FISC - St. Petersburg
|
Primary Datasets
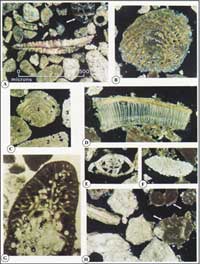
Thin Sections of Surface Sediments: Surface sediments were analyzed to determine the distribution and origin of dominant sand grains following identification criteria of Scholle (1978). Sand grains and sediments in a coral reef environment are derived from skeletal components of calcifying organisms. Each skeletal grain has a specific crystal structure characteristic of a specific organism group, such as calcified red or green algae, corals, molluscs, sea urchins, and so forth, even including microscopic shelled protozoans such as foraminifera (Fig. 12; see Open-File Report 02-192).
To detect this structure and identify the group of origin, grains must be sliced and examined in thin section. Sediments are impregnated with epoxy resin, mounted on a glass slide, and ground down to make thin sections. Thin sections are examined under a special petrographic microscope that transmits polarized light through the glass slide and the grains, illuminating the diagnostic crystal structures. The three most dominant types of sand grains in the Florida-Caribbean region belong to molluscs, species of a calcifying green alga (Halimeda), and corals (see Sedimentary Grains map and Open-File Report 97-453). Sand-grain percentages in surface sediments reveal the kinds of organisms in the local biologic community and are thus indicators of the type of environment in which those organisms are known to live. Where molluscan grains dominate the sand, living molluscs dominate the community. The same is said for Halimeda grains or for grains of other organisms. Coral is the exception. Coral grains dominate areas where dead or dying corals are undergoing bioerosion (e.g., Aronson and Precht, 1997; Lidz and Hallock, 2000). High percentages of coral grains are thus biologic (source) and geologic (sand) indicators of poor coral health. Thorp (1935), Ginsburg (1956), Swinchatt (1965), and Enos (1977) examined grain composition in modern sediments along limited transects on the Florida shelf. The USGS mapped sand-grain percentages throughout the Florida reef tract (Lidz and Hallock, 2000), providing the only shelf-wide baseline dataset for use in monitoring future petrographic changes and thus spatial changes in structure/environment of biologic communities. Comparative studies show that percentages of coral grains in Florida sands increased significantly between the 1950s and 1990s (Lidz et al., 1985; Lidz and Hallock, 2000). An increase has been noted in sands around coral reefs elsewhere as well (Belize, Aronson and Precht, 1997, 2001; Jamaica, Precht and Aronson, 1997). The increase is consistent with coral reef decline and is due to a process called bioerosion (i.e., skeletal breakdown by other organisms). The first bioeroders to attack dead corals are organisms that bore into the skeleton. These 'first responders' include certain fungi, sponges, molluscs, and clams. Certain fish, such as parrotfish, and sea urchins like the black spiny urchin also graze the skeleton for its algal coat, breaking off skeletal bits. Recent temporal changes in biologic community structure at a specific site can be similarly determined by examining thin sections of sands in a sediment core, but such a study has not been done. Thin-section analyses are used in conjunction with other data to evaluate environmental change at the site through time. Other researchers have conducted petrographic studies on the Miami Limestone and Key Largo Limestone (e.g., Hoffmeister and Multer, 1964; Multer and Hoffmeister, 1968; Enos and Perkins, 1977; Coniglio, 1981; Harrison et al., 1982; Coniglio and Harrison, 1983; Halley and Evans, 1983; McNeill et al., 1996; Multer et al., 2002). Those studies were done from a perspective of identifying depositional framework and did not provide grain percentages that could be used to compare with percentages in modern sediments. |

![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: [disc] /pubs/pp/2007/1751/professional-paper/thin-sections.html
Page Contact Information: Feedback
Page Last Modified: December 01, 2016 @ 04:13 PM (JSS)