lakeshore for their support. We also thank Allie Holdhausen, National Park Service, Mississippi National Recreation Area, and the outstanding dive teams from the U.S. Geological Survey Great Lakes Science Center for providing dive services for the project.
Abstract
This study compared the efficacy of a benthic mat alone with carbon dioxide infusion under a mat for killing Dreissena polymorpha (Pallas, 1771) (zebra mussel). Three sites were selected in Loon Lake, Sleeping Bear Dunes National Lakeshore, Benzie County, Michigan, for replication of reference, benthic mat, and carbon dioxide mat treatments. Within a site, three 4-meter (m) x 4-m plots were delineated for each treatment and a reference. Pretreatment samples were collected to estimate zebra mussel density and macroinvertebrate community composition in reference plots. Zebra mussels (about 360) from outside of the treatment plots were caged and placed in the plots before treatment. Benthic mats (4.25 m x 4.25 m; polyethylene with a vinyl coating) were anchored on the lake bottom with sandbags and weights. Carbon dioxide was infused under a mat of the same material to a maximum of 200 milligrams per liter (mg/L; pH=6.13) every 2–4 hours, for about 12 hours. Benthic and carbon dioxide mats were deployed for 5 days. One day after mat removal, we assessed mortality of resident and sentinel caged zebra mussels and macroinvertebrate community abundance and diversity in each plot. Average pH (as a proxy for carbon dioxide) under the carbon dioxide mats was between 6.38 and 6.80, equivalent to 170.5 and 103.0 mg/L carbon dioxide, respectively. In the posttreatment survey, few zebra mussels were observed in the benthic mat and carbon dioxide treatment plots compared to the reference plots; survival was lowest in the carbon dioxide plots. Mortality of sentinel caged mussels was greater than 80 percent in carbon dioxide treatments compared to mean mortalities of 20.6 percent and 12.7 percent in the benthic mat and reference plots, respectively. Macroinvertebrate community total abundance was lower in both mat treatments compared to reference plots, but diversity was comparable among all treatments. Our study demonstrated that carbon dioxide treatment near 200 mg/L could produce greater than 80-percent mortality of zebra mussels within 5 days. Refinement of the carbon dioxide mat and delivery system could increase spatial coverage of the treatment and broaden its use to other habitats.
Introduction
Dreissena polymorpha (Pallas, 1771) (zebra mussel) and D. rostiformis bugensis (Andrusov, 1897) (quagga mussel) are nonindigenous invasive species introduced to North America in the 1980s, presumably by transatlantic ships arriving at the Laurentian Great Lakes (Hebert and others, 1989; Mills and others, 1996). Since their arrival, dreissenid mussels have caused substantial ecological damage across North America. As prolific filter feeders, these mussels remove suspended particles and phytoplankton, resulting in decreased food availability for native planktivores and increased macrophyte abundance from increased light transmittance (MacIsaac, 1996). Dreissenids can cause a shift in nutrients from the pelagic to the benthic zone (Higgins and Vander Zanden, 2010), changing trophic structure and adversely affecting fish populations (McEachran and others, 2019; Hansen and others, 2020; Zorn and Kramer, 2022; Cunningham and Dunlop, 2023). Zebra mussels have exacerbated the decline of native freshwater mussels (Order Unionida) by competing for food and habitat and biofouling native mussel shells (Haag and others, 1993; Dzierżyńska‐Białończyk and others, 2018; Beason and Schwalb, 2022). Options for controlling invasive mussels in open water are limited. Physical control tools have been used as an alternative to chemical methods that may persist after application. For example, benthic mats were used in Lake Tahoe (not shown) (Wittmann and others, 2012); Lake Waco (not shown) (Conry and others, 2024); and Good Harbor Reef, Lake Michigan (not shown) (Kunze, 2023), to kill molluscs in selected, localized areas. Mats must remain in place for weeks to months to create anoxic conditions that kill dreissenids. These lengthy treatment times thereby restrict benthic mat use to a small number of placements each year in temperate areas where seasonal temperatures and winter freezing limit deployment to warmer months. Additionally, the anoxic conditions under the mat can cause substantial nontarget mortality.
The goal of this study was to determine if we could reduce open water benthic mat treatment times and potentially reduce adverse effects to nontarget benthic invertebrates by introducing carbon dioxide as a “selective toxin” under the mats. Benthic mats have been used to contain molluscicides to help maintain an effective concentration in the benthic habitat. For example, the biopesticide Zequanox was applied under a benthic mat at Good Harbor Reef, Leelanau County, Michigan, in a dreissenid mussel control demonstration (LimnoTech, 2020). In this study, we evaluated an alternative to Zequanox that could be more cost effective. Carbon Dioxide–Carp is registered as a pesticide for use in open water to control aquatic invasive species (U.S. Environmental Protection Agency, 2019). In laboratory studies, carbon dioxide has demonstrated selective toxicity for zebra mussels compared to unionid mussels. For example, at 20 degrees Celsius (°C), the 96-hour lethal concentration to 10 percent of animals for Lampsilis cardium (plain pocketbook) was 173 standard atmosphere (atm) partial pressure of carbon dioxide (PCO2; 95-percent confidence interval [CI] of 147–198 atm; about [~] 260 milligrams per liter [mg/L] carbon dioxide) compared to a lethal concentration to 99 percent of zebra mussels of 118 atm PCO2 (CI 109–127 atm; ~170 mg/L carbon dioxide). The lethal time to 99-percent mortality of zebra mussels in 110–120 atm PCO2 ranged from 100 hours at 20 °C to 300 hours at 5 °C. Mean survival of juvenile L. siliquoidea (fatmucket) exceeded 85 percent in carbon dioxide treatments that were lethal to 99 percent of zebra mussels (Waller and others, 2020). Exposure to 100 mg/L carbon dioxide for 10 weeks did not cause juvenile mussel mortality but did reduce shell growth and tissue condition; the same treatment also reduced overall benthic invertebrate biomass, but chironomid and oligochaete abundance increased (Waller and others, 2021). Carbon dioxide infusion under a mat could produce mortality faster than a benthic mat alone, allowing multiple deployments of the carbon dioxide mat over the same time needed for a single benthic mat deployment.
In this study, we tested a novel system for delivery of carbon dioxide water under a benthic mat as a tool to kill zebra mussels. We compared the response of zebra mussels and benthic macroinvertebrates among reference, benthic mat, and carbon dioxide mat treatments.
Purpose and Scope
The purpose of the report is to describe the evaluation of carbon dioxide infusion under a benthic mat for invasive mussel control in open water. Data were collected and analyzed by the U.S. Geological Survey from before and after replicated benthic mat treatments with and without carbon dioxide infusion. Experimental treatments were done at an inland lake in Michigan in 2024. Specifically, the report includes results of water quality conditions before, during, and after treatments; density, size (shell length), and percentage of live resident zebra mussels on colonized substrates in study plots; size and mortality of caged sentinel zebra mussels after treatment; and macroinvertebrate community diversity and abundance before (in reference plots) and after treatment (reference and treatment plots). Data collected from this study are available in the data release (Meulemans and others, 2026). Code that supports this study is available in a software release (Waller and others, 2026).
Methods
This section of the report describes the study location and treatment plots used in the evaluation of benthic mats with and without carbon dioxide infusion. Methods are described for delineation of reference, benthic mat, and carbon dioxide mat treatment plots; design and deployment of the carbon dioxide infusion system; and development of standard curve to estimate carbon dioxide concentration from pH. We also describe methods to assess posttreatment mortality of resident and sentinel zebra mussels, the macroinvertebrate community, and water quality.
Study Location and Treatment Plots
The study was completed at Sleeping Bear Dunes National Lakeshore in Loon Lake (44° 42′ 30.24″ N., 86° 07′ 48″ W.), Benzie County, Mich. (fig. 1). Loon Lake is 38.4 hectares in surface area with a maximum depth of 20.1 meters (m) and mean depth of 9.0 m (Stockwell and Gannon, 1975). The Platte River enters Loon Lake on the eastern shoreline and outlets at the northernmost extension of the lake (Stockwell and Gannon, 1975). The lake has a well-established population of zebra mussels and few extant native mussels (Nichols and others, 2003; D. Woolnough, Central Michigan University, unpub. data, March 2024).
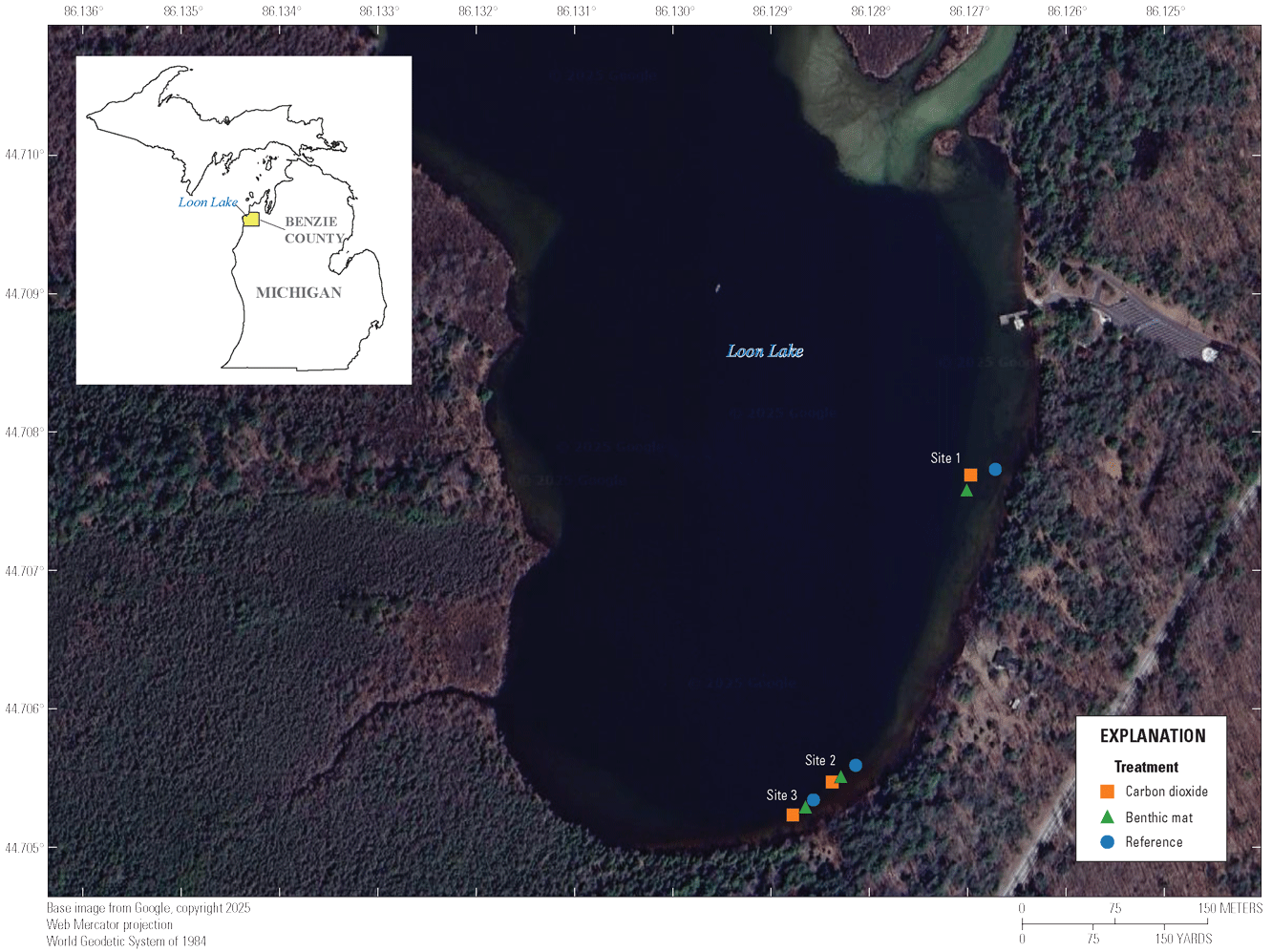
Map showing Loon Lake, Benzie County, Michigan, and the locations of three study sites. At each site, three 16-square-meter plots were designated as reference, benthic mat, and carbon dioxide mat plots.
In June 2024, divers completed reconnaissance surveys throughout Loon Lake to assess zebra mussel density and to locate suitable study sites. The substrate in the littoral zone is sand and few cobbles and an overlay of marl. The lakebed drops rapidly to greater than (>) 6 m in depth within ~30 m from the shore. The substrate at depths >3 m is primarily silt and fine organic material. Most of the available substrate for zebra mussel colonization is woody debris. Divers reported that zebra mussel populations were sparsely distributed in clumps in association with the available hard substrate. We avoided the northern half of the lake between the inflow and outflow of the Platte River because it is heavily used by recreational boaters. Sites were selected south of the Platte River inflow along the eastern shoreline, between ~1 and 3 m in depth, and containing some mussel-colonized substrate. Three sites were selected (sample size = 3 benthic mat, 3 carbon dioxide mat, 3 reference) for replication of reference, benthic mat, and carbon dioxide mat treatments (fig. 1). Our experimental unit was the site. Within a site, three 4-m x 4-m plots were delineated for each treatment and a reference. Plots within each site were separated by ~8–16 m; water depth in the plots ranged from 0.5 to 2.0 m. Small floats marked the corners of each plot, and the midpoint of each carbon dioxide plot was recorded from a portable Global Positioning System (table 1).
Test Treatment Materials
The material used for the benthic and carbon dioxide infusion mats was woven polyethylene with a vinyl coating that was 14 mil thick, acid resistant, and nonreactive. Each mat was 4.25 m x 4.25 m, which included a 0.25-m apron for placement of anchoring weights. The benthic mat was rolled onto a 6-m (length) x 15-centimeter (cm; diameter) polyvinyl chloride (PVC) pipe for deployment. The mat was carried from shore and unrolled over the plot by divers. The perimeter of the mat was anchored by sandbags (~10 bags per mat) and cinder blocks to prevent influx of freshwater under the mat.
The carbon dioxide infusion mat was supported by a fiberglass and PVC frame (fig. 2) consisting of a center hub and four arms extending to each corner of the mat. The top of the hub had a connecting port for inflowing carbon dioxide-infused water, and the lower part of the hub had a base (strainer grate, 0.5-m height x 15-cm diameter) to hold the center of the mat above the substrate. Four arms (3 m in length) made of fiberglass rods surrounded by 19-millimeter diameter PVC pipe extended from the hub to each corner of the mat, creating a frame for attachment of the mat. The arms were connected to the hub by 180-degree hinges that allowed unfolding the mat for deployment and collapsing the arms for retrieval. Vinyl loops (~5 cm x 15 cm) were glued to the top of the mat to secure the arms of the frame. When fully deployed, the mat covered 16 square meters (m2), not including the apron (0.25-m width) around the perimeter, which was used for placement of sandbags to anchor and seal the mat edge. Additionally, a chain (15.8 mm width) was placed on the apron around the perimeter of the mat to reduce loss of carbon dioxide from the mat.
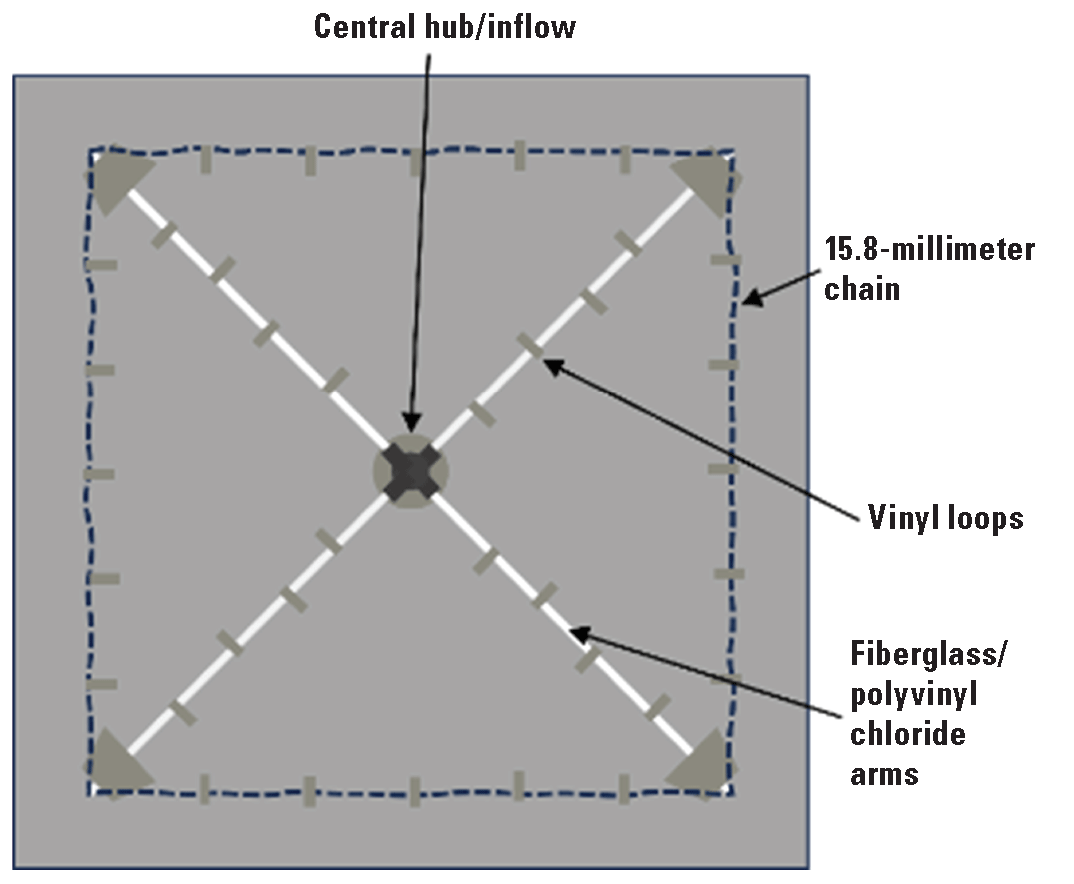
Diagram showing the carbon dioxide frame and mat design, as viewed from the top. The mat was 4.25 meters x 4.25 meters. The frame consisted of four arms connected to a central hub. The center of the hub had a connection for delivery of carbon dioxide-infused water and a 0.5-meter base to elevate the center of the mat.
Carbon dioxide was applied as Carbon Dioxide–Carp (U.S. Environmental Protection Agency registration no. 6704–95) under the regulatory authority of Michigan Department of Environmental Quality Aquatic Nuisance Control (Certificate of Coverage no. MIG032005) and U.S. Environmental Protection Agency Experimental Use Permit exemption for application outside the scope of the current label. Hereafter, we refer to Carbon Dioxide–Carp as “carbon dioxide.” The carbon dioxide infusion and delivery system consisted of two diaphragm pumps (Seaflo model SVDP1, 20-liter per minute flow rate) to deliver lake water, a 30-pound cylinder of carbon dioxide (food-grade compressed carbon dioxide gas), gas regulator and flow meters, two downward-flow gas infusion chambers, and one outflow hose. The infusion and delivery system was mounted to brackets on a 5.5-m (length) aluminum boat (fig. 3), and the proximate end of the delivery hose was secured to either a navigation buoy or a cylinder block adjacent to each carbon dioxide plot. Water was pumped from the lake into the infusion chambers and a regulator and flow meter were used to adjust delivery rate of carbon dioxide to a large air stone within the infusion chambers. The outflowing water from the two chambers was connected to a single hose for delivery of carbon dioxide infused water to the center of the mat.

Photograph showing carbon dioxide delivery system.
Pretreatment Assessment of Zebra Mussel Density and Macroinvertebrate Communities
The purpose of the pretreatment survey was to estimate the percentage of live and freshly dead mussels in the plots before treatment; however, the reconnaissance survey of the study sites determined that hard substrate was scarce in the plots and zebra mussel density was highly clustered on woody debris. Therefore, we limited the pretreatment survey to the reference plots and completed biased quadrat surveys by actively searching for colonized substrate (such as woody debris, rocks, and plastic debris). Divers placed a 1-m x 1-m quadrat in areas with suitable substrate, collected attached zebra mussels and (or) the substrate from within the quadrat, placed the sample in a mesh dive bag, and returned the bag to the surface for enumeration. Four quadrats were collected from each reference plot (total area surveyed was 4 m2 per plot). Zebra mussels were assessed as live or freshly dead. Shell was not included in estimates of mussel density. Freshly dead mussels were defined as valves agape with soft tissue intact or intact valves and shiny shell nacre. Older dead shell was defined as single or intact valves with chalky nacre and darkened or eroded periostracum. After completion of pretreatment surveys, divers collected colonized substrates from the area outside of the plots and placed them into plots to augment zebra mussel density.
The macroinvertebrate community at each site was sampled by collecting triplicate petite Ponar (grab) samples (0.046-m2 sampling area) in the reference plots. We did not collect Ponar samples inside the treatment plots to minimize disturbance. Samples were washed through a 500-micron screen in the field; preserved in 70 percent ethanol; and transferred to the laboratory for sorting, identification, and enumeration. Macroinvertebrates were identified to family level, except for oligochaetes and chironomids.
Sentinel Mussels
Zebra mussels were collected from the lake in areas outside of the study sites for placement as sentinel mussels and for a standard assessment of mortality in each plot. Mussels were gently scraped with a scalpel or paint scraper from docks, rocks, and woody debris 1 to 2 days before treatment began and transferred to a cooler of lake water. Within several hours of collection, we assessed each mussel for viability by applying gentle pressure to open the valves; mussels that resisted opening were deemed suitable for testing. Mussels were indiscriminately distributed in groups of 28–32, caged within a nylon mesh bag (40-cm length) held open with a PVC ring (7.6-cm diameter x 2.5-cm height; fig. 4A), and transferred to totes filled with lake water for overnight holding. The next day, each cage was indiscriminately removed from the tote, placed onto a plastic tray (15 cm x 25 cm) along with a brick, and secured inside a second mesh bag (60-cm length). The tray prevented the mussel cages from sinking into the substrate and becoming covered with sediment, and the brick prevented the tray from overturning (fig. 4B). Within each plot, 12 cages of sentinel mussels were distributed in a similar pattern: 1 at each corner, 4 near the midpoint, and 4 halfway between the midpoint and each corner (fig. 5). A total of 360 mussels were placed in each plot.

Photographs showing (A) sentinel Dreissena polymorpha (zebra mussels) caged within a mesh bag that was held open by a rigid plastic ring and (B) zebra mussel cages placed onto a tray along with a brick. The tray and contents were placed into a second mesh bag for containment. Within each plot, 12 trays were deployed.
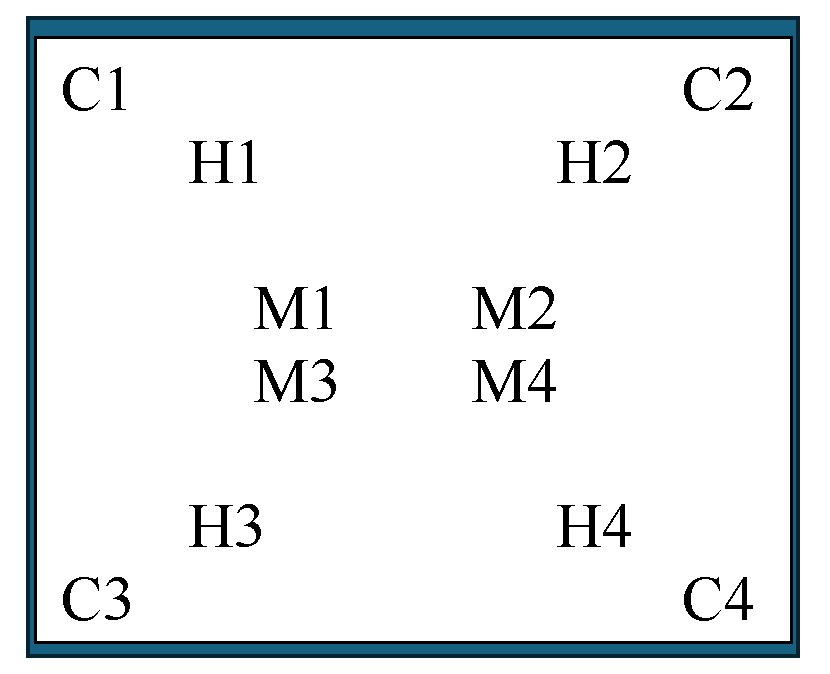
Diagram showing arrangement of sentinel Dreissena polymorpha (zebra mussel) cages within each plot. Each cage had 30 (plus or minus 2) mussels (360 mussels per plot). Four cages were placed in each area under the mat: the corners (C), midpoints (M), and halfway from each corner to each midpoint (H).
Reference Plots
Reference plots were located at a distance (8–16 m) from the carbon dioxide mat to prevent the potential effect of carbon dioxide leakage and minimize disturbance in the plot. On the day of mat deployment, the plot corners were marked by small floats, and caged sentinel zebra mussels were placed into the plot by divers, as described in the previous section. A miniDO2T (PME; Vista, California) dissolved oxygen (DO) and temperature logger was placed in the center of the plot for continuous measurement of these variables.
Benthic Mat Application
Benthic mat plots were between the carbon dioxide plot and reference plot but at a distance of 8–16 m to prevent the potential effect of carbon dioxide leakage and minimize between-plot disturbance. On the day of mat deployment, the plot corners were marked by small floats, and cages of zebra mussels were placed into the plot by divers, as described in the previous section. A miniDO2T DO and temperature logger was placed in the center of the plot for continuous measurement of these variables. The benthic mat was rolled out and over the plot by divers and anchored in place with sandbags.
Carbon Dioxide Measurement and Application
We used pH as a proxy for carbon dioxide concentration. Before treatment began, a pH-carbon dioxide concentration curve was developed with lake water. A sample of lake water was infused with carbon dioxide to produce ~50, 100, 150, 200, 250, and 300 mg/L carbon dioxide (fig. 6). The concentration of dissolved carbon dioxide at each pH level was determined by titration in triplicate with 0.363 normal (N) or 3.63 N sodium hydroxide to a pH of 8.3. Based on the curve, the target pH to produce 200 mg/L carbon dioxide was estimated at 6.13 (95-percent CI of 5.90–6.30).
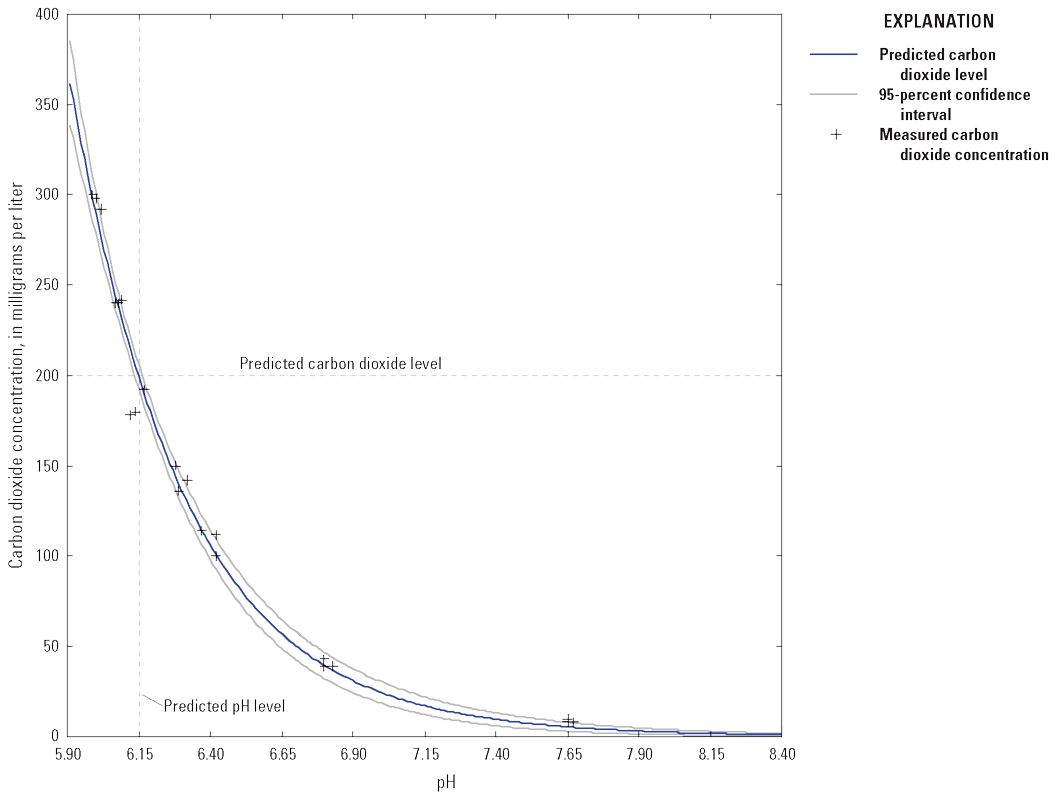
Graph showing carbon dioxide-pH curve using Loon Lake water. Carbon dioxide was bubbled into a sample of lake water, and dissolved carbon dioxide was measured by titration across a range of pH levels. The target carbon dioxide concentration of 200 milligrams per liter was estimated at pH 6.13.
On the day of carbon dioxide mat deployment, a YSI 6920 V2 sonde was placed ~1 m from the plot midpoint for real-time, continuous measurement of pH (as a surrogate for carbon dioxide concentration), temperature, and DO. The proximal end of the sonde cable was secured either to a navigation buoy or a cinder block on shore. Divers moved the carbon dioxide mat from the boat or shore and placed it over the plot. The corners were anchored with sandbags, and the perimeter was secured with additional sandbags (~10 sandbags per mat) and chain. The midpoint of the mat was elevated ~0.5 m above the substrate by the central hub to maintain adequate water volume for carbon dioxide infusion.
Carbon dioxide was intermittently infused under the mats for 120 hours (5 days) during August 20–24, 2024, in plot 1 and during August 21–25, 2024, in plots 2 and 3. The pH under the carbon dioxide mat was monitored continuously during infusion, and when the target pH (6.13 plus or minus 0.22) was reached, water injection under the mat was stopped, and the hose and sonde cable were disconnected and secured to the navigation buoy or cinder block on shore. Every 2–4 hours between dawn and dusk, we reconnected to the sonde and measured pH and DO. Additional “bump” treatments of carbon dioxide were applied for pH >6.20 to maintain the concentration near the target.
Water Quality Monitoring
Water chemistry (alkalinity, hardness) was measured on grab samples collected at two time points, during and after treatment, and two locations during the study. One sample was collected near the site 1 benthic mat plot and another between sites 2 and 3. Total hardness and alkalinity were analyzed in triplicate using the ethylenediaminetetraacetic acid titrimetric method (method 2340C; American Public Health Association, 2018) and by titrating to an endpoint of pH 4.5 (method 2320B; American Public Health Association, 2018), respectively.
Water quality (DO, pH, temperature, and specific conductance) was measured daily at each plot. We used a Van Dorn sampler to collect water ~0.3 m above the bottom. Measurements were made immediately after collection using a water-quality meter equipped with luminescent optical DO, pH, and specific conductance probes (models HQ40d, LDO 10101, PHC 70501, and CDC40101; Hach Company, Loveland, Colorado); temperature was measured with a digital thermometer (Thermapen model Mk4; ThermoWorks Company, American Fork, Utah).
Treatment Termination
At 5 days (120 hours), the sandbags were removed from the perimeter of the benthic mat, and divers floated the mat to the surface and moved it away from the plot with minimal disturbance of the substrate. In carbon dioxide treatment plots, the perimeter of the mat was lifted to allow gradual influx of freshwater and dilution of the carbon dioxide concentration. This was done to minimize the discharge of carbon dioxide to the surrounding water when the mat was removed. We monitored dissipation of the carbon dioxide by measuring pH every 5–10 minutes at the edge of the mat, middepth, and 3 m from the edge. We used the sonde pH measurement to determine when carbon dioxide under the mat was equal to ambient levels (outside of the mat). The next day, we lifted the mat to the surface and moved it away from the plot with minimal disturbance of the substrate. In all plots the miniDO2T sensor was retrieved, logging was stopped, and the data were downloaded. Caged sentinel zebra mussels were retrieved by divers and transferred to the boat for immediate assessment of mortality.
Posttreatment Sampling of Zebra Mussels and Macroinvertebrates
After removal of benthic mats and carbon dioxide-infused mats, we assessed mortality of caged zebra mussels and the percentage of live resident zebra mussels. Because of the limited density of resident zebra mussels in the plots, divers searched the entirety of each 4-m x 4-m plot for zebra mussels after mat removal. Colonized debris and hard substrate were collected and placed into a dive bag, transferred to shore, and enumerated for live and freshly dead zebra mussels. Divers retrieved and transferred caged zebra mussels to shore for assessment. Mussels were assessed as dead or alive based on the criteria that were previously described. Live and freshly dead mussels were retained for shell length measurement. We also collected macroinvertebrate samples and measured water quality in each plot following the methods described previously in “Pretreatment Assessment of Zebra Mussel Density and Macroinvertebrate Communities.”
We measured shell length of a subsample of sentinel mussels from cages (number of measurements [n] =100 mussels per site) and resident mussels from posttreatment surveys, using a digital caliper (nearest 0.1 millimeter). The number of mussels collected in surveys varied among sites, ranging from 16 (site 2) to >150 (site 1); therefore, we measured 100 mussels from site 1 and all mussels collected from sites 2 and 3.
Data Analyses
Data summary and analysis include the following: (1) water hardness and alkalinity, (2) water quality (DO, pH, temperature, and specific conductance) during and after the treatment period, (3) the density and percentage of live resident zebra mussels within the reference plot before treatment, (4) the density and percentage of live resident zebra mussels in all plots 1 day after treatment, (5) percentage of mortality of caged sentinel zebra mussels in all plots, (6) length of resident zebra mussels (collected in plot surveys) and caged sentinel zebra mussels, (7) pH and estimated carbon dioxide concentration during the exposure period, (8) macroinvertebrate diversity and abundance in reference plots before treatment, and (9) macroinvertebrate diversity and abundance in all plots 1 day after treatment. The experimental unit in all analyses was the individual plots. Significance was defined as α=0.05. We used R (ver. 4.5.1; R Core Team, 2025) for confirmatory data analysis, and key packages included vegan (ver. 2.7–1; Oksanen and others, 2025) for multivariate statistics, lme4 (ver. 1.1.37; Bates and others, 2015) and lmerTest (ver. 3.1.3; Kuznetsova and others, 2017) for mixed-effect models, and ggplot2 (ver. 3.5.2; Wickham, 2016) for plotting. Analysis of variance was used to test for treatment and time (pre- and posttreatment) effects on macroinvertebrate abundance and diversity. We used distance-based redundancy analysis with Manhattan distance to examine overall changes in macroinvertebrates by treatment and time (Legendre and Legendre, 2012). We used a generalized linear model with binomial error family and logit link function (also known as a logistic regression) to estimate the effect of treatments on zebra mussel survival for caged sentinel mussels (Bolker, 2008). A generalized linear mixed-effect model with binomial error family and logit link function was used to estimate the effect of treatments and exposure time (pre- and posttreatment) on resident zebra mussel survival collected in surveys with site treated as a random effect (Bolker, 2008). We used a linear model to estimate the association of mussel shell length versus site, group, and the interaction of site and group (Bolker 2008). We defined the association as α=0.05 for the type I error rate. The software release for data analysis is available at Waller and others (2026).
Results
Water Quality Conditions
Measurements of water quality before treatment in the study plots averaged a pH level of 8.36 (standard deviation [SD] 0.12), DO of 9.00 mg/L (SD 0.14), and conductivity of 423.3 (SD 124.5; n=3). Water chemistry from site 1 (n=6) and composite samples from sites 2 and 3 (n=6) had mean alkalinity of 142.8 mg/L as calcium carbonate (SD 1.99) and mean hardness of 143.8 mg/L as calcium carbonate (SD 5.26). Daily water quality was measured in reference plots and adjacent to the treatment plots (table 2) with DO >9.0.0 mg/L and pH >8.00.
Table 2.
Mean (standard deviation) water quality (dissolved oxygen, pH, temperature, and specific conductance) in treatment (external to the mats) and reference plots at three sites in Loon Lake, Benzie County, Michigan.[mg/L, milligram per liter; °C, degree Celsius; µS/cm, microsiemens per centimeter at 25 °C; n, number of measurements; CO2, carbon dioxide]
During treatment, mean pH and DO values were similar at all sampling points and did not differ from the benthic mat and reference plots (table 3). During additional bump carbon dioxide treatments, we monitored pH at the edge of the carbon dioxide mat and detected some leakage of carbon dioxide on three occasions. All occurrences of pH less than [<] 7.0 were at site 3 near the edge of the mat (6.47 on August 23, 2024; 6.98 on August 25, 2024; and 6.84 to 6.94 on August 26, 2024). We immediately added sandbags to the area of the mat where the leak was detected and monitored pH until we confirmed that the leak had stopped.
Table 3.
Mean (standard deviation) water quality (dissolved oxygen and pH) in carbon dioxide plots at three locations external to the carbon dioxide mat. Measurements were made daily during carbon dioxide infusion.[mg/L, milligram per liter; n, number of measurements; m, meter]
Dissolved Oxygen Monitoring
The DO data for site 1 are missing for the first 24 hours because of a logging error. Continuous monitoring of DO in the reference plots indicated concentrations remained at >8.0 mg/L (fig. 7). Under the benthic mat and carbon dioxide mat, DO was similar at sites 1 and 2. At site 1, DO fluctuated between ~5.0 and 8.0 mg/L, except for the final day of treatment when the concentration declined <4.0 mg/L under the benthic mat and carbon dioxide mat. At site 2, DO ranged between ~4.0 and 7.0 mg/L under the benthic mat and carbon dioxide mat, and the lowest reading was on day 2 of the treatment. DO was most variable and lowest at site 3. Under the benthic mat at site 3, DO varied on a 24-hour cycle, ranging from about <2.0 to 8.0 mg/L. Under the carbon dioxide mat at site 3, DO trended downward in the initial 48 hours and then stayed mostly at <4.0 mg/L for the remaining 72 hours.
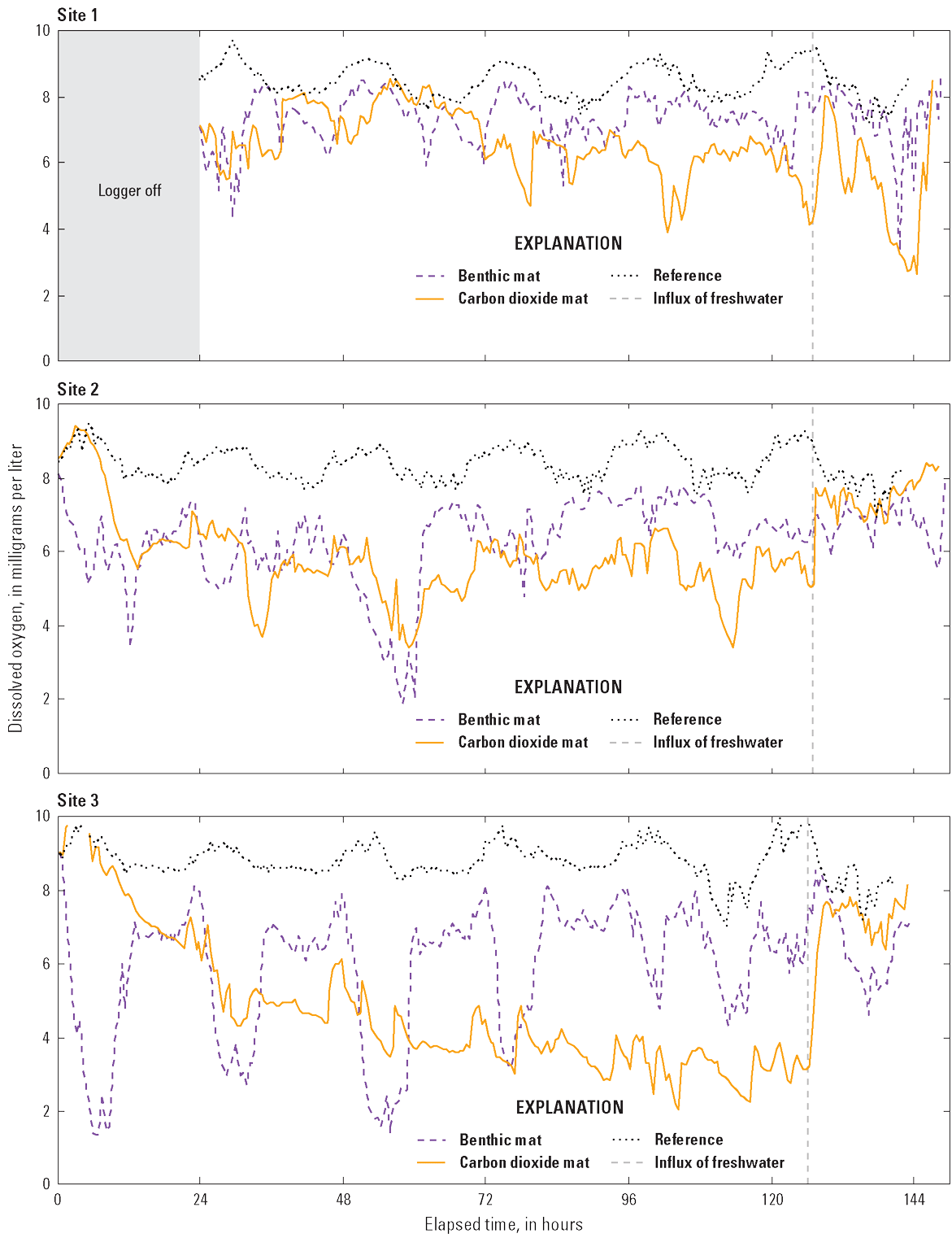
Graphs showing dissolved oxygen concentration in treatment and reference plots at three sites in Loon Lake, Benzie County, Michigan. Carbon dioxide was infused under mats intermittently for 5 days. At the conclusion of the treatment period, the edges of the mat were lifted to allow an influx of freshwater.
Water quality under the carbon dioxide mat was continuously logged by the YSI sonde (fig. 8; table 4). At the completion of treatment on day 5 at site 1, we determined that the sonde had detached from the cinder block and was near an edge of the mat that had become slack, which resulted in wide fluctuations in measurements (pH range from 6.0 to >8.0) and a higher mean pH (6.80) at site 1 (table 4; fig. 8). The pH measurements were more consistent and remained closer to the target at sites 2 and 3 (fig. 8; table 4). Overall, the mean (and estimated) pH was greater than the target pH of 6.13, and the estimated mean carbon dioxide concentration was <200 mg/L at all three sites (table 4). The total mass of carbon dioxide used to treat the three 16-m2 plots for 5 days was 11.5 kilograms (25.3 pounds).
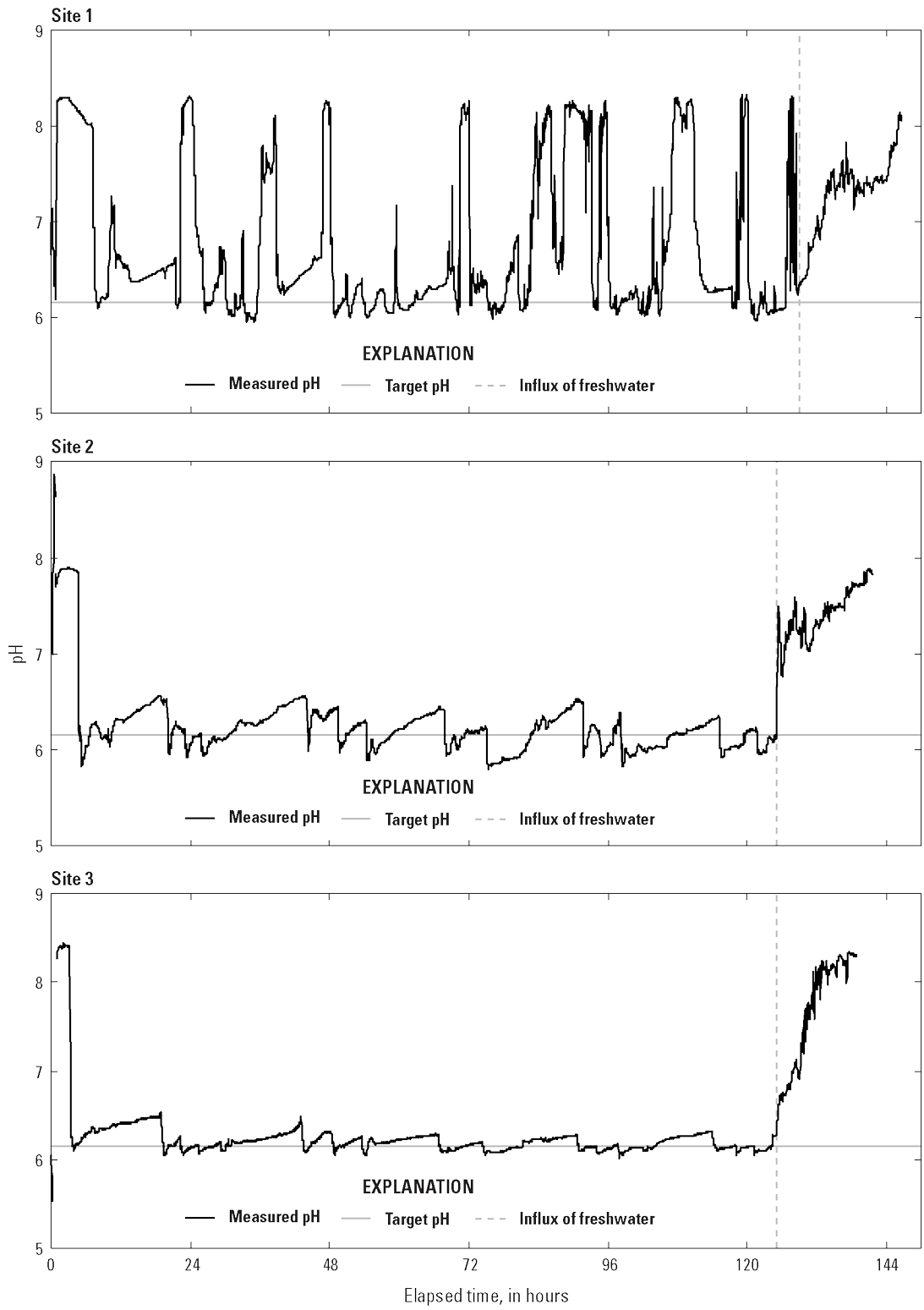
Graphs showing pH concentration under the carbon dioxide mats, as measured by YSI 6920 V2 sondes at each of three sites in Loon Lake, Benzie County, Michigan. Carbon dioxide was infused under mats intermittently for 5 days. At the conclusion of the treatment period, the edges of the mat were lifted to allow an influx of freshwater.
Table 4.
Mean (standard deviation) water quality (dissolved oxygen, pH, temperature, specific conductance, and carbon dioxide) at the three carbon dioxide-infused mat treatment plots. Water quality variables were measured by a YSI sonde unit with pH, dissolved oxygen, and conductivity and temperature probes.[mg/L, milligram per liter; °C, degree Celsius; µS/cm, microsiemens per centimeter at 25 °C; n, number of measurements recorded in 5-day treatment]
Treatment Effects on Resident Zebra Mussels
The effects of the benthic mat and carbon dioxide mat on resident mussels in the plots were confounded by low mussel density (table 5), limited hard substrate, and the effects of carbon dioxide on mussel attachment. Substrates that were colonized with zebra mussels and placed into the carbon dioxide plots before treatment had few to no mussels attached at the posttreatment survey, likely because of carbon dioxide effects on byssal thread detachment (Waller and others, 2020). The entirety of each 4-m x 4-m plot was surveyed, and all colonized substrate was collected in the posttreatment assessment. The estimated survival of mussels in the reference plot did not differ from zero for treatment time (fig. 9A; 0.111, z=0.393, 95-percent CI of −0.442–0.664, p=0.694). Therefore, pre- and posttreatment survey data for the reference plots were combined. The percentage of live zebra mussels in the benthic mat (difference of −2.13, 95-percent CI of −2.97–1.29, z=4.98, p<0.001) and carbon dioxide mat treatments (difference of −1.83, 95-percent CI of −2.48–−1.17, z=5.44, p<0.001) was lower compared to that of the reference site (1.70, 95-percent CI of 1.17–2.23; fig. 9B; table 5).
Table 5.
Survey results of resident Dreissena polymorpha (zebra mussels) in reference and treatment plots.[pre, pretreatment; post, posttreatment; CO2, carbon dioxide]

Graphs showing regression coefficients for density of live Dreissena polymorpha (zebra mussels) in quadrat surveys from logistic regression transformed to probability scale with 95-percent confidence intervals. A, regression coefficients for reference and carbon dioxide and benthic mat treatments; B, regression coefficients for pre- and posttreatment mussel density in reference plots.
Caged Sentinel Zebra Mussel Mortality
Sentinel zebra mussels in the benthic mat (difference of −0.728, z=−6.21, p<0.01, 95-percent CI of −0.956–−0.50) and the carbon dioxide mat (difference of −4.49, z=−30.5, p<0.01, 95-percent CI of −4.79 to −4.21) treatments had lower survival rates than those in the reference plots (1.96, 95-percent CI of 1.78–2.14) (table 6; fig. 10). In carbon dioxide mat treatments, zebra mussels experienced 100-percent mortality at sites 1 and 3 and >80-percent mortality at site 2. In comparison, mortality in benthic mat treatments ranged from 13.5 to 27 percent (table 6; fig. 10).
Table 6.
Mean (standard error) mortality of sentinel caged Dreissena polymorpha (zebra mussels) in reference plots and benthic mat and carbon dioxide mat treatment plots.[n, number of measurements (3 plots per treatment); CO2, carbon dioxide]
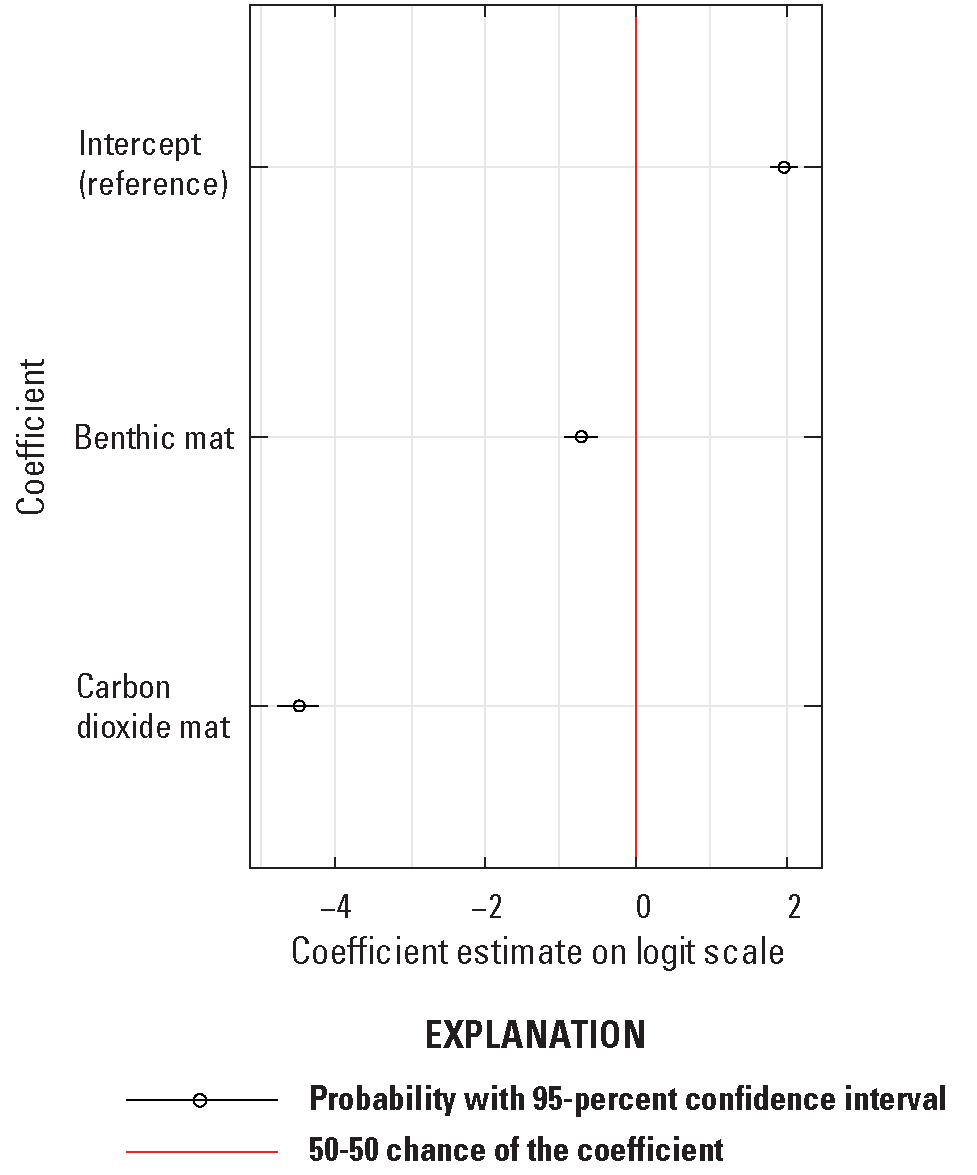
Regression coefficients for mortality of caged Dreissena polymorpha (zebra mussels) from logistic regression transformed to probability scale with 95-percent confidence intervals.
Survival of caged mussels varied by location within the plot, but it differed by treatment and site. Zebra mussel survival in the carbon dioxide treatment was higher in cages located midway and at mat corners (only at site 2; difference of 2.48, 95-percent CI of 1.85–3.11, z=3.89, p<0.001), whereas survival under the benthic mats was lower at the corners (difference of −13.81, 95-percent CI of −13.53–−14.09, z=−4.895, p<0.001; fig. 11).
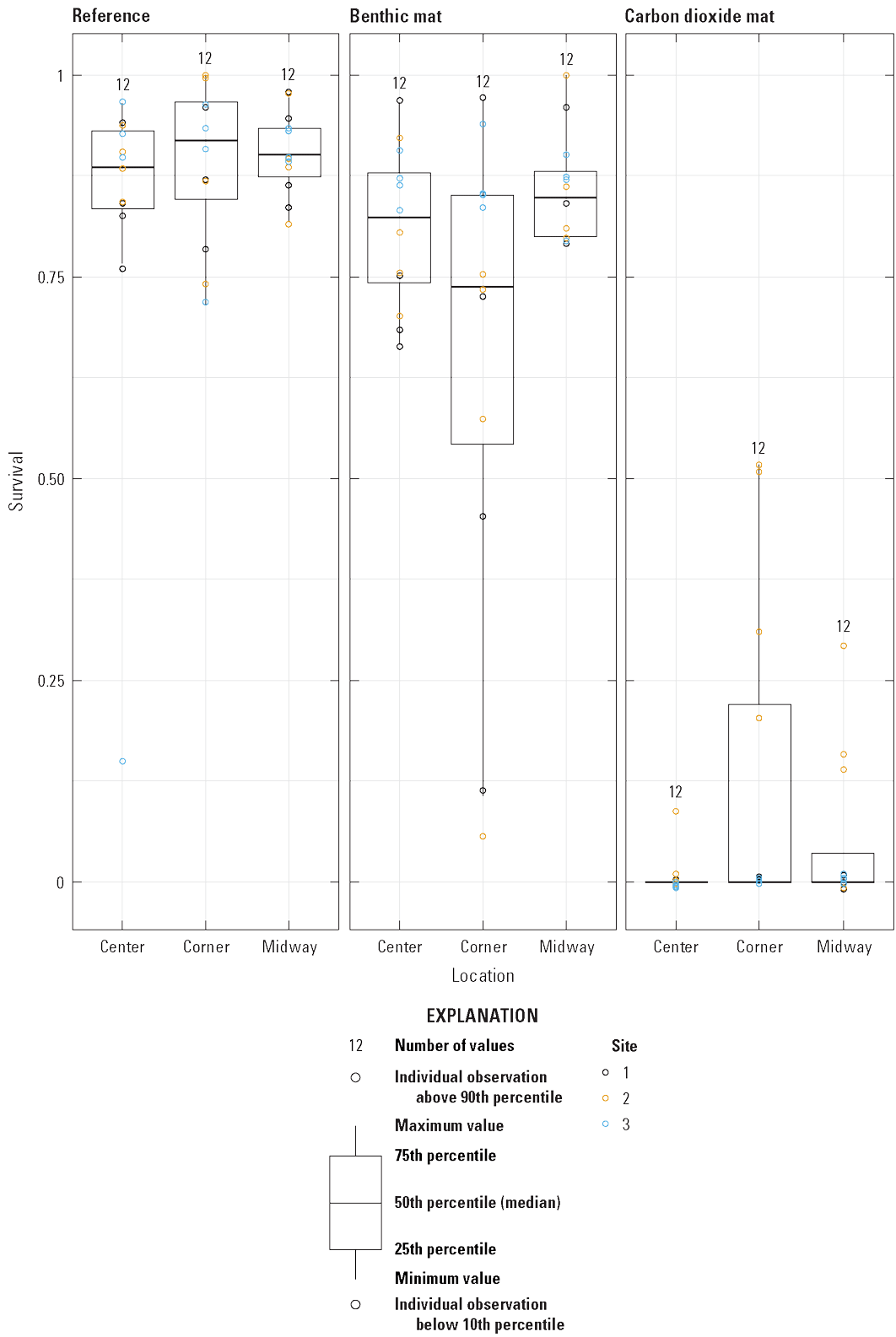
Boxplot showing raw data points for Dreissena polymorpha (zebra mussel) survival by location within the plot and under the benthic or carbon dioxide mat.
Zebra Mussel Lengths
The mean shell length of resident mussels collected in surveys was less than that of caged sentinel mussels (difference of −4.0 95-percent CI of 3.2–5.0, z=8.95, p<0.001; table 7; fig. 12). However, there was no difference in the shell length of mussels within groups at site 1 (mean of 15.1, 95-percent CI of 14.3–15.8) compared to site 2 (difference of −0.2, 95-percent CI of −1.3–0.9, z=−0.380, p=0.704) or site 3 (difference of 0.9, 95-percent CI of −1.3–0.9, z=1.79, p=0.743).
Table 7.
Mean (standard deviation) shell length of representative resident Dreissena polymorpha (zebra mussels) collected in plot surveys (pre- and posttreatment) and sentinel mussels caged in cages and placed in the plots.[mm, millimeter]
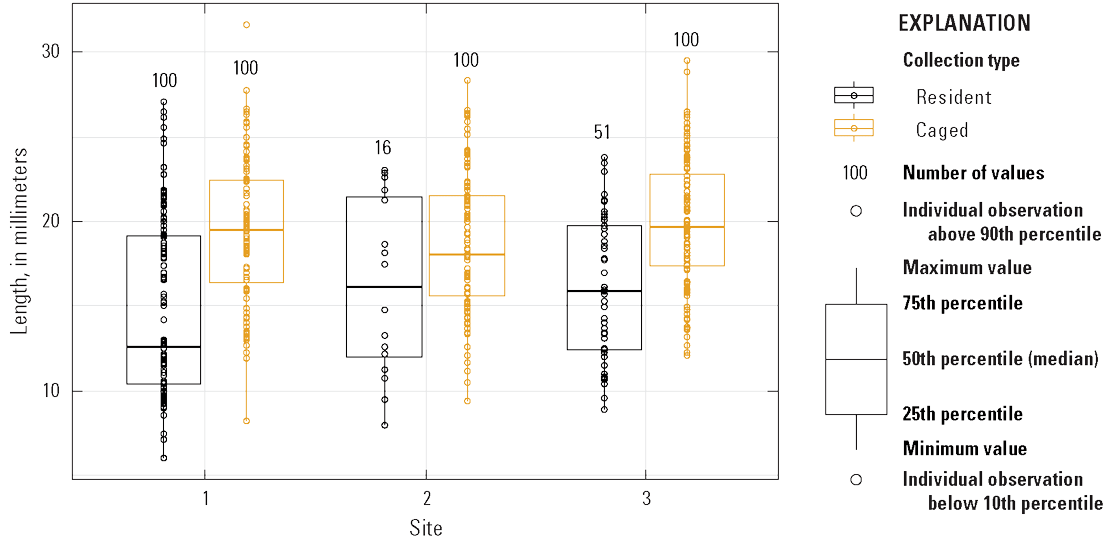
Boxplot showing Dreissena polymorpha (zebra mussel) shell length from each site.
Macroinvertebrate Abundance and Diversity
We compared benthic macroinvertebrate abundance and diversity in the reference plots before and after treatment as a measure of temporal variability in the community during the study period. Within a reference plot, no difference was detected between the pre- and posttreatment sampling periods in macroinvertebrate abundance or diversity (degrees of freedom [df]=1, f-statistic [f]=0.2879, probability [p]=0.777; fig. 13; table 8). Macroinvertebrate abundance and diversity 1 day after treatment were compared between the benthic mat and carbon dioxide mat plots and reference plots. The abundance of benthic macroinvertebrates was less in the benthic and carbon dioxide mat treatments compared to the reference plots (df=2, f=5.76, p=0.002; fig. 13). Diversity did not vary between the reference plots and the mat treatments (df=3, f=2.46, p=0.081; table 8). Distance-based redundancy analysis explained 97 percent of predictive variability with one axis, mostly treatment (variance=3389.7, f=5.76, df=2, p<0.001) and a large amount of residual variance (9416.7, df=32; fig. 14).
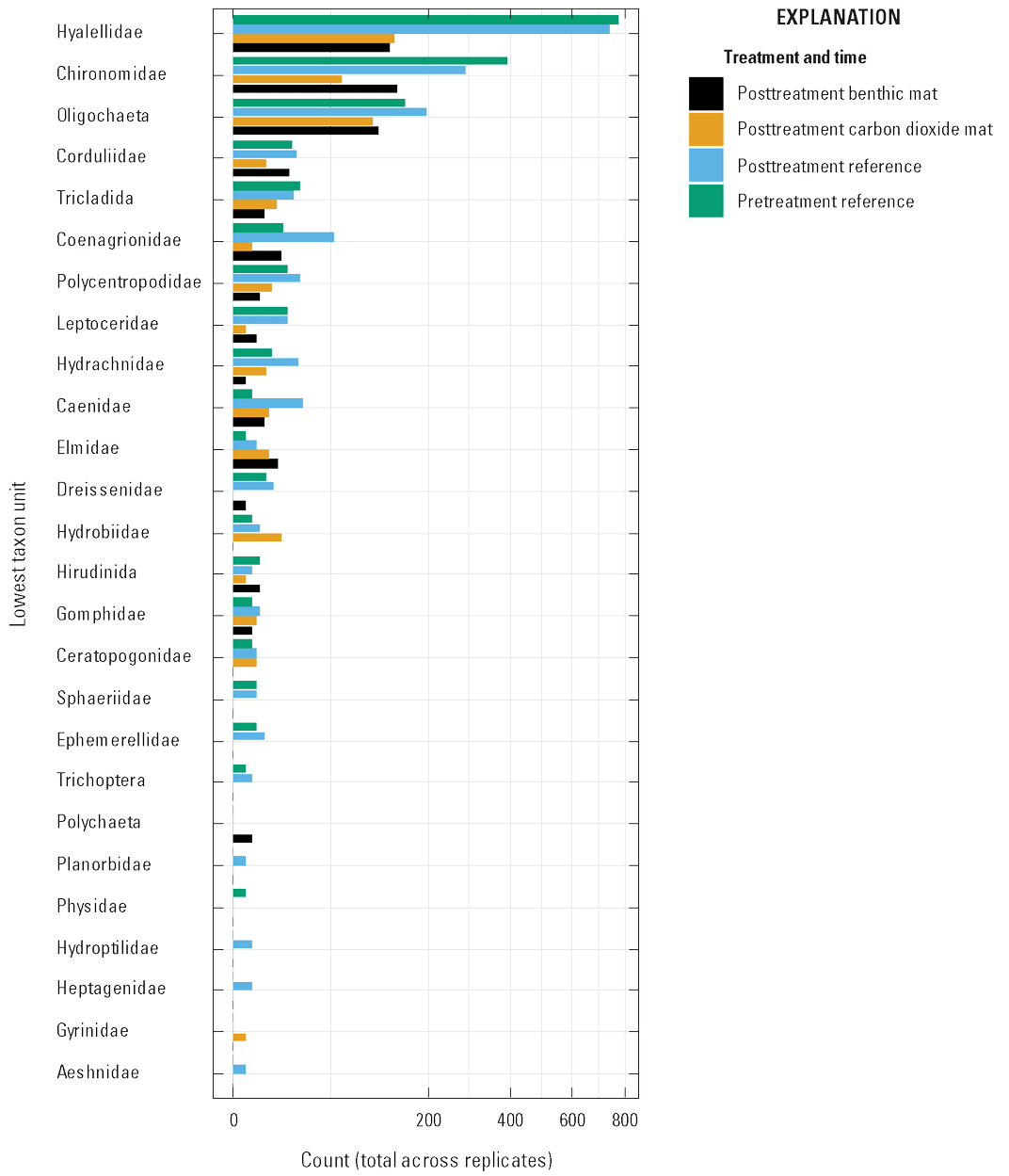
Graph showing total number of macroinvertebrate individuals from each site across all replicates in reference plots (pre- and posttreatment) and in all plots posttreatment.
Table 8.
Mean Shannon-Wiener diversity and abundance (standard error) of macroinvertebrates in reference, benthic mat, and carbon dioxide mat treatments. Benthic mat and carbon dioxide mat treatment plots were not sampled before mat placement to minimize disturbance of the plots.[CO2, carbon dioxide]
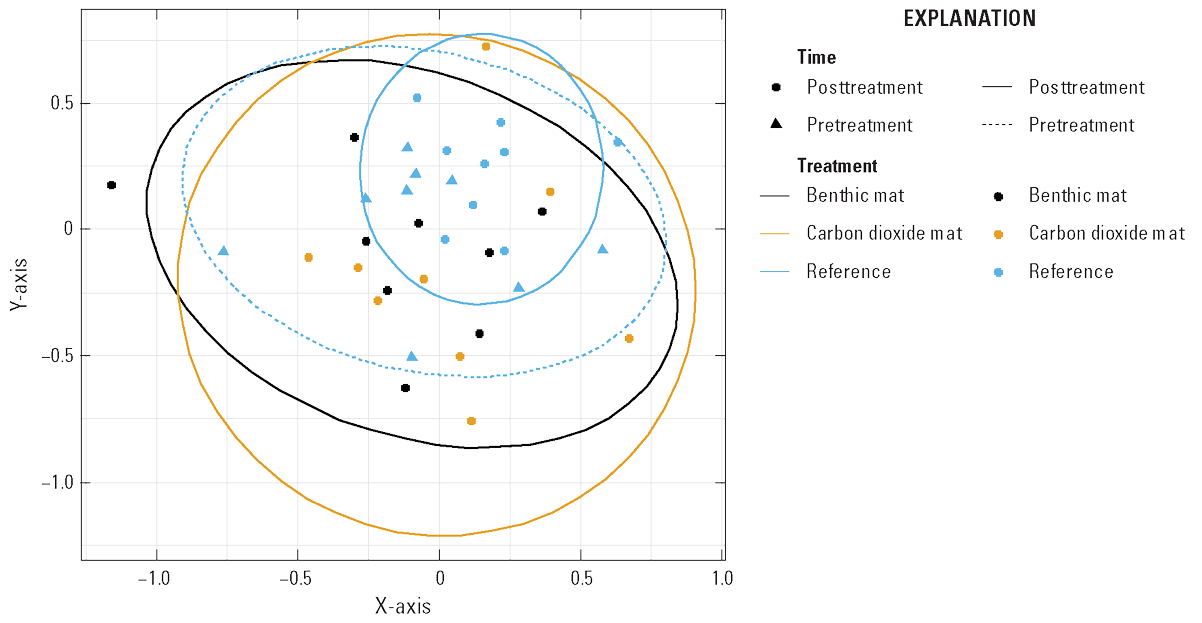
Graph showing distance-based redundancy model of macroinvertebrate community by time (pre- and posttreatment) in the reference plots and posttreatment in all plots.
Discussion
Benthic mat treatment combined with carbon dioxide (mean estimated concentration between 103.0 and 170.5 mg/L for 120 hours) produced >80-percent mortality of caged sentinel zebra mussels compared to a mean mortality of 20.6 percent and 12.7 percent in benthic mat alone and reference plots, respectively. Mussel mortality was almost 100 percent under two of the three carbon dioxide treatment mats, and only at site 2 was 20-percent survival of sentinel mussels observed. Reasons for the survival of mussels at site 2 are unclear and are not explained by differences in pH or carbon dioxide concentration. Mussel mortality in carbon dioxide mat treatments was 100 percent at site 1, where the mean pH was higher (and carbon dioxide lower) and highly variable compared to sites 2 and 3. On the other hand, pH trends were more similar at sites 2 and 3 during the 5-day treatment period, although DO averaged 5.0 mg/L at site 3 compared to an average of >8.0 mg/L at site 2. The location of mussels under the carbon dioxide mat did not affect mortality at sites 1 and 3; survival at site 2 was greater in cages at the corners, indicating possible microhabitat differences, such as spring input and vegetation or reduced carbon dioxide exposure at the corners, may have existed.
Measuring the effect of carbon dioxide on resident mussels was confounded by the sparse number of mussels in the plots, coupled with the detachment response of mussels in carbon dioxide. In posttreatment surveys of the benthic mat and carbon dioxide plots, few mussels were collected in the treatment plots relative to the reference plots. The total density of mussels was lowest in carbon dioxide treatments, followed by the benthic mats (table 5). Differences in density may be due to natural variation among the plots. Another contributing factor may be the detachment response of zebra mussels to a stressor. Exposure to 111 mg/L carbon dioxide for 96 hours at 20 °C was estimated to cause 75 percent of zebra mussels to detach (Waller and others, 2020); therefore, exposure for 5 days at ~170 mg/L carbon dioxide in our study likely caused similar detachment, resulting in “clean” hard substrates in survey collections. Hypoxia and physical smothering with the benthic mat could also induce detachment. Effects of the mats on attachment could be further investigated by placing standard settlement plates and a known number of mussels in the plots or deploying the mats in a system with more hard substrate and higher dreissenid mussel density.
Benthic mats produce mortality by creating anoxia and suffocating mussels; however, zebra mussels are relatively tolerant of hypoxia, depending on water temperature and acclimation temperature (Walz, 1973; Clarke and McMahon, 1996; Yu and Culver, 1999). For example, complete anoxia for 7 days at 22 °C produced 100-percent mortality of zebra mussels (Walz, 1973), whereas Clarke and McMahon (1996) reported that the critical hypoxia threshold for zebra mussels was 0.9–1.3 mg DO/L at 25 °C. The entry of any freshwater under the mat from wave action or springs can further extend the lethal treatment time. In this study, the 5-day treatment with a 4-m x 4-m mat was minimally lethal to zebra mussels. DO decreased to <2.0 mg/L for periods of time, but complete anoxia was not achieved. The limited size and square shape of the benthic mat produced edge effects, as indicated by greater mortality at the center of the mat, and less mortality at the corners, where ingress of freshwater with higher DO was more likely. These results were not unexpected because past studies achieved high mollusc mortality with larger benthic mats deployed for 28 days (Wittmann and others, 2012) to 5 months (Conry and others, 2024). The pattern of DO concentration was similar between the benthic mats and carbon dioxide mats at sites 1 and 2 in the first 72 hours, whereas DO under the benthic mat at site 3 was much more variable. After 72 hours, DO trended downward under every carbon dioxide mat, relative to the comparable benthic mat, perhaps because of an overall increase in respiration.
The effects of benthic mat and carbon dioxide treatments on the macroinvertebrate community were variable and generally minimal. Total macroinvertebrate abundance decreased to a similar level in the treatment plots, compared to the reference plots, but species diversity did not differ, indicating a reduction across taxa. Seven taxa were collected only from the reference plots, all in low abundance (fig. 13). The most abundant taxa in our samples, including Hyalellidae, Chironomidae, and Oligochaeta, are relatively tolerant of low DO (Hilsenhoff, 1988; Barbour and others, 1999). These taxa were less abundant in the benthic mat and carbon dioxide mat plots, but none were eliminated by the treatments. Moderately sensitive taxa, including Corduliidae, Polycentropodiae, Leptoceridae, Elmidae, and Ceratopogonidae (Hilsenhoff, 1988; Barbour and others, 1999), were detected in the carbon dioxide treatment plots in varying abundance compared to the reference plots (fig. 13). The most sensitive taxa in our samples, Gomphidae, Aeshnidae, and Ephemerellidae (Hilsenhoff, 1988; Barbour and others, 1999), were low in abundance. Of these, Gomphidae was collected from benthic mat and carbon dioxide mat treatments and in similar numbers to the reference plots. Because of the low abundance of organisms in these taxa, we could not test for treatment effects. Our study could have benefited from additional sampling of woody debris and vegetation to potentially increase diversity and the number of sensitive taxa. Repeating sampling after treatment, such as 7 days and 30 days posttreatment, could also provide information on how quickly macroinvertebrates recolonized the area.
The anoxia produced under benthic mats is selective only for those organisms that tolerate low DO, such as chironomids and oligochates; however, benthic mats are most often deployed for weeks to months, creating anoxia that would kill tolerant taxa (Wittmann and others, 2012; Conry and others, 2024). Carbon dioxide is selective for zebra mussels compared to native unionid mussels in short, acute exposures (Waller and others, 2020). Based on our results, treatment with carbon dioxide under a mat for 5 days would pose no more risk to macroinvertebrates than a benthic mat for the same period but would be much more effective for killing dreissenid mussels.
This was a proof-of-concept trial, and the system could be improved to increase spatial coverage and feasibility of the carbon dioxide benthic mat treatment. Compared to the carbon dioxide mat, a benthic mat is simple and requires only placement and retrieval; however, the benthic mat must remain in place for an extended period and will cause substantial nontarget mortality. The carbon dioxide mat requires specialized construction for infusion and delivery of carbon dioxide. Enlarging the size of a single treatment area would require a revised design of the mat used in this study. The rigid PVC frame in our mat was unwieldy to deploy and retrieve, and scaling up to longer arms would only decrease its practicality. The next iteration of the carbon dioxide mat could evaluate a hydrostatic frame for support of the mat and delivery of the carbon dioxide-infused water. The treatment area could most simply be enlarged by placing multiple mats in an area and setting up a central infusion point. The system for manually infusing carbon dioxide produced variability in concentrations, especially between dusk and dawn when treatments were not applied. Other methods for delivery of the carbon dioxide under the mat could be explored, including an automated monitoring system that turns on when pH increases to more than an established threshold. Other alternatives to water-infused carbon dioxide could be seeding the treatment plot with dry ice pellets immediately before mat placement. Additionally, an exposure period <5 days can be lethal to dreissenids (Barbour and others, 2024). Continuous exposure to ~170–200 mg/L carbon dioxide for 96–100 hours produced 99-percent mortality of zebra mussels at 20 °C. Although this exposure was continuous, mussels were not stressed by low DO concentration, which remained at >6.0 mg/L (Waller and others, 2020). Even shorter treatment (48–72 hours) could be effective if done during midsummer when water temperatures are higher (>20 °C), hypoxia under the mat is likely, and mussels are more sensitive after spawning (Costa and others, 2008).
Summary
This study tested a novel system for delivering carbon dioxide-infused water under a mat as a tool to kill invasive mussels. We compared the efficacy of a benthic mat alone with carbon dioxide infusion under a mat for killing zebra mussels (Dreissena polymorpha) (Pallas, 1771). The study was completed in August 2024 in Sleeping Bear Dunes National Lakeshore in Loon Lake, Benzie County, Michigan. Three sites were selected in the lake for replication of reference, benthic mat, and carbon dioxide mat treatments. Within a site, three 4-meter (m) x 4-m plots were delineated for each treatment and a reference. We completed pretreatment sampling to estimate zebra mussel density and macroinvertebrate community composition in reference plots. Zebra mussels (about 360) from outside of the treatment plots were caged and placed in the plots before treatment. Benthic mats (4.25 m x 4.25 m; polyethylene with a vinyl coating) were placed on the bottom and anchored with sandbags and weights. Carbon dioxide was infused under a mat of the same material and size. Water was pumped from the lake into two downward-flow gas infusion chambers, and the outflowing water was connected to a single hose that delivered carbon dioxide-infused water to the midpoint of the mat at a target concentration of 200 milligrams per liter (mg/L), equivalent to a pH of about 6.13. Carbon dioxide-infused water was injected under the mats every 2–4 hours from dawn to dusk, about 12 hours, as needed to maintain the concentration near 200 mg/L.
The benthic mat and carbon dioxide mat were deployed for 5 days. Water quality (temperature, dissolved oxygen, and pH) was monitored continuously under the carbon dioxide mat, and “bump” carbon dioxide treatments were applied every 2–4 hours from dawn to dusk to maintain the target concentration, as estimated from pH measurements. Dissolved oxygen was continuously measured in all plots with a MiniDOot sensor. We surveyed plots 1 day after the termination of treatment and removal of mats to estimate the percentage of dead zebra mussels. We then assessed the mortality of caged mussels in each plot. Macroinvertebrates were sampled with a petite Ponar and identified and enumerated to determine total abundance and diversity. During the 5-day treatment period, water quality in the areas external to the mats was similar among treatment and reference plots. Under the benthic and carbon dioxide mats, dissolved oxygen varied among sites, ranging from 2.0 to 8.0 mg/L, but was generally lower compared to the reference plots. The pH under the carbon dioxide mats averaged 6.80 (site 1) to 6.38 (site 3), equal to a carbon dioxide concentration of 103.9–170.5 mg/L, respectively, over the 5-day period. The posttreatment survey detected relatively few resident zebra mussels in the benthic mat and carbon dioxide mat treatment plots relative to the reference plots; despite low abundance, the mean percentage alive was lowest in the carbon dioxide treatment followed by 50 percent alive in the benthic mat plots. Mortality of caged sentinel mussels was greater than 80 percent in carbon dioxide treatments compared to a mean mortality of 20.6 percent under benthic mats and 12.7 percent in the untreated reference plots. The benthic mat and carbon dioxide mat treatments had similar effects on the macroinvertebrate community. Abundance was reduced in both treatments, but diversity was similar to the reference plots. The study demonstrated that a benthic mat with carbon dioxide infusion can produce substantially greater mortality of zebra mussels than a benthic mat alone. Additional work to improve the carbon dioxide mat could focus on improving design for deployment and retrieval, increasing mat size, determining the minimum effective carbon dioxide treatment time, evaluating alternative carbon dioxide delivery methods, and identifying additional measures that could increase efficacy, such as the addition of biodegradable organics.
References Cited
Barbour, M.T., Gerritsen, J., Snyder, B.D., and Stribling, J.B., 1999, Rapid bioassessment protocols for use in streams and wadeable rivers—Periphyton, benthic macroinvertebrates and fish (2d ed.): Washington, D.C., U.S. Environmental Protection Agency, Office of Water, EPA 841–B–99–002, 337 p., accessed February 24, 2026, at https://www3.epa.gov/region1/npdes/merrimackstation/pdfs/ar/AR-1164.pdf.
Barbour, M.T., Meulemans, M.J., Severson, T.J., Wise, J.K., and Waller, D.L., 2024, Carbon dioxide toxicity to zebra mussels (Dreissena polymorpha) is dependent on water chemistry: Environmental Toxicology and Chemistry, v. 43, no. 6, p. 1312–1319. [Also available at https://doi.org/10.1002/etc.5864.]
Bates, D., Mächler, M., Bolker, B., and Walker, S., 2015, Fitting linear mixed-effects models using lme4: Journal of Statistical Software, v. 67, no. 1, p. 1–48. [Also available at https://doi.org/10.18637/jss.v067.i01.]
Beason, E., and Schwalb, A.N., 2022, Impact of zebra mussels on physiological conditions of unionid mussels in Texas: Aquatic Sciences, v. 84, no. 2, p. 21, accessed March 3, 2022, at https://doi.org/10.1007/s00027-022-00853-8.
Clarke, M., and McMahon, R.F., 1996, Effects of hypoxia and low-frequency agitation on byssogenesis in the freshwater mussel Dreissena polymorpha (Pallas): The Biological Bulletin, v. 191, no. 3, p. 413–420, accessed April 28, 2025, at https://doi.org/10.2307/1543014.
Conry, T.M., Tibbs, J.E., Baird, M.S., Van Zee, B.E., McLane, H.R., Mobley, B.W., Southard, G.M., and McGarrity, M.E., 2024, Rapid response and eradication of zebra mussels (Dreissena polymorpha) from Lake Waco, Texas, USA, using a gas impermeable benthic barrier approach: Management of Biological Invasions, v. 15, no. 2, p. 239–250. [Also available at https://doi.org/10.3391/mbi.2024.15.2.05.]
Costa, R., Aldridge, D.C., and Moggridge, G.D., 2008, Seasonal variation of zebra mussel susceptibility to molluscicidal agents: Journal of Applied Ecology, v. 45, no. 6, p. 1712–1721. [Also available at https://doi.org/10.1111/j.1365-2664.2008.01555.x.]
Cunningham, K.E., and Dunlop, E.S., 2023, Declines in lake whitefish larval densities after dreissenid mussel establishment in Lake Huron: Journal of Great Lakes Research, v. 49, no. 2, p. 491–505. [Also available at https://doi.org/10.1016/j.jglr.2022.12.015.]
Dzierżyńska‐Białończyk, A., Jermacz, Ł., Maćkiewicz, T., Gajewska, J., and Kobak, J., 2018, Mechanisms and impact of differential fouling of the zebra mussel Dreissena polymorpha on different unionid bivalves: Freshwater Biology, v. 63, no. 7, p. 687–699. [Also available at https://doi.org/10.1111/fwb.13107.]
Haag, W.R., Berg, D.J., Garton, D.W., and Farris, J.L., 1993, Reduced survival and fitness in native bivalves in response to fouling by the introduced zebra mussel (Dreissena polymorpha) in western Lake Erie: Canadian Journal of Fisheries and Aquatic Sciences, v. 50, no. 1, p. 13–19. [Also available at https://doi.org/10.1139/f93-002.]
Hansen, G.J.A., Ahrenstorff, T.D., Bethke, B.J., Dumke, J.D., Hirsch, J., Kovalenko, K.E., LeDuc, J.F., Maki, R.P., Rantala, H.M., and Wagner, T., 2020, Walleye growth declines following zebra mussel and Bythotrephes invasion: Biological Invasions, v. 22, no. 4, p. 1481–1495. [Also available at https://doi.org/10.1007/s10530-020-02198-5.]
Hebert, P.D.N., Muncaster, B.W., and Mackie, G.L., 1989, Ecological and genetic studies on Dreissena polymorpha (Pallas)—A new mollusc in the Great Lakes: Canadian Journal of Fisheries and Aquatic Sciences, v. 46, no. 9, p. 1587–1591. [Also available at https://doi.org/10.1139/f89-202.]
Higgins, S.N., and Vander Zanden, M.J., 2010, What a difference a species makes—A meta-analysis of dreissenid mussel impacts on freshwater ecosystems: Ecological Monographs, v. 80, no. 2, p. 179–196, accessed January 11, 2019, at https://doi.org/10.1890/09-1249.1.
Hilsenhoff, W.L., 1988, Rapid field assessment of organic pollution with a family-level biotic index: Journal of the North American Benthological Society, v. 7, no. 1, p. 65–68. [Also available at https://doi.org/10.2307/1467832.]
Kunze, T., 2023, Benthic algal and macroinvertebrate response to the removal of dreissenid mussels in the nearshore zone of Lake Michigan: University of Wisconsin–Milwaukee, master’s thesis, 109 p., accessed April 24, 2025, at https://minds.wisconsin.edu/bitstream/handle/1793/93327/Kunze_uwm_0263M_13600.pdf?sequence=1&isAllowed=y.
Kuznetsova, A., Brockhoff, P.B., and Christensen, R.H.B., 2017, lmerTest package—Tests in linear mixed effects models: Journal of Statistical Software, v. 82, no. 13, accessed January 8, 2025, at https://doi.org/10.18637/jss.v082.i13.
LimnoTech, 2020, Good Harbor Bay dreissenid mussel control demonstration project—Final project report: Ann Arbor, Mich., Great Lakes Commission and Invasive Mussel Collaborative Partners, prepared by LimnoTech, 84 p., accessed April 2, 2025, at https://invasivemusselcollaborative.net/wp-content/uploads/2020/12/Final_Report_Good_Harbor_Zequanox.pdf.
MacIsaac, H.J., 1996, Potential abiotic and biotic impacts of zebra mussels on the inland waters of North America: American Zoologist, v. 36, no. 3, p. 287–299. [Also available at https://doi.org/10.1093/icb/36.3.287.]
McEachran, M.C., Trapp, R.S., Zimmer, K.D., Herwig, B.R., Hegedus, C.E., Herzog, C.E., and Staples, D.F., 2019, Stable isotopes indicate that zebra mussels (Dreissena polymorpha) increase dependence of lake food webs on littoral energy sources: Freshwater Biology, v. 64, no. 1, p. 183–196, accessed January 8, 2025, at https://doi.org/10.1111/fwb.13206.
Meulemans, M.J., Waller, D.L., Wise, J.K., Severson, T.J., Barbour, M.T., and Morris, B.E., 2026, Evaluation of benthic barrier layers/tarps for open water control of invasive mussels in 2024 in Loon Lake, Benzie Co., Michigan, USA: U.S. Geological Survey data release, accessed February 13, 2026, at https://doi.org/10.5066/P13NC3TU.
Mills, E.L., Rosenberg, G., Spidle, A.P., Ludyanskiy, M., Pligin, Y., and May, B., 1996, A review of the biology and ecology of the quagga mussel (Dreissena bugensis), a second species of freshwater dreissenid introduced to North America: American Zoologist, v. 36, no. 3, p. 271–286. [Also available at https://doi.org/10.1093/icb/36.3.271.]
Oksanen, J., Simpson, G.L., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P.R., O’Hara, R.B., Solymos, P., Henry, M., Stevens, H., Szoecs, E., Wagner, H., Barbour, M., Bedward, M., Bolker, B., Borcard, D., Borman, T., Carvalho, G., Chirico, M., De Caceres, M., Durand, S., Antoniazi Evangelista, H.B., FitzJohn, R., Friendly, M., Furneaux, B., Hannigan, G., Hill, M.O., Lahti, L., Martino, C., McGlinn, D., Ouellette, M.-H., Cunha, E.R., Smith, T., Stier, A., Ter Braak, C.J.F., and Weedon, J., 2025, vegan—Community ecology package, version 2.7–2: R Foundation for Statistical Computing software release, accessed June 1, 2025, at https://doi.org/10.32614/CRAN.package.vegan.
R Core Team, 2025, R—A language and environment for statistical computing, version 4.5.1 (Great Square Root): R Foundation for Statistical Computing software release, accessed March 4, 2025, at https://www.R-project.org/.
U.S. Environmental Protection Agency, 2019, US EPA pesticide product label, Carbon Dioxide–Carp: U.S. Environmental Protection Agency, 6 p., accessed February 9, 2021, at https://www3.epa.gov/pesticides/chem_search/ppls/006704-00095-20190419.pdf.
Waller, D., Bartsch, L., Bartsch, M.R., Meulemans, M., Severson, T., and Zolper, T.J., 2021, Use of carbon dioxide to prevent zebra mussel (Dreissena polymorpha) settlement and effects on native mussels (Order Unionoida) and benthic communities: Management of Biological Invasion, v. 12, no. 4, p. 927–951. [Also available at https://doi.org/10.3391/mbi.2021.12.4.10.]
Waller, D.L., Bartsch, M.R., Lord, E.G., and Erickson, R.A., 2020, Temperature‐related responses of an invasive mussel and 2 unionid mussels to elevated carbon dioxide: Environmental Toxicology and Chemistry, v. 39, no. 8, p. 1546–1557. [Also available at https://doi.org/10.1002/etc.4743.]
Waller, D.L., Meulemans, M.J., Wise, J., Erickson, R.A., Barbour, M.T., and Morris, B.E.C., 2026, Analysis of open water control of invasive mussels using benthic mats. Part 1—Short-term infusion of carbon dioxide under a mat: U.S. Geological Survey software release, https://doi.org/10.5066/P13JUBYH.
Wittmann, M.E., Chandra, S., Reuter, J.E., Schladow, S.G., Allen, B.C., and Webb, K.J., 2012, The control of an invasive bivalve, Corbicula fluminea, using gas impermeable benthic barriers in a large natural lake: Environmental Management, v. 49, no. 6, p. 1163–1173, accessed May 2, 2025, at https://doi.org/10.1007/s00267-012-9850-5.
Yu, N., and Culver, D.A., 1999, In situ survival and growth of zebra mussels (Dreissena polymorpha) under chronic hypoxia in a stratified lake: Hydrobiologia, v. 392, no. 2, p. 205–215, accessed April 29, 2025, at https://doi.org/10.1023/A:1003697231917.
Zorn, T.G., and Kramer, D.R., 2022, Changes in habitat conditions, fish populations, and the fishery in northern Green Bay, Lake Michigan, 1989–2019: North American Journal of Fisheries Management, v. 42, no. 3, p. 549–571. [Also available at https://doi.org/10.1002/nafm.10715.]
Conversion Factors
International System of Units to U.S. customary units
Temperature in degrees Celsius (°C) may be converted to degrees Fahrenheit (°F) as follows:
°F = (1.8 × °C) + 32.
Datums
Vertical coordinate information is referenced to the North American Vertical Datum of 1988 (NAVD 88).
Horizontal coordinate information is referenced to the North American Datum of 1983 (NAD 83).
Supplemental Information
Specific conductance is in microsiemens per centimeter at 25 degrees Celsius (µS/cm at 25 °C).
Concentrations of chemical constituents in water are in either milligrams per liter (mg/L) or micrograms per liter (µg/L).
Screen size is given in microns.
Mat thickness is given in mil.
For more information about this publication, contact:
Director, USGS Upper Midwest Environmental Sciences Center
2630 Fanta Reed Road
La Crosse, Wisconsin 54603
608–783–6451
For additional information, visit: https://www.usgs.gov/centers/umesc
Publishing support provided by the
USGS Science Publishing Network,
Rolla Publishing Service Center
Disclaimers
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested Citation
Waller, D.L., Erickson, R.A., Wise, J.K., Meulemans, M.J., Morris, B.E.C., Severson, T.J., and Barbour, M.T., 2026, Open water control of invasive mussels using benthic mats—Part 1, short-term infusion of carbon dioxide under a mat: U.S. Geological Survey Open-File Report 2026–1019, 22 p., https://doi.org/10.3133/ofr20261019.
ISSN: 2331-1258 (online)
Study Area
| Publication type | Report |
|---|---|
| Publication Subtype | USGS Numbered Series |
| Title | Open water control of invasive mussels using benthic mats—Part 1, short-term infusion of carbon dioxide under a mat |
| Series title | Open-File Report |
| Series number | 2026-1019 |
| DOI | 10.3133/ofr20261019 |
| Publication Date | June 08, 2026 |
| Year Published | 2026 |
| Language | English |
| Publisher | U.S. Geological Survey |
| Publisher location | Reston, VA |
| Contributing office(s) | Upper Midwest Environmental Sciences Center |
| Description | Report: viii; 22 p.; Data Release; Software Release |
| Larger Work Title | USGS Open-File Report |
| Country | United States |
| State | Michigan |
| County | Benzie County |
| Other Geospatial | Loon Lake |
| Online Only (Y/N) | Y |
| Additional Online Files (Y/N) | N |